The Coat Protein of Citrus Yellow Vein Clearing Virus Interacts with Viral Movement Proteins and Serves as an RNA Silencing Suppressor

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Source and Plant Material
2.2. Gene Cloning
2.3. Yeast Two-Hybrid Assays
2.4. BiFC Assay and Subcellular Localization
2.5. Identification of Viral RSS
2.6. Western Blot Analysis
2.7. Small Interference RNA Hybridization
2.8. In Silico Analysis of Protein Sequences
3. Results
3.1. Characterization of CYVCV Proteins
3.2. Interactions of CYVCV Proteins Identified by Yeast Two-Hybrid (Y2H) Assay
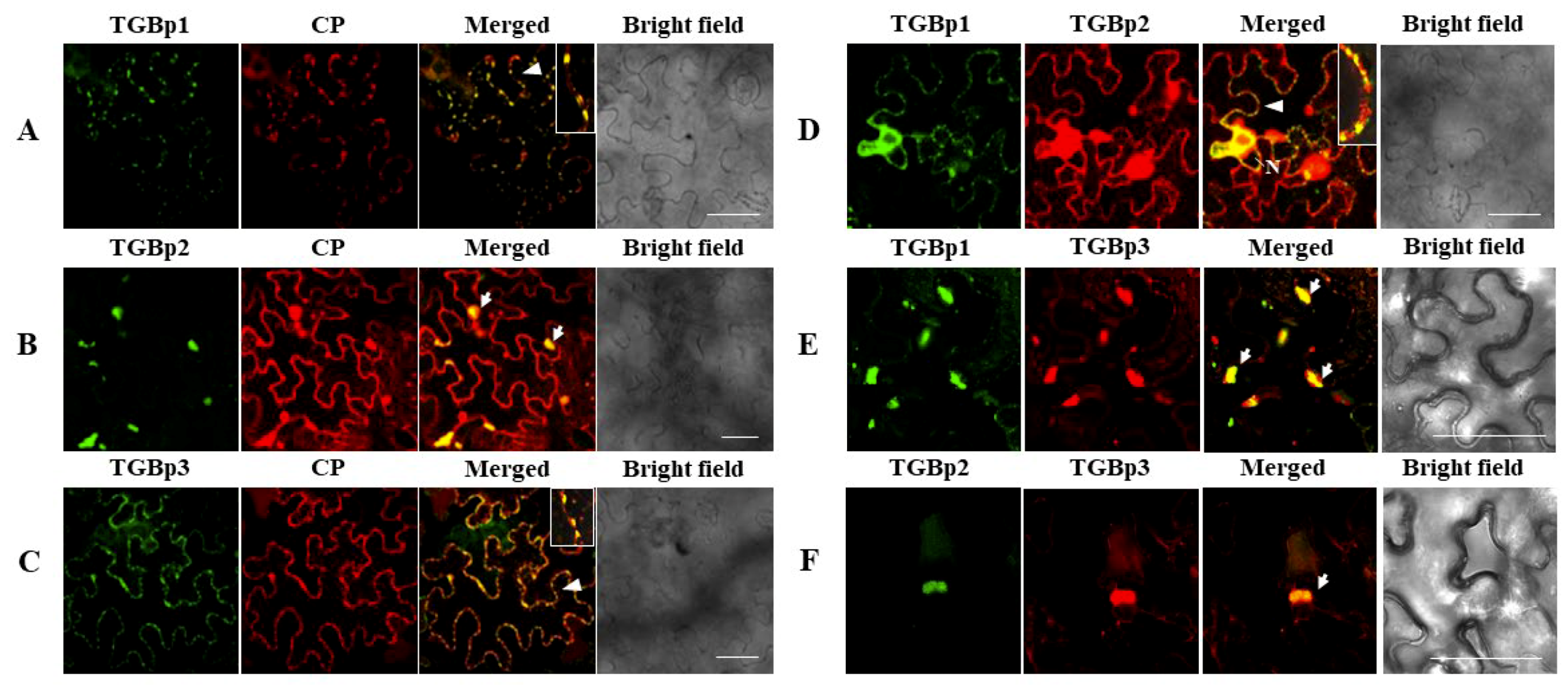
3.3. Intracellular Interactions of CYVCV Proteins
3.4. Subcellular Localization of CYVCV Proteins in N. benthamiana Leaf Cells
3.5. CYVCV CP Functions as an RSS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adams, M.J.; Antoniw, J.F.; Bar-Joseph, M.; Brunt, A.A.; Candresse, T.; Foster, G.D.; Martelli, G.P.; Milne, R.G.; Zavriev, S.K.; Fauquet, C.M. The new plant virus family Flexiviridae and assessment of molecular criteria for species demarcation. Arch. Virol. 2004, 149, 1045–1060. [Google Scholar] [CrossRef]
- Adams, M.J.; Antoniw, J.F.; Kreuze, J. Virgaviridae: A new family of rod-shaped plant viruses. Arch. Virol. 2009, 154, 1967–1972. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.L.; Hung, Y.J.; Tseng, Y.H.; Hsu, H.T.; Yang, J.Y.; Wung, C.H.; Lin, N.S.; Meng, M.; Hsu, Y.H.; Chang, B.Y. The stable association of virion with the triple-gene-block protein 3-based complex of bamboo mosaic virus. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Morozov, S.Y.; Solovyev, A.G. Triple gene block: Modular design of a multifunctional machine for plant virus movement. J. Gen. Virol. 2003, 84, 1351–1366. [Google Scholar] [CrossRef]
- Verchot-Lubicz, J. A new cell-to-cell transport model for potexviruses. Mol. Plant-Microbe Interact. 2005, 18, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Bragg, J.N.; Ganesan, U.; Ruzin, S.; Schichnes, D.; Lee, M.Y.; Vaira, A.M.; Ryu, K.H.; Hammond, J.; Jackson, A.O. Subcellular localization of the barley stripe mosaic virus triple gene block proteins. J. Virol. 2009, 83, 9432–9448. [Google Scholar] [CrossRef]
- Jackson, A.O.; Lim, H.S.; Bragg, J.; Ganesan, U.; Lee, M.Y. Hordeivirus replication, movement, and pathogenesis. Annu. Rev. Phytopathol. 2009, 47, 385–422. [Google Scholar] [CrossRef]
- Park, M.R.; Jeong, R.D.; Kim, K.H. Understanding the intracellular trafficking and intercellular transport of potexviruses in their host plants. Front. Plant Sci. 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Makarov, V.V.; Rybakova, E.N.; Efimov, A.V.; Dobrov, E.N.; Serebryakova, M.V.; Solovyev, A.G.; Yaminsky, I.V.; Taliansky, M.E.; Morozov, S.Y.; Kalinina, N.O. Domain organization of the N-terminal portion of hordeivirus movement protein TGBp1. J. Gen. Virol. 2009, 90, 3022–3032. [Google Scholar] [CrossRef]
- Liou, M.R.; Hu, C.C.; Chou, Y.L.; Chang, B.Y.; Lin, N.S.; Hsu, Y.H. Viral elements and host cellular proteins in intercellular movement of bamboo mosaic virus. Curr. Opin. Virol. 2015, 12, 99–108. [Google Scholar] [CrossRef]
- Chang, B.Y.; Lin, N.S.; Liou, D.Y.; Chen, J.P.; Liou, G.G.; Hsu, Y.H. Subcellular localization of the 28 kDa protein of the triple-gene-block of bamboo mosaic potexvirus. J. Gen. Virol. 1997, 78, 1175–1179. [Google Scholar] [CrossRef]
- Samuels, T.D.; Ju, H.J.; Ye, C.M.; Motes, C.M.; Blancaflor, E.B.; Verchot-Lubicz, J. Subcellular targeting and interactions among the potato virus X TGB proteins. Virology 2007, 367, 375–389. [Google Scholar] [CrossRef]
- Kalinina, N.O.; Rakitina, D.V.; Solovyev, A.G.; Schiemann, J.; Morozov, S.Y. RNA helicase activity of the plant virus movement proteins encoded by the first gene of the triple gene block. Virology 2002, 296, 321–329. [Google Scholar] [CrossRef]
- Wung, C.H.; Hsu, Y.H.; Liou, D.Y.; Huang, W.C.; Lin, N.S.; Chang, B.Y. Identification of the RNA-binding sites of the triple gene block protein 1 of bamboo mosaic potexvirus. J. Gen. Virol. 1999, 80, 1119–1126. [Google Scholar] [CrossRef]
- Liou, D.Y.; Hsu, Y.H.; Wung, C.H.; Wang, W.H.; Lin, N.S.; Chang, B.Y. Functional analyses and identification of two arginine residues essential to the ATP-utilizing activity of the triple gene block protein 1 of bamboo mosaic potexvirus. Virology 2000, 277, 336–344. [Google Scholar] [CrossRef]
- Lin, M.K.; Chang, B.Y.; Liao, J.T.; Lin, N.S.; Hsu, Y.H. Arg-16 and Arg-21 in the N-terminal region of the triple-gene-block protein 1 of bamboo mosaic virus are essential for virus movement. J. Gen. Virol. 2004, 85, 251–259. [Google Scholar] [CrossRef]
- Angell, S.M.; Davies, C.; Baulcombe, D.C. Cell-to-cell movement of potato virus X is associated with a change in the size-exclusion limit of plasmodesmata in trichome cells of Nicotiana clevelandii. Virology 1996, 216, 197–201. [Google Scholar] [CrossRef]
- Howard, A.R.; Heppler, M.L.; Ju, H.J.; Krishnamurthy, K.; Payton, M.E.; Verchot-Lubicz, J. Potato virus X TGBp1 induces plasmodesmata gating and moves between cells in several host species whereas CP moves only in N. benthamiana leaves. Virology 2004, 328, 185–197. [Google Scholar] [CrossRef]
- Atabekov, J.G.; Rodionova, N.P.; Karpova, O.V.; Kozlovsky, S.V.; Poljakov, V.Y. The movement protein-triggered in situ conversion of potato virus X virion RNA from a nontranslatable into a translatable form. Virology 2000, 271, 259–263. [Google Scholar] [CrossRef]
- Rodionova, N.P.; Karpova, O.V.; Kozlovsky, S.V.; Zayakina, O.V.; Arkhipenko, M.V.; Atabekov, J.G. Linear remodeling of helical virus by movement protein binding. J. Mol. Biol. 2003, 333, 565–572. [Google Scholar] [CrossRef]
- Solovyev, A.G.; Stroganova, T.A.; Zamyatnin, A.A.; Fedorkin, O.N.; Schiemann, J.; Morozov, S.Y. Subcellular sorting of small membrane-associated triple gene block proteins: TGBp3-assisted targeting of TGBp2. Virology 2000, 269, 113–127. [Google Scholar] [CrossRef]
- Hsu, H.T.; Chou, Y.L.; Tseng, Y.H.; Lin, Y.H.; Lin, T.M.; Lin, N.S.; Hsu, Y.H.; Chang, B.Y. Topological properties of the triple gene block protein 2 of bamboo mosaic virus. Virology 2008, 379, 1–9. [Google Scholar] [CrossRef]
- Bamunusinghe, D.; Hemenway, C.L.; Nelson, R.S.; Sanderfoot, A.A.; Ye, C.M.; Silva, M.A.T.; Payton, M.; Verchot-Lubicz, J. Analysis of potato virus X replicase and TGBp3 subcellular locations. Virology 2009, 393, 272–285. [Google Scholar] [CrossRef]
- Mathioudakis, M.M.; Rodríguez-Moreno, L.; Sempere, R.N.; Aranda, M.A.; Livieratos, I. Multifaceted capsid proteins: multiple interactions suggest multiple roles for pepino mosaic virus capsid protein. Mol. Plant. Microbe. Interact. 2014, 27, 1356–1369. [Google Scholar] [CrossRef]
- Yang, Y.; Ding, B.; Baulcombe, D.C.; Verchot, J. Cell-to-cell movement of the 25K protein of potato virus X is regulated by three other viral proteins. Mol. Plant. Microbe. Interact. 2000, 13, 599–605. [Google Scholar] [CrossRef]
- Wu, C.H.; Lee, S.C.; Wang, C.W. Viral protein targeting to the cortical endoplasmic reticulum is required for cell-cell spreading in plants. J. Cell Biol. 2011, 193, 521–535. [Google Scholar] [CrossRef]
- Lee, S.C.; Wu, C.H.; Wang, C.W. Traffic of a viral movement protein complex to the highly curved tubules of the cortical endoplasmic reticulum. Traffic 2010, 11, 912–930. [Google Scholar] [CrossRef]
- Lu, H.C.; Chen, C.E.; Tsai, M.H.; Wang, H.I.; Su, H.J.; Yeh, H.H. Cymbidium mosaic potexvirus isolate-dependent host movement systems reveal two movement control determinants and the coat protein is the dominant. Virology 2009, 388, 147–159. [Google Scholar] [CrossRef]
- Weber, P.H.; Bujarski, J.J. Multiple functions of capsid proteins in (+) stranded RNA viruses during plant-virus interactions. Virus Res. 2015, 196, 140–149. [Google Scholar] [CrossRef]
- Qu, F.; Ren, T.; Morris, T.J. The coat protein of turnip crinkle virus suppresses post transcriptional gene silencing at an early initiation step. J. Virol. 2003, 77, 511–522. [Google Scholar] [CrossRef]
- Pérez-Cañamás, M.; Hernández, C. Key importance of small RNA binding for the activity of a glycine-tryptophan (GW) motif-containing viral suppressor of RNA silencing. J. Biol. Chem. 2015, 290, 3106–3120. [Google Scholar] [CrossRef]
- Bayne, E.H.; Rakitina, D.V.; Morozov, S.Y.; Baulcombe, D.C. Cell-to-cell movement of potato potexvirus X is dependent on suppression of RNA silencing. Plant J. 2005, 44, 471–482. [Google Scholar] [CrossRef]
- Voinnet, O.; Lederer, C.; Baulcombe, D.C. A viral movement protein prevents spread of the gene silencing signal in Nicotiana benthamiana. Cell 2000, 103, 157–167. [Google Scholar] [CrossRef]
- Senshu, H.; Ozeki, J.; Komatsu, K.; Hashimoto, M.; Hatada, K.; Aoyama, M.; Kagiwada, S.; Yamaji, Y.; Namba, S. Variability in the level of RNA silencing suppression caused by triple gene block protein 1 (TGBp1) from various potexviruses during infection. J. Gen. Virol. 2009, 90, 1014–1024. [Google Scholar] [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Carstens, E.B. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses. Arch. Virol. 2014, 159, 2831–2841. [Google Scholar] [CrossRef]
- Chen, H.M.; Li, Z.A.; Wang, X.F.; Zhou, Y.; Tang, K.Z.; Zhou, C.Y.; Zhao, X.Y.; Yue, J.Q. First report of citrus yellow vein clearing virus on lemon in Yunnan, China. Plant Dis. 2014, 98, 1747. [Google Scholar] [CrossRef]
- Loconsole, G.; Önelge, N.; Potere, O.; Giampetruzzi, A.; Bozan, O.; Satar, S.; De Stradis, A.; Savino, V.; Yokomi, R.K.; Saponari, M. Identification and characterization of citrus yellow vein clearing virus, a putative new member of the genus Mandarivirus. Phytopathology 2012, 102, 1168–1175. [Google Scholar] [CrossRef]
- Cao, M.J.; Wu, Q.; Atta, S.; Su, H.N.; Yu, Y.Q.; Chen, H.M.; Zhou, C.Y. First molecular evidence of citrus yellow vein clearing virus from citrus in Punjab, Pakistan. Plant Dis. 2016, 100, 540. [Google Scholar] [CrossRef]
- Zhen, S.; Kurth, E.G.; Peremyslov, V.V.; Changyong, Z.; Dolja, V.V. Molecular characterization of a citrus yellow vein clearing virus strain from China. Arch. Virol. 2015, 160, 1811–1813. [Google Scholar] [CrossRef]
- Yu, Y.; Wu, Q.; Su, H.; Wang, X.; Cao, M.; Zhou, C. Small RNA deep sequencing reveals full-length genome of citrus yellow vein clearing virus in Chongqing, China. J. Integr. Agric. 2017, 16, 503–508. [Google Scholar] [CrossRef]
- Feng, Z.; Xue, F.; Xu, M.; Chen, X.; Zhao, W.; Garcia-Murria, M.J.; Mingarro, I.; Liu, Y.; Huang, Y.; Jiang, L.; Zhu, M.; Tao, X. The ER-membrane transport system is critical for intercellular trafficking of the NSm movement protein and tomato spotted wilt tospovirus. PLoS Pathog. 2016, 12, 1–27. [Google Scholar] [CrossRef]
- Kreuze, J.F.; Savenkov, E.I.; Cuellar, W.; Li, X.; Valkonen, J.P.T. Viral class 1 RNase III involved in suppression of RNA silencing. J. Virol. 2005, 79, 7227–7238. [Google Scholar] [CrossRef]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef]
- Kelley, L.A.; Sternberg, M.J.E. Protein structure prediction on the Web: a case study using the Phyre server (http://www.sbg.bio.ic.ac.uk/phyre2). Nat. Protoc. 2009, 4, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Hessa, T.; Meindl-Beinker, N.M.; Bernsel, A.; Kim, H.; Sato, Y.; Lerch-Bader, M.; Nilsson, I.; White, S.H.; von Heijne, G. Molecular code for transmembrane-helix recognition by the Sec61 translocon (http://dgpred.cbr.su.se/). Nature 2007, 450, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Tsirigos, K.D.; Peters, C.; Shu, N.; Käll, L.; Elofsson, A. The TOPCONS web server for consensus prediction of membrane protein topology and signal peptides (http://topcons.net/). Nucleic Acids Res. 2015, 43, W401–W407. [Google Scholar] [CrossRef]
- Voinnet, O. RNA silencing: small RNAs as ubiquitous regulators of gene expression. Curr. Opin. Plant Biol. 2002, 5, 444–451. [Google Scholar] [CrossRef]
- Torrance, L.; Wright, K.M.; Crutzen, F.; Cowan, G.H.; Lukhovitskaya, N.I.; Bragard, C.; Savenkov, E.I. Unusual features of pomoviral RNA movement. Front. Microbiol. 2011, 2, 259. [Google Scholar] [CrossRef]
- Verchot-Lubicz, J.; Torrance, L.; Solovyev, A.G.; Morozov, S.Y.; Jackson, A.O.; Gilmer, D. Varied movement strategies employed by triple gene block–encoding viruses. Mol. Plant-Microbe Interact. 2010, 23, 1231–1247. [Google Scholar] [CrossRef] [PubMed]
- Leshchiner, A.D.; Minina, E.A.; Rakitina, D.V.; Vishnichenko, V.K.; Solovyev, A.G.; Morozov, S.Y.; Kalinina, N.O. Oligomerization of the potato virus X 25-kD movement protein. Biochemistry (Mosc.) 2008, 73, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, K.; Heppler, M.; Mitra, R.; Blancaflor, E.; Payton, M.; Nelson, R.S.; Verchot-Lubicz, J. The potato virus X TGBp3 protein associates with the ER network for virus cell-to-cell movement. Virology 2003, 309, 135–151. [Google Scholar] [CrossRef]
- Sempere, R.N.; Gómez, P.; Truniger, V.; Aranda, M.A. Development of expression vectors based on pepino mosaic virus. Plant Methods 2011, 7, 6. [Google Scholar] [CrossRef]
- Davies, C.; Hills, G.; Baulcombe, D.C. Sub-cellular localization of the 25-kDa protein encoded in the triple gene block of potato virus X. Virology 1993, 197, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, A.R.; Niewiadomski, S.; Prosser, S.W.; Krell, P.; Meng, B. Subcellular localization of the triple gene block proteins encoded by a foveavirus infecting grapevines. Virus Res. 2008, 138, 57–69. [Google Scholar] [CrossRef]
- Zamyatnin, A.A.; Solovyev, A.G.; Savenkov, E.I.; Germundsson, A.; Sandgren, M.; Valkonen, J.P.T.; Morozov, S.Y. Transient coexpression of individual genes encoded by the triple gene block of potato mop-top virus reveals requirements for TGBp1 trafficking. Mol. Plant. Microbe. Interact. 2004, 17, 921–930. [Google Scholar] [CrossRef]
- Lawrence, D.M.; Jackson, A.O. Interactions of the TGB1 protein during cell-to-cell movement of barley stripe mosaic virus. J. Virol. 2001, 75, 8712–8723. [Google Scholar] [CrossRef]
- Lucas, W.J. Plant viral movement proteins: agents for cell-to-cell trafficking of viral genomes. Virology 2006, 344, 169–184. [Google Scholar] [CrossRef]
- Bamunusinghe, D.; Seo, J.K.; Rao, A.L.N. Subcellular localization and rearrangement of endoplasmic reticulum by brome mosaic virus capsid protein. J. Virol. 2011, 85, 2953–2963. [Google Scholar] [CrossRef]
- Cheng, C.P. Host factors involved in the intracellular movement of bamboo mosaic virus. Front. Microbiol. 2017, 8, 759. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.S.; Ju, H.J. The plant cellular systems for plant virus movement. Plant Pathol. J. 2017, 33, 213–228. [Google Scholar] [CrossRef]
- Schepetilnikov, M.V.; Manske, U.; Solovyev, A.G.; Zamyatnin, A.A.; Schiemann, J.; Morozov, S.Y. The hydrophobic segment of potato virus X TGBp3 is a major determinant of the protein intracellular trafficking. J. Gen. Virol. 2005, 86, 2379–2391. [Google Scholar] [CrossRef] [PubMed]
- DeBlasio, S.L.; Xu, Y.; Johnson, R.S.; Rebelo, A.R.; MacCoss, M.J.; Gray, S.M.; Heck, M. The interaction dynamics of two potato leafroll virus movement proteins affects their localization to the outer membranes of mitochondria and plastids. Viruses 2018, 10, 585. [Google Scholar] [CrossRef]
- Chapman, S.; Hills, G.; Watts, J.; Baulcombe, D. Mutational analysis of the coat protein gene of potato virus X: effects on virion morphology and viral pathogenicity. Virology 1992, 191, 223–230. [Google Scholar] [CrossRef]
- Forster, R.L.; Beck, D.L.; Guilford, P.J.; Voot, D.M.; Van Dolleweerd, C.J.; Andersen, M.T. The coat protein of white clover mosaic potexvirus has a role in facilitating cell-to-cell transport in plants. Virology 1992, 191, 480–484. [Google Scholar] [CrossRef]
- Cui, T.; Bin, Y.; Yan, J.; Mei, P.; Li, Z.; Zhou, C.; Song, Z. Development of infectious cDNA clones of citrus yellow vein clearing virus using a novel and rapid strategy. Phytopathology 2018, 108, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. RNA silencing as a plant immune system against viruses. Trends Genet. 2001, 17, 449–459. [Google Scholar] [CrossRef]
- Csorba, T.; Burgyán, J. Antiviral silencing and suppression of gene silencing in plants. In Current Research Topics in Plant Virology; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–33. [Google Scholar]
- Himber, C.; Dunoyer, P.; Moissiard, G.; Ritzenthaler, C.; Voinnet, O. Transitivity-dependent and -independent cell-to-cell movement of RNA silencing. EMBO J. 2003, 22, 4523–4533. [Google Scholar] [CrossRef]
- Ma, X.; Hong, N.; Moffett, P.; Zhou, Y.; Wang, G. Functional analysis of apple stem pitting virus coat protein variants. Virol. J. 2019, 16. [Google Scholar] [CrossRef]
- Okano, Y.; Senshu, H.; Hashimoto, M.; Neriya, Y.; Netsu, O.; Minato, N.; Yoshida, T.; Maejima, K.; Oshima, K.; Komatsu, K.; Yamaji, Y.; Namba, S. In planta recognition of a double-stranded RNA synthesis protein complex by a potexviral RNA silencing suppressor. Plant Cell 2014, 26, 2168–2183. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rehman, A.U.; Li, Z.; Yang, Z.; Waqas, M.; Wang, G.; Xu, W.; Li, F.; Hong, N. The Coat Protein of Citrus Yellow Vein Clearing Virus Interacts with Viral Movement Proteins and Serves as an RNA Silencing Suppressor. Viruses 2019, 11, 329. https://doi.org/10.3390/v11040329
Rehman AU, Li Z, Yang Z, Waqas M, Wang G, Xu W, Li F, Hong N. The Coat Protein of Citrus Yellow Vein Clearing Virus Interacts with Viral Movement Proteins and Serves as an RNA Silencing Suppressor. Viruses. 2019; 11(4):329. https://doi.org/10.3390/v11040329
Chicago/Turabian StyleRehman, Atta Ur, Zhuoran Li, Zuokun Yang, Muhammad Waqas, Guoping Wang, Wenxing Xu, Feng Li, and Ni Hong. 2019. "The Coat Protein of Citrus Yellow Vein Clearing Virus Interacts with Viral Movement Proteins and Serves as an RNA Silencing Suppressor" Viruses 11, no. 4: 329. https://doi.org/10.3390/v11040329
APA StyleRehman, A. U., Li, Z., Yang, Z., Waqas, M., Wang, G., Xu, W., Li, F., & Hong, N. (2019). The Coat Protein of Citrus Yellow Vein Clearing Virus Interacts with Viral Movement Proteins and Serves as an RNA Silencing Suppressor. Viruses, 11(4), 329. https://doi.org/10.3390/v11040329