Experimental Infection of Sand Flies by Massilia Virus and Viral Transmission by Co-Feeding on Sugar Meal

 ,
, 

Abstract
:1. Introduction
2. Material and Methods
2.1. Sand Fly Colonies
2.2. Virus Strain
2.3. Different Infection Routes for MASV Infection in Sand Flies
2.4. MASV Detection in Experimentally Infected Sand Flies
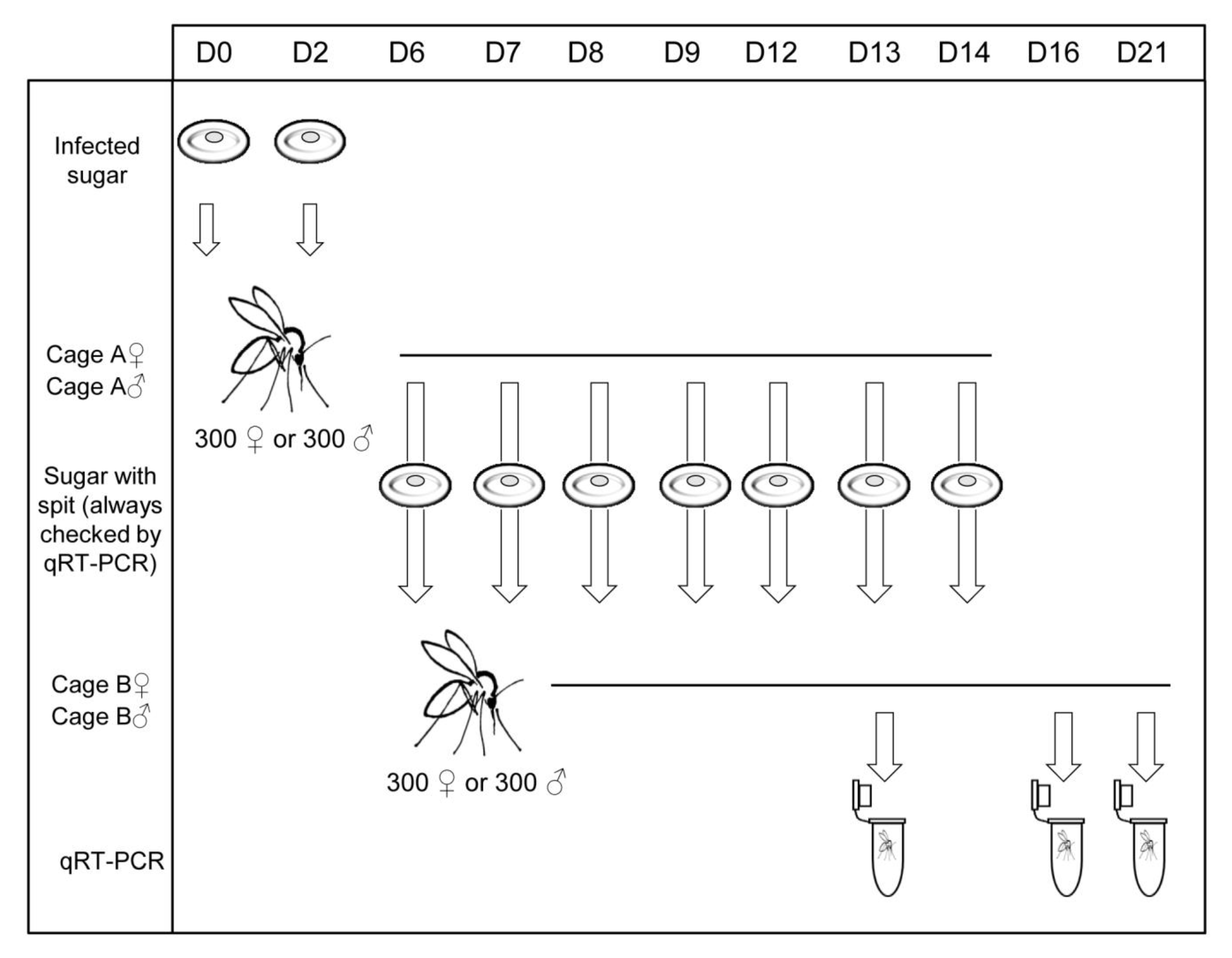
2.5. Co-Feeding Experiments (CoF)
2.6. Susceptibility of Various Sand Fly Species to MASV
2.7. Tissue Specificity of MASV
2.8. Statistical Analysis
3. Results
3.1. Infection of P. perniciosus L1 Using Larval Food
3.2. Infections of P. perniciosus L4 by Larval Food
3.3. P. perniciosus Females Infected by Bloodmeal
3.4. P. perniciosus Males and Females Infected by Sugar Meal
3.5. Co-Feeding Experiments
3.6. Susceptibility of Various Sand Fly Species to MASV
3.7. Tissue Specificity of MASV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Killick-Kendrick, R. The biology and control of phlebotomine sand flies. Clin. Dermatol. 1999, 17, 279–289. [Google Scholar] [CrossRef]
- Maroli, M.; Feliciangeli, M.D.; Bichaud, L.; Charrel, R.N.; Gradoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med. Vet. Entomol. 2013, 27, 123–147. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Bichaud, L.; de Lamballerie, X.; Alten, B.; Gould, E.A.; Charrel, R.N. Sandfly-borne phleboviruses of Eurasia and Africa: Epidemiology, genetic diversity, geographic range, control measures. Antivir. Res. 2013, 100, 54–74. [Google Scholar] [CrossRef] [Green Version]
- Maes, P.; Adkins, S.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, E.; Blair, C.D.; Briese, T.; et al. Taxonomy of the order Bunyavirales: Second update 2018. Arch. Virol. 2019, 164, 927–941. [Google Scholar] [CrossRef] [PubMed]
- Hubalek, Z.; Rudolf, I. Tick-borne viruses in Europe. Parasitol. Res. 2012, 111, 9–36. [Google Scholar] [CrossRef] [PubMed]
- Horne, K.M.; Vanlandingham, D.L. Bunyavirus-vector interactions. Viruses 2014, 6, 4373–4397. [Google Scholar] [CrossRef] [PubMed]
- Charrel, R.N.; Gallian, P.; Navarro-Mari, J.M.; Nicoletti, L.; Papa, A.; Sanchez-Seco, M.P.; Tenorio, A.; de Lamballerie, X. Emergence of Toscana virus in Europe. Emerg. Infect. Dis. 2005, 11, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Charrel, R.N.; Bichaud, L.; de Lamballerie, X. Emergence of Toscana virus in the mediterranean area. World J. Virol. 2012, 1, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Es-sette, N.; Ajaoud, M.; Anga, L.; Mellouki, F.; Lemrani, M. Toscana virus isolated from sandflies, Morocco. Parasit. Vectors 2015, 8, 205. [Google Scholar] [CrossRef] [PubMed]
- Charrel, R.N.; Izri, A.; Temmam, S.; de Lamballerie, X.; Parola, P. Toscana virus RNA in Sergentomyia minuta files. Emerg. Infect. Dis. 2006, 12, 1299–1300. [Google Scholar] [CrossRef]
- Ergunay, K.; Kasap, O.E.; Orsten, S.; Oter, K.; Gunay, F.; Yoldar, A.Z.; Dincer, E.; Ozkul, A. Phlebovirus and Leishmania detection in sandflies from eastern Thrace and northern Cyprus. Parasit. Vectors 2014, 7, 1–13. [Google Scholar] [CrossRef]
- Ayhan, N.; Alten, B.; Ivovic, V.; Martinkovic, F.; Kasap, O.E.; Ozbel, Y.; de Lamballerie, X.; Charrel, R.N. Cocirculation of Two Lineages of Toscana Virus in Croatia. Front. Public Health 2017, 12, 336. [Google Scholar] [CrossRef]
- Es-sette, N.; Ajaoud, M.; Bichaud, L.; Hamdi, S.; Mellouki, F.; Charrel, R.N.; Lemrani, M. Phlebotomus sergenti a common vector of Leishmania tropica and Toscana virus in Morocco. J. Vector Borne Dis. 2014, 51, 86. [Google Scholar]
- Charrel, R.N.; Moureau, G.; Temmam, S.; Izri, A.; Marty, P.; Parola, P.; da Rosa, A.T.; Tesh, R.B.; de Lamballerie, X. Massilia virus, a novel Phlebovirus (Bunyaviridae) isolated from sandflies in the Mediterranean. Vector Borne Zoonotic Dis. 2009, 9, 519–530. [Google Scholar] [CrossRef]
- Tesh, R.B.; Chaniotis, B.N. Transovarial transmission of viruses by phlebotomine sandflies. Ann. N. Y. Acad. Sci. 1975, 266, 125–134. [Google Scholar] [CrossRef]
- Tesh, R.B.; Modi, G.B. Maintenance of Toscana virus in Phlebotomus perniciosus by vertical transmission. Am. J. Trop. Med. Hyg. 1987, 36, 189–193. [Google Scholar] [CrossRef]
- Tesh, R.B.; Lubroth, J.; Guzman, H. Simulation of arbovirus overwintering: Survival of Toscana virus (Bunyaviridae: Phlebovirus) in its natural sand fly vector Phlebotomus perniciosus. Am. J. Trop. Med. Hyg. 1992, 47, 574–581. [Google Scholar] [CrossRef]
- Maroli, M.; Ciufolini, M.G.; Verani, P. Vertical transmission of Toscana virus in the sandfly, Phlebotomus perniciosus, via the second gonotrophic cycle. Med. Vet. Entomol. 1993, 7, 283–286. [Google Scholar] [CrossRef]
- Labuda, M.; Nuttall, P.A.; Kožuch, O.; Elečková, E.; Williams, T.; Žuffová, E.; Sabo, A. Non-viraemic transmission of tick-borne encephalitis virus: A mechanism for arbovirus survival in nature. Experientia 1993, 49, 802–805. [Google Scholar] [CrossRef]
- Higgs, S.; Schneider, B.S.; Vanlandingham, D.L.; Klingler, K.A.; Gould, E.A. Nonviremic transmission of West Nile virus. Proc. Natl. Acad. Sci. USA 2005, 102, 8871–8874. [Google Scholar] [CrossRef] [Green Version]
- Mead, D.G.; Ramberg, F.B.; Besselsen, D.G.; Maré, C.J. Transmission of vesicular stomatitis virus from infected to noninfected black flies co-feeding on nonviremic deer mice. Science 2000, 287, 485–487. [Google Scholar] [CrossRef]
- Volf, P.; Volfova, V. Establishment and maintenance of sand fly colonies. J. Vector Ecol. 2011, 36, 1–9. [Google Scholar] [CrossRef]
- Alwassouf, S.; Maia, C.; Ayhan, N.; Coimbra, M.; Cristovao, J.M.; Richet, H.; Bichaud, L.; Campino, L.; Charrel, R.N. Neutralization-based seroprevalence of Toscana virus and sandfly fever Sicilian virus in dogs and cats from Portugal. J. Gen. Virol. 2016, 97, 2816–2823. [Google Scholar] [CrossRef]
- Dincer, E.; Gargari, S.; Ozkul, A.; Ergunay, K. Potential animal reservoirs of Toscana virus and coinfections with Leishmania infantum in Turkey. Am. J. Trop. Med. Hyg. 2015, 92, 690–697. [Google Scholar] [CrossRef]
- Möckel, N.; Gisder, S.; Genersch, E. Horizontal transmission of deformed wing virus: Pathological consequences in adult bees (Apis mellifera) depend on the transmission route. J. Gen. Virol. 2011, 92, 370–377. [Google Scholar] [CrossRef]
- Ferreira, Á.G.; Naylor, H.; Esteves, S.S.; Pais, I.S.; Martins, N.E.; Teixeira, L. The Toll-dorsal pathway is required for resistance to viral oral infection in Drosophila. PLoS Pathog. 2014, 10, e1004507. [Google Scholar] [CrossRef] [PubMed]
- Stevanovic, A.L.; Johnson, K.N. Infectivity of Drosophila C virus following oral delivery in Drosophila larvae. J. Gen. Virol. 2015, 96, 1490–1496. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.L.; Houk, E.J.; Kramer, L.D.; Reeves, W.C. Intrinsic factors affecting vector competence of mosquitoes for arboviruses. Annu. Rev. Entomol. 1983, 28, 229–262. [Google Scholar] [CrossRef]
- Franz, A.W.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue barriers to arbovirus infection in mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef]
- Sudeep, A.B.; Mandar, P.; Ghodke, Y.K.; George, R.P.; Gokhale, M.D. Vector competence of two Indian populations of Culex quinquefasciatus (Diptera: Culicidae) mosquitoes to three West Nile virus strains. J. Vector Borne Dis. 2015, 52, 185. [Google Scholar]
- Diallo, M.; Thonnon, J.; Fontenille, D. Vertical transmission of the yellow fever virus by Aedes aegypti (Diptera, Culicidae): Dynamics of infection in F1 adult progeny of orally infected females. Am. J. Trop. Med. Hyg. 2000, 62, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Wasinpiyamongkol, L.; Thongrungkiat, S.; Jirakanjanakit, N.; Apiwathnasorn, C. Susceptibility and transovarial transmission of dengue virus in Aedes aegypti: A preliminary study of morphological variations. Southeast Asian J. Trop. Med. Public Health 2003, 34, 131–135. [Google Scholar]
- Castro, M.G.D.; Nogueira, R.M.R.; Schatzmayr, H.G.; Miagostovich, M.P.; Lourenço-de-Oliveira, R. Dengue virus detection by using reverse transcription-polymerase chain reaction in saliva and progeny of experimentally infected Aedes albopictus from Brazil. Memórias Inst. Oswaldo Cruz 2004, 99, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Saiyasombat, R.; Bolling, B.G.; Brault, A.C.; Bartholomay, L.C.; Blitvich, B.J. Evidence of efficient transovarial transmission of Culex flavivirus by Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2011, 48, 1031–1038. [Google Scholar] [CrossRef]
- Volf, P.; Kiewegová, A.; Nemec, A. Bacterial colonisation in the gut of Phlebotomus duboscqi (Diptera: Psychodidae): Transtadial passage and the role of female diet. Folia Parasitol. 2002, 49, 73–77. [Google Scholar] [CrossRef]
- Chavshin, A.R.; Oshaghi, M.A.; Vatandoost, H.; Yakhchali, B.; Zarenejad, F.; Terenius, O. Malpighian tubules are important determinants of Pseudomonas transstadial transmission and longtime persistence in Anopheles stephensi. Parasit. Vector 2015, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Hakim, R.S.; Baldwin, K.; Smagghe, G. Regulation of midgut growth, development, and metamorphosis. Ann. Rev. Entomol. 2010, 55, 593–608. [Google Scholar] [CrossRef]
- Fernandes, K.M.; Neves, C.A.; Serrão, J.E.; Martins, G.F. Aedes aegypti midgut remodeling during metamorphosis. Parasitol. Int. 2014, 63, 506–512. [Google Scholar] [CrossRef]
- Chavshin, A.R.; Oshaghi, M.A.; Vatandoost, H.; Yakhchali, B.; Raeisi, A.; Zarenejad, F. Escherichia coli expressing a green fluorescent protein (GFP) in Anopheles stephensi: A preliminary model for paratransgenesis. Symbiosis 2013, 60, 17–24. [Google Scholar] [CrossRef]
- Terra, W.R. Evolution of digestive systems of insects. Ann. Rev. Entomol. 1990, 35, 181–200. [Google Scholar] [CrossRef]
- Peters, W. Peritrophic Membranes; Springer Press: Berlin/Heidelberg, Germany, 1992; Volume 2, pp. 1–238. ISBN 978-3-642-84416-4. [Google Scholar]
- Lehane, M.J. Peritrophic matrix structure and function. Ann. Rev. Entomol. 1997, 42, 525–550. [Google Scholar] [CrossRef]
- Pruzinova, K.; Sadlova, J.; Seblova, V.; Homola, M.; Votypka, J.; Volf, P. Comparison of bloodmeal digestion and the peritrophic matrix in four sand fly species differing in susceptibility to Leishmania donovani. PLoS ONE 2015, 10, e0128203. [Google Scholar] [CrossRef]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Souza-Neto, J.; Cosme, R.T.; Rovira, J.; Ortiz, A.; Pascale, J.M.; Dimopoulos, G. Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl. Trop. Dis. 2012, 6, e1561. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Short, S.M.; Bahia, A.C.; Saraiva, R.G.; Dong, Y.; Kang, S.; Tripathi, A.; Mlambo, G.; Dimopoulos, G. Chromobacterium Csp_P reduces malaria and dengue infection in vector mosquitoes and has entomopathogenic and in vitro anti-pathogen activities. PLoS Pathog. 2014, 10, e1004398. [Google Scholar] [CrossRef]
- Tang, Y.; Ward, R.D. Sugar feeding and fluid destination control in the phlebotomine sandfly Lutzomyia longipalpis (Diptera: Psychodidae). Med. Vet. Entomol. 1998, 12, 13–19. [Google Scholar] [CrossRef]
- Van den Hurk, A.F.; Johnson, P.H.; Hall-Mendelin, S.; Northill, J.A.; Simmons, R.J.; Jansen, C.C.; Frances, S.P.; Smith, G.A.; Ritchie, S.A. Expectoration of flaviviruses during sugar feeding by mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 845–850. [Google Scholar] [CrossRef]
- Moore, J.S.; Kelly, T.B.; Killick-Kendrick, R.; Killick-Kendrick, M.; Wallbanks, K.R.; Molyneux, D.H. Honeydew sugars in wild-caught Phlebotomus ariasi detected by high performance liquid chromatography (HPLC) and gas chromatography (GC). Med. Vet. Entomol. 1987, 1, 427–434. [Google Scholar] [CrossRef]
- Molyneux, D.H.; Moore, J.; Maroli, M. Sugars in sandflies. Parassitologia 1991, 33, 431–436. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | D4 | D5 | D6 | D7 | D10 | D11 | D12 | D13 | D14 | D17 | D18 | D19 | D20 | D21 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Females 1 | + | + | + | + | − | − | ||||||||
| Females 2 | + | + | + | + | − | + | ||||||||
| Females 3 | + | + | + | + | + | + | − | + | + | − | − | + | + | − |
| Males 1 | + | + | + | + | − | + | ||||||||
| Males 2 | + | + | + | + | − | + | ||||||||
| Males 3 | + | + | + | + | + | + | − | + | − | − | − | + | − | + |
| Cage B | D7 | D8 | D9 | D12 | D13 | D14 | D15 | D16 | D21 |
|---|---|---|---|---|---|---|---|---|---|
| F cage: RNA copies in sugar drop | 440 | neg | 192 | 199 | neg | 3 | |||
| Infected F/tested F | 7/30 (23%) | 5/30 (17%) | 7/200 (3.5%) | ||||||
| M cage: RNA copies in sugar drop | 1919 | neg | 2258 | 142 | 1 | neg | |||
| Infected M/tested M | 9/30 (30%) | 2/30 (7%) | 0/92 (0%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jancarova, M.; Bichaud, L.; Hlavacova, J.; Priet, S.; Ayhan, N.; Spitzova, T.; Volf, P.; Charrel, R.N. Experimental Infection of Sand Flies by Massilia Virus and Viral Transmission by Co-Feeding on Sugar Meal. Viruses 2019, 11, 332. https://doi.org/10.3390/v11040332
Jancarova M, Bichaud L, Hlavacova J, Priet S, Ayhan N, Spitzova T, Volf P, Charrel RN. Experimental Infection of Sand Flies by Massilia Virus and Viral Transmission by Co-Feeding on Sugar Meal. Viruses. 2019; 11(4):332. https://doi.org/10.3390/v11040332
Chicago/Turabian StyleJancarova, Magdalena, Laurence Bichaud, Jana Hlavacova, Stephane Priet, Nazli Ayhan, Tatiana Spitzova, Petr Volf, and Remi N. Charrel. 2019. "Experimental Infection of Sand Flies by Massilia Virus and Viral Transmission by Co-Feeding on Sugar Meal" Viruses 11, no. 4: 332. https://doi.org/10.3390/v11040332
APA StyleJancarova, M., Bichaud, L., Hlavacova, J., Priet, S., Ayhan, N., Spitzova, T., Volf, P., & Charrel, R. N. (2019). Experimental Infection of Sand Flies by Massilia Virus and Viral Transmission by Co-Feeding on Sugar Meal. Viruses, 11(4), 332. https://doi.org/10.3390/v11040332