Viral Infection of Human Natural Killer Cells


Abstract
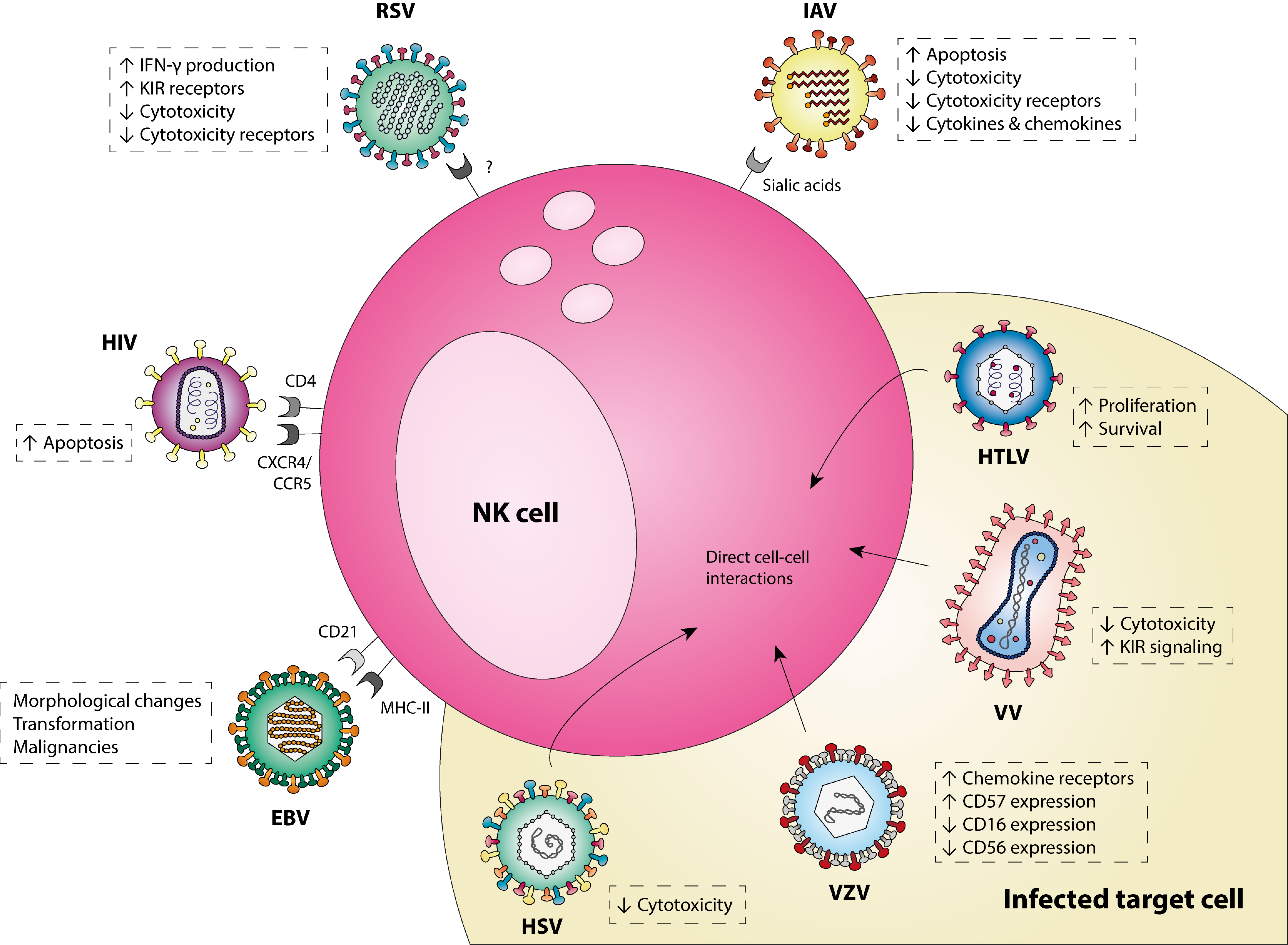
:1. Introduction
2. Entry Mechanisms
2.1. Receptor-Mediated Viral Entry
2.2. Cell–Cell Interaction-Mediated Viral Entry
2.3. Unknown Internalization Mechanism
3. Modulation of NK Cell Function and Phenotype
3.1. Influence on Effector Function
3.2. Influence on Phenotype
4. Contribution to Viral Load
5. Conclusion and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Lodoen, M.B.; Lanier, L.L. Natural killer cells as an initial defense against pathogens. Curr. Opin. Immunol. 2006, 18, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Orange, J.S. Human natural killer cell deficiencies and susceptibility to infection. Microbes Infect. 2002, 4, 1545–1558. [Google Scholar] [CrossRef]
- Smyth, M.J.; Cretney, E.; Kelly, J.M.; Westwood, J.A.; Street, S.E.; Yagita, H.; Takeda, K.; van Dommelen, S.L.; Degli-Esposti, M.A.; Hayakawa, Y. Activation of NK cell cytotoxicity. Mol. Immunol. 2005, 42, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Wiley, S.R.; Schooley, K.; Smolak, P.J.; Din, W.S.; Huang, C.P.; Nicholl, J.K.; Sutherland, G.R.; Smith, T.D.; Rauch, C.; Smith, C.A.; et al. Identification and characterization of a new member of the TNF family that induces apoptosis. Immunity 1995, 3, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.S. Biology of Natural Killer Cells in Cancer and Infection. Cancer Investig. 2002, 20, 405–419. [Google Scholar] [CrossRef]
- Pegram, H.J.; Andrews, D.M.; Smyth, M.J.; Darcy, P.K.; Kershaw, M.H. Activating and inhibitory receptors of natural killer cells. Immunol. Cell Biol. 2011, 89, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Fruh, K.; Gruhler, A.; Krishna, R.M.; Schoenhals, G.J. A comparison of viral immune escape strategies targeting the MHC class I assembly pathway. Immunol. Rev. 1999, 168, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Karre, K.; Ljunggren, H.G.; Piontek, G.; Kiessling, R. Selective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategy. Nature 1986, 319, 675–678. [Google Scholar] [CrossRef]
- Arnon, T.I.; Lev, M.; Katz, G.; Chernobrov, Y.; Porgador, A.; Mandelboim, O. Recognition of viral hemagglutinins by NKp44 but not by NKp30. Eur. J. Immunol. 2001, 31, 2680–2689. [Google Scholar] [CrossRef]
- Mandelboim, O.; Lieberman, N.; Lev, M.; Paul, L.; Arnon, T.I.; Bushkin, Y.; Davis, D.M.; Strominger, J.L.; Yewdell, J.W.; Porgador, A. Recognition of haemagglutinins on virus-infected cells by NKp46 activates lysis by human NK cells. Nature 2001, 409, 1055–1060. [Google Scholar] [CrossRef]
- Mercer, J.; Helenius, A. Gulping rather than sipping: Macropinocytosis as a way of virus entry. Curr. Opin. Microbiol. 2012, 15, 490–499. [Google Scholar] [CrossRef]
- Smyth, L.A.; Afzali, B.; Tsang, J.; Lombardi, G.; Lechler, R.I. Intercellular transfer of MHC and immunological molecules: Molecular mechanisms and biological significance. Am. J. Transplant. 2007, 7, 1442–1449. [Google Scholar] [CrossRef]
- Mao, H.; Tu, W.; Qin, G.; Law, H.K.; Sia, S.F.; Chan, P.L.; Liu, Y.; Lam, K.T.; Zheng, J.; Peiris, M.; et al. Influenza virus directly infects human natural killer cells and induces cell apoptosis. J. Virol. 2009, 83, 9215–9222. [Google Scholar] [CrossRef]
- Guo, H.; Kumar, P.; Moran, T.M.; Garcia-Sastre, A.; Zhou, Y.; Malarkannan, S. The functional impairment of natural killer cells during influenza virus infection. Immunol. Cell Biol. 2009, 87, 579–589. [Google Scholar] [CrossRef] [Green Version]
- Manicassamy, B.; Manicassamy, S.; Belicha-Villanueva, A.; Pisanelli, G.; Pulendran, B.; Garcia-Sastre, A. Analysis of in vivo dynamics of influenza virus infection in mice using a GFP reporter virus. Proc. Natl. Acad. Sci. USA 2010, 107, 11531–11536. [Google Scholar] [CrossRef] [Green Version]
- Shinya, K.; Ebina, M.; Yamada, S.; Ono, M.; Kasai, N.; Kawaoka, Y. Avian flu: Influenza virus receptors in the human airway. Nature 2006, 440, 435–436. [Google Scholar] [CrossRef]
- Mao, H.; Tu, W.; Liu, Y.; Qin, G.; Zheng, J.; Chan, P.L.; Lam, K.T.; Peiris, J.S.; Lau, Y.L. Inhibition of human natural killer cell activity by influenza virions and hemagglutinin. J. Virol. 2010, 84, 4148–4157. [Google Scholar] [CrossRef]
- Chehimi, J.; Bandyopadhyay, S.; Prakash, K.; Perussia, B.; Hassan, N.F.; Kawashima, H.; Campbell, D.; Kornbluth, J.; Starr, S.E. In vitro infection of natural killer cells with different human immunodeficiency virus type 1 isolates. J. Virol. 1991, 65, 1812–1822. [Google Scholar]
- Valentin, A.; Rosati, M.; Patenaude, D.J.; Hatzakis, A.; Kostrikis, L.G.; Lazanas, M.; Wyvill, K.M.; Yarchoan, R.; Pavlakis, G.N. Persistent HIV-1 infection of natural killer cells in patients receiving highly active antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2002, 99, 7015–7020. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, H.B.; Wang, G.; Plasterer, M.C.; Zack, J.A.; Ramasastry, P.; Mumenthaler, S.M.; Kitchen, C.M. CD4+ NK cells can be productively infected with HIV, leading to downregulation of CD4 expression and changes in function. Virology 2009, 387, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Harada, H.; Goto, Y.; Ohno, T.; Suzu, S.; Okada, S. Proliferative activation up-regulates expression of CD4 and HIV-1 co-receptors on NK cells and induces their infection with HIV-1. Eur. J. Immunol. 2007, 37, 2148–2155. [Google Scholar] [CrossRef] [Green Version]
- Lusso, P.; Malnati, M.S.; Garzino-Demo, A.; Crowley, R.W.; Long, E.O.; Gallo, R.C. Infection of natural killer cells by human herpesvirus 6. Nature 1993, 362, 458–462. [Google Scholar] [CrossRef]
- George, L.C.; Rowe, M.; Fox, C.P. Epstein-barr virus and the pathogenesis of T and NK lymphoma: A mystery unsolved. Curr. Hematol. Malig. Rep. 2012, 7, 276–284. [Google Scholar] [CrossRef]
- Trempat, P.; Tabiasco, J.; Andre, P.; Faumont, N.; Meggetto, F.; Delsol, G.; Gascoyne, R.D.; Fournie, J.J.; Vivier, E.; Brousset, P. Evidence for early infection of nonneoplastic natural killer cells by Epstein-Barr virus. J. Virol. 2002, 76, 11139–11142. [Google Scholar] [CrossRef]
- Speck, P.; Haan, K.M.; Longnecker, R. Epstein-Barr virus entry into cells. Virology 2000, 277, 1–5. [Google Scholar] [CrossRef]
- D’Orazio, J.A.; Stein-Streilein, J. Human natural killer (NK) cells present staphylococcal enterotoxin B (SEB) to T lymphocytes. Clin. Exp. Immunol. 1996, 104, 366–373. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.H.; Le, A.M.; Lanier, L.L. Natural killer cells activated in a human mixed lymphocyte response culture identified by expression of Leu-11 and class II histocompatibility antigens. J. Exp. Med. 1984, 159, 993–1008. [Google Scholar] [CrossRef] [Green Version]
- Tabiasco, J.; Vercellone, A.; Meggetto, F.; Hudrisier, D.; Brousset, P.; Fournie, J.J. Acquisition of viral receptor by NK cells through immunological synapse. J. Immunol. 2003, 170, 5993–5998. [Google Scholar] [CrossRef]
- Isobe, Y.; Sugimoto, K.; Yang, L.; Tamayose, K.; Egashira, M.; Kaneko, T.; Takada, K.; Oshimi, K. Epstein-Barr virus infection of human natural killer cell lines and peripheral blood natural killer cells. Cancer Res. 2004, 64, 2167–2174. [Google Scholar] [CrossRef]
- Takahashi, M.; Asabe, S.; Gotanda, Y.; Kishimoto, J.; Tsuda, F.; Okamoto, H. TT virus is distributed in various leukocyte subpopulations at distinct levels, with the highest viral load in granulocytes. Biochem. Biophys. Res. Commun. 2002, 290, 242–248. [Google Scholar] [CrossRef]
- Chivero, E.T.; Bhattarai, N.; Rydze, R.T.; Winters, M.A.; Holodniy, M.; Stapleton, J.T. Human pegivirus RNA is found in multiple blood mononuclear cells in vivo and serum-derived viral RNA-containing particles are infectious in vitro. J. Gen. Virol. 2014, 95, 1307–1319. [Google Scholar] [CrossRef] [Green Version]
- Rice, G.P.; Schrier, R.D.; Oldstone, M.B. Cytomegalovirus infects human lymphocytes and monocytes: Virus expression is restricted to immediate-early gene products. Proc. Natl. Acad. Sci. USA 1984, 81, 6134–6138. [Google Scholar] [CrossRef]
- Hart, D.N.; Baker, B.W.; Inglis, M.J.; Nimmo, J.C.; Starling, G.C.; Deacon, E.; Rowe, M.; Beard, M.E. Epstein-Barr viral DNA in acute large granular lymphocyte (natural killer) leukemic cells. Blood 1992, 79, 2116–2123. [Google Scholar]
- Chiang, A.K.; Tao, Q.; Srivastava, G.; Ho, F.C. Nasal NK- and T-cell lymphomas share the same type of Epstein-Barr virus latency as nasopharyngeal carcinoma and Hodgkin’s disease. Int. J. Cancer 1996, 68, 285–290. [Google Scholar] [CrossRef]
- York, I.A.; Johnson, D.C. Direct contact with herpes simplex virus-infected cells results in inhibition of lymphokine-activated killer cells because of cell-to-cell spread of virus. J. Infect. Dis. 1993, 168, 1127–1132. [Google Scholar] [CrossRef]
- Campbell, T.M.; McSharry, B.P.; Steain, M.; Ashhurst, T.M.; Slobedman, B.; Abendroth, A. Varicella zoster virus productively infects human natural killer cells and manipulates phenotype. PLoS Pathog. 2018, 14, e1006999. [Google Scholar] [CrossRef]
- Lin, S.J.; Cheng, P.J.; Lin, T.Y.; Lee, P.T.; Hsiao, H.S.; Kuo, M.L. Effect of influenza A infection on umbilical cord blood natural killer function regulation with interleukin-15. J. Infect. Dis. 2012, 205, 745–756. [Google Scholar] [CrossRef]
- Mao, H.; Liu, Y.; Sia, S.F.; Peiris, J.S.M.; Lau, Y.L.; Tu, W. Avian influenza virus directly infects human natural killer cells and inhibits cell activity. Virol. Sin. 2017, 32, 122–129. [Google Scholar] [CrossRef]
- Casali, P.; Rice, G.P.; Oldstone, M.B. Viruses disrupt functions of human lymphocytes. Effects of measles virus and influenza virus on lymphocyte-mediated killing and antibody production. J. Exp. Med. 1984, 159, 1322–1337. [Google Scholar] [CrossRef] [Green Version]
- Krzyzaniak, M.A.; Zumstein, M.T.; Gerez, J.A.; Picotti, P.; Helenius, A. Host cell entry of respiratory syncytial virus involves macropinocytosis followed by proteolytic activation of the F protein. PLoS Pathog. 2013, 9, e1003309. [Google Scholar] [CrossRef]
- van Erp, E.A.; Feyaerts, D.; Duijst, M.; Mulder, H.L.; Wicht, O.; Luytjes, W.; Ferwerda, G.; van Kasteren, P.B. Respiratory syncytial virus (RSV) infects primary neonatal and adult natural killer cells and affects their antiviral effector function. J. Infect. Dis. 2019, 219, 723–733. [Google Scholar] [CrossRef]
- Kirwan, S.; Merriam, D.; Barsby, N.; McKinnon, A.; Burshtyn, D.N. Vaccinia virus modulation of natural killer cell function by direct infection. Virology 2006, 347, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Puig, J.M.; Sanchez, L.; Roy, G.; Blasco, R. Susceptibility of different leukocyte cell types to Vaccinia virus infection. Virol. J. 2004, 1, 10. [Google Scholar] [CrossRef]
- Rosenthal, K.L.; Zinkernagel, R.M.; Hengartner, H.; Groscurth, P.; Dennert, G.; Takayesu, D.; Prevec, L. Persistence of vesicular stomatitis virus in cloned interleukin-2-dependent natural killer cell lines. J. Virol. 1986, 60, 539–547. [Google Scholar]
- Valentin, A.; Pavlakis, G.N. Natural killer cells are persistently infected and resistant to direct killing by HIV-1. Anticancer Res. 2003, 23, 2071–2075. [Google Scholar]
- Yamamoto, N.; Okada, M.; Koyanagi, Y.; Kannagi, M.; Hinuma, Y. Transformation of human leukocytes by cocultivation with an adult T cell leukemia virus producer cell line. Science 1982, 217, 737–739. [Google Scholar] [CrossRef]
- Igakura, T.; Stinchcombe, J.C.; Goon, P.K.; Taylor, G.P.; Weber, J.N.; Griffiths, G.M.; Tanaka, Y.; Osame, M.; Bangham, C.R. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science 2003, 299, 1713–1716. [Google Scholar] [CrossRef]
- Lo, K.M.; Vivier, E.; Rochet, N.; Dehni, G.; Levine, H.; Haseltine, W.A.; Anderson, P. Infection of human natural killer (NK) cells with replication-defective human T cell leukemia virus type I provirus. Increased proliferative capacity and prolonged survival of functionally competent NK cells. J. Immunol. 1992, 149, 4101–4108. [Google Scholar]
- Mercer, J.; Helenius, A. Vaccinia virus uses macropinocytosis and apoptotic mimicry to enter host cells. Science 2008, 320, 531–535. [Google Scholar] [CrossRef]
- Welliver, T.P.; Garofalo, R.P.; Hosakote, Y.; Hintz, K.H.; Avendano, L.; Sanchez, K.; Velozo, L.; Jafri, H.; Chavez-Bueno, S.; Ogra, P.L.; et al. Severe human lower respiratory tract illness caused by respiratory syncytial virus and influenza virus is characterized by the absence of pulmonary cytotoxic lymphocyte responses. J. Infect. Dis. 2007, 195, 1126–1136. [Google Scholar] [CrossRef]
- Heltzer, M.L.; Coffin, S.E.; Maurer, K.; Bagashev, A.; Zhang, Z.; Orange, J.S.; Sullivan, K.E. Immune dysregulation in severe influenza. J. Leukoc. Biol. 2009, 85, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, W.M.; Forti, R.L.; Vogler, L.B.; Lawton, A.R.; Gregg, C.R. Spontaneous and interferon resistant natural killer cell anergy in AIDS. AIDS Res. 1983, 1, 221–229. [Google Scholar] [CrossRef]
- Plaeger-Marshall, S.; Spina, C.A.; Giorgi, J.V.; Mitsuyasu, R.; Wolfe, P.; Gottlieb, M.; Beall, G. Alterations in cytotoxic and phenotypic subsets of natural killer cells in acquired immune deficiency syndrome (AIDS). J. Clin. Immunol. 1987, 7, 16–23. [Google Scholar] [CrossRef]
- Scott-Algara, D.; Vuillier, F.; Cayota, A.; Dighiero, G. Natural killer (NK) cell activity during HIV infection: A decrease in NK activity is observed at the clonal level and is not restored after in vitro long-term culture of NK cells. Clin. Exp. Immunol. 1992, 90, 181–187. [Google Scholar] [CrossRef]
- Brenner, B.G.; Dascal, A.; Margolese, R.G.; Wainberg, M.A. Natural killer cell function in patients with acquired immunodeficiency syndrome and related diseases. J. Leukoc. Biol. 1989, 46, 75–83. [Google Scholar] [CrossRef]
- Chivero, E.T.; Bhattarai, N.; McLinden, J.H.; Xiang, J.; Stapleton, J.T. Human Pegivirus (HPgV; formerly known as GBV-C) inhibits IL-12 dependent natural killer cell function. Virology 2015, 485, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Larranaga, C.L.; Ampuero, S.L.; Luchsinger, V.F.; Carrion, F.A.; Aguilar, N.V.; Morales, P.R.; Palomino, M.A.; Tapia, L.F.; Avendano, L.F. Impaired immune response in severe human lower tract respiratory infection by respiratory syncytial virus. Pediatr. Infect. Dis. J. 2009, 28, 867–873. [Google Scholar] [CrossRef]
- Gregoire, C.; Chasson, L.; Luci, C.; Tomasello, E.; Geissmann, F.; Vivier, E.; Walzer, T. The trafficking of natural killer cells. Immunol. Rev. 2007, 220, 169–182. [Google Scholar] [CrossRef]
- Streilein, J.W. Skin-associated lymphoid tissues (SALT): Origins and functions. J. Investig. Dermatol. 1983, 80, 12s–16s. [Google Scholar] [CrossRef]
- Hussell, T.; Openshaw, P.J. Intracellular IFN-gamma expression in natural killer cells precedes lung CD8+ T cell recruitment during respiratory syncytial virus infection. J. Gen. Virol. 1998, 79 Pt 11, 2593–2601. [Google Scholar] [CrossRef]
- Li, F.; Zhu, H.; Sun, R.; Wei, H.; Tian, Z. Natural killer cells are involved in acute lung immune injury caused by respiratory syncytial virus infection. J. Virol. 2012, 86, 2251–2258. [Google Scholar] [CrossRef]
- Wang, J.; Li, F.; Zheng, M.; Sun, R.; Wei, H.; Tian, Z. Lung natural killer cells in mice: Phenotype and response to respiratory infection. Immunology 2012, 137, 37–47. [Google Scholar] [CrossRef]
- Andoniou, C.E.; Coudert, J.D.; Degli-Esposti, M.A. Killers and beyond: NK-cell-mediated control of immune responses. Eur. J. Immunol. 2008, 38, 2938–2942. [Google Scholar] [CrossRef] [Green Version]
- Schuster, I.S.; Coudert, J.D.; Andoniou, C.E.; Degli-Esposti, M.A. “Natural Regulators”: NK Cells as Modulators of T Cell Immunity. Front. Immunol. 2016, 7, 235. [Google Scholar] [CrossRef]


{kind=link}
| Virus | Family | Entry Mechanism | Modulation of NK Cells | Productive Infection | Reference |
|---|---|---|---|---|---|
| Torque teno virus (TTV) | Anelloviridae | Unknown | Unknown | Yes | [31] |
| Human pegivirus (HPgV) | Flaviviridae | Unknown | ↑ Survival ↓ IFN-γ production | Unknown | [32] |
| Cytomegalovirus (CMV) | Herpesviridae | Unknown | Unknown | No | [33] |
| Epstein Barr virus (EBV) | Herpesviridae | Acquisition of receptor after cell–cell interaction Receptor: CD21 Co-receptor: MHC-II | Morphological changes Transformation NK cell malignancies | No | [24,25,29,30,34,35] |
| Herpes simplex virus (HSV) | Herpesviridae | Cell-cell interaction with HSV-infected fibroblasts | Unknown | Unknown | [36] |
| Human herpesvirus 6 (HHV-6) | Herpesviridae | Unknown | ↑ CD4 expression | Unknown | [23] |
| Varicella zoster virus (VZV) | Herpesviridae | Cell-cell interaction with VZV-infected epithelial cells | ↑ CD57 expression ↑ Chemokine receptors ↓ CD56 expression ↓ FcγRIII expression | Yes | [37] |
| Influenza A virus (IAV) | Orthomyxoviridae | Clathrin- and caveolin-dependent endocytosis Receptor: sialic acids | ↑ Apoptosis ↓ Cytotoxicity ↓ Cytotoxicity receptors ↓ Cytokines and chemokines | No | [14,15,18,38,39] |
| Measles virus (MV) | Paramyxoviridae | Unknown | ↓ Cytotoxicity | Unknown | [40] |
| Respiratory syncytial virus (RSV) | Pneumoviridae | Possibly macropinocytosis Receptor: FcγRIIIA (RSV-antibody complexes) | ↑ IFN-γ production ↑ KIR expression ↓ Cytotoxicity ↓Cytotoxicity receptors | No | [41,42] |
| Vaccinia virus (VV) | Poxviridae | Cell-cell interaction | ↑ KIR signaling ↓ Cytotoxicity | No | [43,44] |
| Vesicular stomatitis virus (VSV) | Rhabdoviridae | Unknown | Unknown | Yes | [45] |
| Human immunodeficiency virus 1 (HIV-1) | Retroviridae | Receptor-mediated entry Receptor: CD4 Co-receptors: CXC4/CCR5 | ↑ Apoptosis | Yes | [19,20,21,22,46] |
| Human T-lymphotropic virus (HTLV) | Retroviridae | Cell-cell interaction with T cells | ↑ Proliferation ↑ Survival | Unknown | [47,48,49] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Erp, E.A.; van Kampen, M.R.; van Kasteren, P.B.; de Wit, J. Viral Infection of Human Natural Killer Cells. Viruses 2019, 11, 243. https://doi.org/10.3390/v11030243
van Erp EA, van Kampen MR, van Kasteren PB, de Wit J. Viral Infection of Human Natural Killer Cells. Viruses. 2019; 11(3):243. https://doi.org/10.3390/v11030243
Chicago/Turabian Stylevan Erp, Elisabeth A., Mirjam R. van Kampen, Puck B. van Kasteren, and Jelle de Wit. 2019. "Viral Infection of Human Natural Killer Cells" Viruses 11, no. 3: 243. https://doi.org/10.3390/v11030243
APA Stylevan Erp, E. A., van Kampen, M. R., van Kasteren, P. B., & de Wit, J. (2019). Viral Infection of Human Natural Killer Cells. Viruses, 11(3), 243. https://doi.org/10.3390/v11030243