Histo-Blood Group Antigens in Children with Symptomatic Rotavirus Infection


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Specimens
2.2. Rotavirus Detection and Genotyping
2.3. Determination of Histo-Blood Group Antigens in Saliva
2.4. Genotypic Characterization of the FUT2 Gene (Secretor Status)
2.5. Statistical Analysis
3. Results
3.1. Study Population and Sample Collection
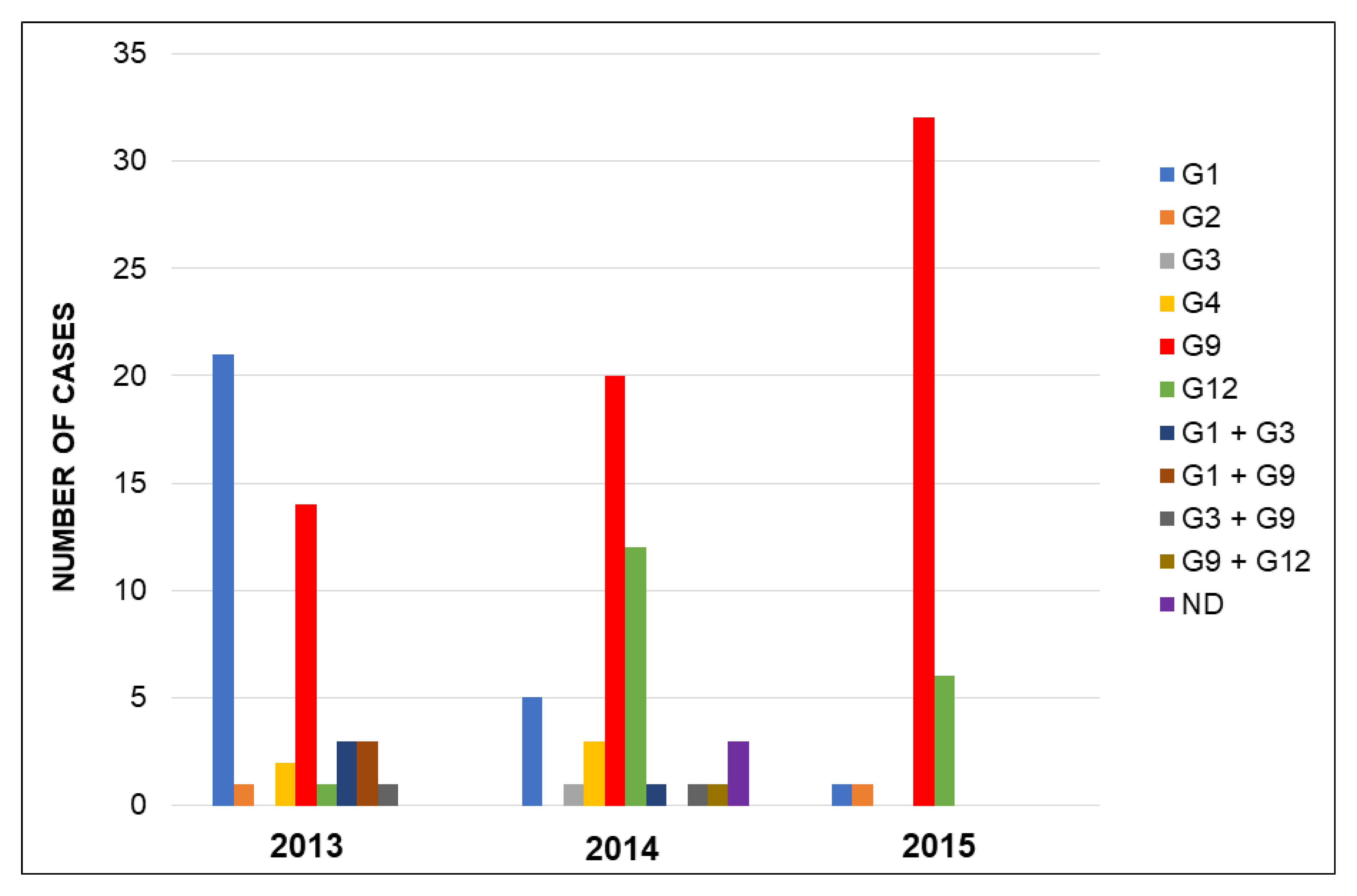
3.2. Rotavirus Genotypes
3.3. Secretor (FUT2) Status
3.4. Lewis and ABO Phenotypes
3.5. Association between HBGAs and Rotavirus Genotypes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Steele, A.D.; Duque, J.; Parashar, U.D. WHO-coordinated Global Rotavirus Surveillance Network. 2008 estimate of worldwide rotavirus-associated mortality in children younger than 5 years before the introduction of universal rotavirus vaccination programmes: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 136–141. [Google Scholar] [CrossRef]
- WHO. Meeting of the Strategic Advisory Group of Experts on immunization, October 2009--Conclusions and recommendations. Biologicals. 2010, 38, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Armah, G.E.; Sow, S.O.; Breiman, R.F.; Dallas, M.J.; Tapia, M.D.; Feikin, D.R.; Binka, F.N.; Steele, A.D.; Laserson, K.F.; Anasah, N.A.; et al. Efficacy of pentavalent rotavirus vaccine against severe rotavirus gastroenteritis in infants in developing countries in sub-Saharan Africa: A randomised, double-blind, placebo-controlled trial. Lancet 2010, 376, 606–614. [Google Scholar] [PubMed]
- Madhi, S.A.; Cunliffe, N.A.; Steele, D.; Witte, D.; Kirsten, M.; Louw, C.; Ngwira, B.; Victor, J.C.; Gillard, P.H.; Cheuvart, B.B.; et al. Effect of human rotavirus vaccine on severe diarrhea in African infants. N. Engl. J. Med. 2010, 362, 289–298. [Google Scholar] [PubMed]
- Harris, V.C.; Armah, G.; Fuentes, S.; Korpela, K.E.; Parashar, U.; Victor, J.C.; Tate, J.; de Weerth, C.; Giaquinto, C.; Wiersinga, W.J.; et al. Significant correlation between the infant gut microbiome and rotavirus vaccine response in rural Ghana. J. Infect. Dis. 2017, 215, 34–41. [Google Scholar] [CrossRef]
- Harris, V.C.; Haak, B.W.; Handley, S.A.; Jiang, B.; Velasquez, D.E.; Hykes, B.L., Jr.; Droit, L.; Berbers, G.A.N.; Kemper, E.M.; van Leeuwen, E.M.M.; et al. Effect of antibiotic-mediated microbiome modulation on rotavirus vaccine immunogenicity: A human, randomized-control proof-of-concept trial. Cell Host Microbe 2018, 24, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Xia, M.; Tan, M.; Zhong, W.; Wei, C.; Wang, L.; Morrow, A.; Jiang, X. Spike protein VP8* of human rotavirus recognizes histo-blood group antigens in a type-specific manner. J. Virol. 2012, 86, 4833–4843. [Google Scholar] [CrossRef]
- Nordgren, J.; Sharma, S.; Bucardo, F.; Nasir, W.; Günaydin, G.; Ouermi, D.; Nitiema, L.W.; Becker-Dreps, S.; Simpore, J.; Hammarstrom, L.; et al. Both lewis and secretor status mediate susceptibility to rotavirus infections in a rotavirus genotype-dependent manner. Clin. Infect. Dis. 2014, 59, 1567–1573. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, Y.; Tan, M. Histo-blood group antigens as receptors for rotavirus, new understanding on rotavirus epidemiology and vaccine strategy. Emerg. Microbes Infect. 2017, 6, e22. [Google Scholar] [CrossRef] [PubMed]
- Henry, S.; Jovall, P.Å.; Ghardashkhani, S.; Elmgren, A.; Martinsson, T.; Larson, G.; Samuelsson, B. Structural and immunochemical identification of Le(a), Le(b), H type 1, and related glycolipids in small intestinal mucosa of a group O Le(a-b-) nonsecretor. Glycoconj. J. 1997, 14, 209–223. [Google Scholar] [CrossRef]
- Marionneau, S.; Cailleau-Thomas, A.; Rocher, J.; Le Moullac-Vaidye, B.; Ruvoën, N.; Clément, M.; Le Pendu, J. ABH and Lewis histo-blood group antigens, a model for the meaning of oligosaccharide diversity in the face of a changing world. Biochimie 2001, 83, 565–573. [Google Scholar]
- Le Pendu, J. Histo-blood group antigen and human milk oligosaccharides: Genetic polymorphism and risk of infectious diseases. Adv. Exp. Med. Biol. 2004, 554, 135–143. [Google Scholar]
- Böhm, R.; Fleming, F.E.; Maggioni, A.; Dang, V.T.; Holloway, G.; Coulson, B.S.; von Itzstein, M.; Haselhorst, T. Revisiting the role of histo-blood group antigens in rotavirus host-cell invasion. Nat. Commun. 2015, 6, 5907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, D.C.; Currier, R.L.; Staat, M.A.; Sahni, L.C.; Selvarangan, R.; Halasa, N.B.; Englund, J.A.; Weinberg, G.A.; Boom, J.A.; Szilagyi, P.G.; et al. Epidemiologic association between FUT2 secretor status and severe rotavirus gastroenteritis in children in the United States. JAMA Pediatr. 2015, 169, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-F.; Long, Y.; Tan, M.; Zhang, T.; Huang, Q.; Jiang, X.; Tan, W.-F.; Li, J.-D.; Hu, G.-F.; Tang, S.; et al. P[8] and P[4] Rotavirus infection associated with secretor phenotypes among children in South China. Sci. Rep. 2016, 6, 34591. [Google Scholar] [PubMed]
- Lindesmith, L.; Moe, C.; Marionneau, S.; Ruvoen, N.; Jiang, X.; Lindblad, L.; Stewart, P.; LePendu, J.; Baric, R. Human susceptibility and resistance to Norwalk virus infection. Nat. Med. 2003, 9, 548–553. [Google Scholar] [PubMed]
- Hutson, A.M.; Atmar, R.L.; Graham, D.Y.; Estes, M.K. Norwalk virus infection and disease is associated with ABO histo-blood group type. J. Infect. Dis. 2002, 185, 1335–1337. [Google Scholar] [CrossRef]
- Todd, S.; Page, N.A.A.; Duncan Steele, A.; Peenze, I.; Cunliffe, N.A. Rotavirus Strain Types Circulating in Africa: Review of Studies Published during 1997–2006. J. Infect. Dis. 2010, 202, S34–S42. [Google Scholar] [CrossRef]
- Nordgren, J.; Nitiema, L.W.; Sharma, S.; Ouermi, D.; Traore, A.S.; Simpore, J.; Svensson, L. Emergence of unusual G6P[6] rotaviruses in children, Burkina Faso, 2009–2010. Emerg. Infect Dis. 2012, 18, 589–597. [Google Scholar] [CrossRef]
- Bucardo, F.; Nordgren, J.; Reyes, Y.; Gonzalez, F.; Sharma, S.; Svensson, L. The Lewis A phenotype is a restriction factor for Rotateq and Rotarix vaccine-take in Nicaraguan children. Sci. Rep. 2018, 8, 1502. [Google Scholar] [CrossRef] [Green Version]
- Ayouni, S.; Sdiri-Loulizi, K.; de Rougemont, A.; Estienney, M.; Ambert-Balay, K.; Aho, S.; Hamami, S.; Aouni, M.; Neji-Guediche, M.; Pothier, P.; et al. Rotavirus P[8] Infections in Persons with Secretor and Nonsecretor Phenotypes, Tunisia. Emerg. Infect. Dis. 2015, 21, 2055–2058. [Google Scholar] [CrossRef] [Green Version]
- Iturriza-Gómara, M.; Dallman, T.; Bányai, K.; Böttiger, B.; Buesa, J.; Diedrich, S.; Fiore, L.; Johansen, K.; Koopmans, M.; Korsun, N.; et al. Rotavirus genotypes co-circulating in Europe between 2006 and 2009 as determined by EuroRotaNet, a pan-European collaborative strain surveillance network. Epidemiol. Infect. 2011, 139, 895–909. [Google Scholar] [CrossRef]
- Nordgren, J.; Nitiema, L.W.; Ouermi, D.; Simpore, J.; Svensson, L. Host genetic factors affect susceptibility to norovirus infections in Burkina Faso. PLoS ONE 2013, 8, e69557. [Google Scholar] [CrossRef]
- Serpa, J.; Mendes, N.; Reis, C.A.; Santos Silva, L.F.; Almeida, R.; Le Pendu, J.; David, L. Two new FUT2 (fucosyltransferase 2 gene) missense polymorphisms, 739G→A and 839T→C, are partly responsible for non-secretor status in a Caucasian population from Northern Portugal. Biochem. J. 2004, 383, 469–474. [Google Scholar] [CrossRef]
- Marionneau, S.; Ruvoën, N.; Le Moullac-Vaidye, B.; Clement, M.; Cailleau-Thomas, A.; Ruiz-Palacios, G.; Huang, P.; Jiang, X.; Le Pendu, J. Norwalk Virus binds to histo-blood group antigens present on gastroduodenal epithelial cells of secretor individuals. Gastroenterology 2002, 122, 1967–1977. [Google Scholar]
- Carlsson, B.; Kindberg, E.; Buesa, J.; Rydell, G.E.; Lidón, M.F.; Montava, R.; Abu Mallouh, R.; Grahn, A.; Rodriguez-Diaz, J.; Bellido, J.; et al. The G428A nonsense mutation in FUT2 provides strong but not absolute protection against symptomatic GII.4 norovirus infection. PLoS ONE. 2009, 4, e5593. [Google Scholar]
- Tan, M.; Jiang, X. Histo-blood group antigens: A common niche for norovirus and rotavirus. Expert Rev. Mol. Med. 2014, 16, e5. [Google Scholar] [CrossRef]
- Liu, Y.; Ramelot, T.A.; Huang, P.; Liu, Y.; Li, Z.; Feizi, T.; Zhong, W.; Wu, F.T.; Tan, M.; Kennedy, M.A.; et al. Glycan specificity of P[19] rotavirus and comparison with those of related P genotypes. J. Virol. 2016, 90, 9983–9996. [Google Scholar] [CrossRef]
- Hu, L.; Sankaran, B.; Laucirica, D.R.; Patil, K.; Salmen, W.; Ferreon, A.C.M.; Tsoi, P.S.; Lasanajak, Y.; Smith, D.F.; Ramani, S.; et al. Glycan recognition in globally dominant human rotaviruses. Nat. Commun. 2018, 9, 2631. [Google Scholar]
- Hu, L.; Crawford, S.E.; Czako, R.; Cortes-Penfield, N.W.; Smith, D.F.; Le Pendu, J.; Estes, M.K.; Prasad, B.V. Cell attachment protein VP8* of a human rotavirus specifically interacts with A-type histo-blood group antigen. Nature 2012, 485, 256–259. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, P.; Jiang, B.; Tan, M.; Morrow, A.L.; Jiang, X. Poly-LacNAc as an age-specific ligand for rotavirus P[11] in neonates and infants. PLoS ONE 2013, 8, e78113. [Google Scholar] [CrossRef]
- Santos, N.; Hoshino, Y. Global distribution of rotavirus serotypes/genotypes and its implication for the development and implementation of an effective rotavirus vaccine. Rev. Med. Virol. 2005, 15, 29–56. [Google Scholar]
- Das, S.; Varghese, V.; Chaudhuri, S.; Barman, P.; Kojima, K.; Dutta, P.; Bhattacharya, S.K.; Krishnan, T.; Kobayashi, N.; Naik, T.N. Genetic variability of human rotavirus strains isolated from eastern and northern India. J. Med. Virol. 2004, 72, 156–161. [Google Scholar] [CrossRef]
- Shim, J.O.; Son, D.W.; Shim, S.-Y.; Ryoo, E.; Kim, W.; Jung, Y.-C. Clinical characteristics and genotypes of rotaviruses in a neonatal intensive care unit. Pediatr. Neonatol. 2012, 53, 18–23. [Google Scholar]
- Waggie, Z.; Hawkridge, A.; Hussey, G.D. Review of rotavirus studies in Africa: 1976–2006. J. Infect. Dis. 2010, 202, S23–S33. [Google Scholar]
- Van Trang, N.; Vu, H.T.; Le, N.T.; Huang, P.; Jiang, X.; Anh, D.D. Association between norovirus and rotavirus infection and histo-blood group antigen types in Vietnamese children. J. Clin. Microbiol. 2014, 52, 1366–1374. [Google Scholar]
- Ameno, S.; Kimura, H.; Ameno, K.; Zhang, X.; Kinoshita, H.; Kubota, T.; Ijiri, I. Lewis and Secretor gene effects on Lewis antigen and postnatal development of Lewis blood type. Biol. Neonatol. 2001, 79, 91–96. [Google Scholar] [CrossRef]
- Imbert-Marcille, B.M.; Barbé, L.; Dupé, M.; Le Moullac-Vaidye, B.; Besse, B.; Peltier, C.; Ruvoen-Clouet, N.; Le Pendu, J. A FUT2 gene common polymorphism determines resistance to rotavirus a of the P[8] genotype. J. Infect. Dis. 2014, 209, 1227–1230. [Google Scholar]
- Rodríguez-Díaz, J.; García-Mantrana, I.; Vila-Vicent, S.; Gozalbo-Rovira, R.; Buesa, J.; Monedero, V.; Collado, M.C. Relevance of secretor status genotype and microbiota composition in susceptibility to rotavirus and norovirus infections in humans. Sci. Rep. 2017, 7, 45559. [Google Scholar] [CrossRef]
- Pérez-Ortín, R.; Santiso-Bellón, C.; Vila-Vicent, S.; Carmona-Vicente, N.; Rodríguez-Díaz, J.; Buesa, J. Rotavirus symptomatic infection among vaccinated children in Valencia, Spain. BMC Infect. Dis. submitted.
- Barbé, L.; Le Moullac-Vaidye, B.; Echasserieau, K.; Bernardeau, K.; Carton, T.; Bovin, N.; Nordgren, J.; Svensson, L.; Ruvoen-Clouet, N.; Le Pendu, J. Histo-blood group antigen-binding specificities of human rotaviruses are associated with gastroenteritis but not with in vitro infection. Sci. Rep. 2018, 8, 12961. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Patients | Controls a | p Value b | Odds Ratio c | Donors | p Value b | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| (n = 133) (%) | (n = 50) (%) | 95% CI | (n = 283,399) (%) | |||||||
| Blood Group | ||||||||||
| O | 50 | (37.6) | 15 | (36.6) | 0.365 | * | 146,454 | (51.7) | 0.003 | |
| A | 64 | (48.1) | 17 | (41.5) | 0.89 | (0.4–2) | 110,273 | (38.9) | ||
| B | 10 | (7.5) | 7 | (17.1) | 2.31 | (0.7–7.2) | 19,054 | (6.7) | ||
| AB | 9 | (6.8) | 2 | (4.9) | 0.78 | (0.1 - 3.6) | 7618 | (2.7) | ||
| Secretor (FUT2) | ||||||||||
| Secretor | 131 | (98.5) | 35 | (70) | 0.000 | * | NA | - | ||
| Non-secretor | 2 | (1.5) | 15 | (30) | 25 | (6.7–100) | NA | |||
| Lewis (FUT3) | ||||||||||
| Negative | 8 | (6) | 3 | (6) | 1.000 | * | 31,090 | (11) | 0.091 | |
| Positive | 125 | (94) | 47 | (94) | 0.97 | (0.26–4.84) | 252,309 | (89) | ||
| Lewis A | ||||||||||
| Negative | 73 | (54.9) | 15 | (30) | 0.005 | * | 225,219 | (79.5) | 0.000 | |
| Positive | 60 | (45.1) | 35 | (70) | 2.81 | (1.42–5.78) | 56,180 | (20.5) | ||
| Lewis B | ||||||||||
| Negative | 10 | (7.5) | 18 | (36) | 0.000 | * | 88,784 | (31.3) | 0.000 | |
| Positive | 123 | (92.5) | 32 | (64) | 0.15 | (0.06–0.35) | 194,615 | (68.7) | ||
| Lewis A/B | ||||||||||
| Le a– b– | 8 | (6) | 3 | (6) | 0.000 | * | 31,090 | (11) | 0.000 | |
| Le a– b+ | 65 | (48.9) | 12 | (24) | 0.49 | (0.1–2.6) | 194,129 | (68.5) | ||
| Le a+ b– | 2 | (1.5) | 15 | (30) | 16.47 | (2.6–169.7) | 57,694 | (20.3) | ||
| Le a+ b+ | 58 | (43.6) | 20 | (40) | 0.9 | (0.2–4.6) | 486 | (0.2) | ||
| G1P[8] | G2P[4] | G3P[8] | G4P[8] | G9P[8] | G12P[8] | p Value a | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (n = 27) (%) | (n = 2) (%) | (n = 1) (%) | (n = 5) (%) | (n = 66) (%) | (n = 19) (%) | ||||||||
| Blood Group | |||||||||||||
| O | 10 | (37) | 0 | (0) | 0 | (0) | 2 | (40) | 24 | (36.4) | 9 | (47.4) | 0.826 |
| A | 14 | (51.8) | 2 | (100) | 1 | (100) | 3 | (60) | 30 | (45.4) | 7 | (36.8) | |
| B | 2 | (7.4) | 0 | (0) | 0 | (0) | 0 | (0) | 7 | (10.6) | 1 | (5.3) | |
| AB | 1 | (3.7) | 0 | (0) | 0 | (0) | 0 | (0) | 5 | (7.6) | 2 | (10.5) | |
| Secretor | |||||||||||||
| Heterozygous | 16 | (59.3) | 2 | (100) | 1 | (100) | 3 | (60) | 44 | (66.7) | 8 | (42.1) | 0.265 |
| Homozygous | 10 | (37) | 0 | (0) | 0 | (0) | 2 | (40) | 22 | (33.3) | 11 | (57.9) | |
| Non-secretor | 1 | (3.7) | 0 | (0) | 0 | (0) | 0 | (0) | 0 | (0) | 0 | (0) | |
| Lewis A / B | |||||||||||||
| Le a– b– | 4 | (50) | 0 | (0) | 0 | (0) | 0 | (0) | 2 | (25) | 1 | (12.5) | 0.146 |
| Le a– b+ | 15 | (23.1) | 1 | (1.5) | 0 | (0) | 3 | (4.6) | 30 | (46.1) | 11 | (16.9) | |
| Le a+ b– | 1 | (50) | 0 | (0) | 0 | (0) | 0 | (0) | 0 | (0) | 0 | (0) | |
| Le a+ b+ | 7 | (12.1) | 1 | (1.7) | 1 | (1.7) | 2 | (3.4) | 34 | (58.6) | 7 | (12.1) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Ortín, R.; Vila-Vicent, S.; Carmona-Vicente, N.; Santiso-Bellón, C.; Rodríguez-Díaz, J.; Buesa, J. Histo-Blood Group Antigens in Children with Symptomatic Rotavirus Infection. Viruses 2019, 11, 339. https://doi.org/10.3390/v11040339
Pérez-Ortín R, Vila-Vicent S, Carmona-Vicente N, Santiso-Bellón C, Rodríguez-Díaz J, Buesa J. Histo-Blood Group Antigens in Children with Symptomatic Rotavirus Infection. Viruses. 2019; 11(4):339. https://doi.org/10.3390/v11040339
Chicago/Turabian StylePérez-Ortín, Raúl, Susana Vila-Vicent, Noelia Carmona-Vicente, Cristina Santiso-Bellón, Jesús Rodríguez-Díaz, and Javier Buesa. 2019. "Histo-Blood Group Antigens in Children with Symptomatic Rotavirus Infection" Viruses 11, no. 4: 339. https://doi.org/10.3390/v11040339
APA StylePérez-Ortín, R., Vila-Vicent, S., Carmona-Vicente, N., Santiso-Bellón, C., Rodríguez-Díaz, J., & Buesa, J. (2019). Histo-Blood Group Antigens in Children with Symptomatic Rotavirus Infection. Viruses, 11(4), 339. https://doi.org/10.3390/v11040339