First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Screening
2.2. Virus Cultivation
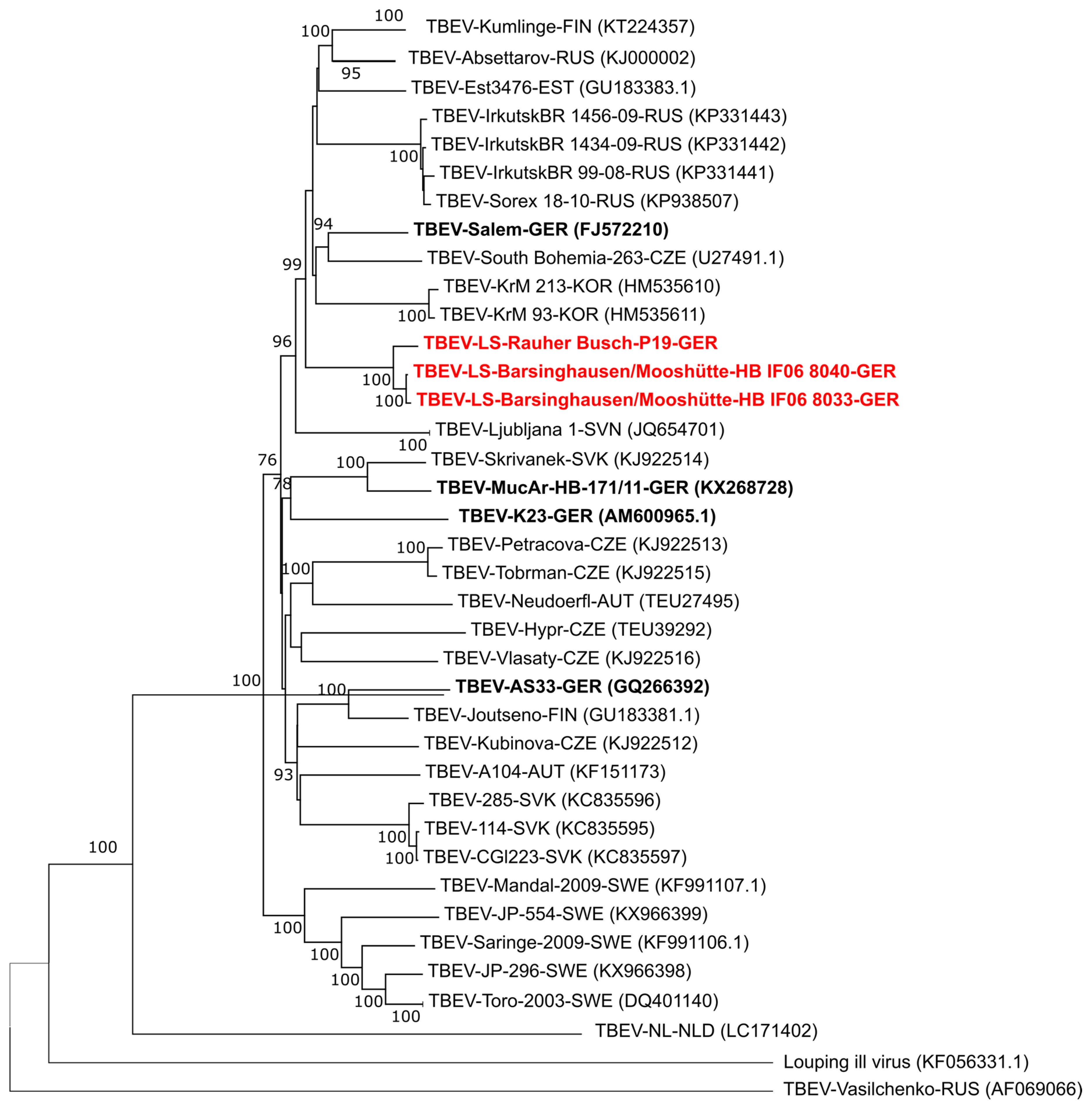
2.3. PCR and Phylogenetic Analysis of TBEV Isolates
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gritsun, T.S.; Lashkevich, V.A.; Gould, E.A. Tick-borne encephalitis. Antivir. Res. 2003, 57, 129–146. [Google Scholar] [CrossRef]
- Lindquist, L.; Vapalahti, O. Tick-borne encephalitis. Lancet 2008, 371, 1861–1871. [Google Scholar] [CrossRef]
- Hayasaka, D.; Nagata, N.; Fujii, Y.; Hasegawa, H.; Sata, T.; Suzuki, R.; Gould, E.A.; Takashima, I.; Koike, S. Mortality following peripheral infection with Tick-borne encephalitis virus results from a combination of central nervous system pathology, systemic inflammatory and stress responses. Virology 2009, 390, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.X. Tick-borne encephalitis virus: Advances in molecular biology and vaccination strategy in the next century. Zent. Bakteriol. 1999, 289, 506–510. [Google Scholar] [CrossRef]
- Kupča, A.M.; Essbauer, S.; Zoeller, G.; de Mendonça, P.G.; Brey, R.; Rinder, M.; Pfister, K.; Spiegel, M.; Doerrbecker, B.; Pfeffer, M.; et al. Isolation and molecular characterization of a tick-borne encephalitis virus strain from a new tick-borne encephalitis focus with severe cases in Bavaria, Germany. Ticks Tick-Borne Dis. 2010, 1, 44–51. [Google Scholar] [CrossRef]
- Dai, X.; Shang, G.; Lu, S.; Yang, J.; Xu, J. A new subtype of eastern tick-borne encephalitis virus discovered in Qinghai-Tibet Plateau, China. Emerg. Microbes Infect. 2018, 7, 74. [Google Scholar] [CrossRef]
- Kovalev, S.Y.; Mukhacheva, T.A. Reconsidering the classification of tick-borne encephalitis virus within the Siberian subtype gives new insights into its evolutionary history. Infect. Genet. Evol. 2017, 55, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Suss, J. Tick-borne encephalitis in Europe and beyond--the epidemiological situation as of 2007. Euro Surveill. 2008, 13, 717–727. [Google Scholar]
- Charrel, R.N.; Attoui, H.; Butenko, A.M.; Clegg, J.C.; Deubel, V.; Frolova, T.V.; Gould, E.A.; Gritsun, T.S.; Heinz, F.X.; Labuda, M.; et al. Tick-borne virus diseases of human interest in Europe. Clin. Microbiol. Infect. 2004, 10, 1040–1055. [Google Scholar] [CrossRef] [Green Version]
- Mandl, C.W.; Heinz, F.X.; Stöckl, E.; Kunz, C. Genome sequence of tick-borne encephalitis virus (Western subtype) and comparative analysis of nonstructural proteins with other flaviviruses. Virology 1989, 173, 291–301. [Google Scholar] [CrossRef]
- Heinz, F.X. Epitope mapping of flavivirus glycoproteins. Adv. Virus Res. 1986, 31, 103–168. [Google Scholar] [PubMed]
- Epidemiologisches Bulletin 17/2018. In Robert Koch Institut. 2018, Vol. 17. Available online: https://www.rki.de/DE/Content/Infekt/EpidBull/Archiv/2018/Ausgaben/17_18.html (accessed on 15 March 2019).
- Dobler, G.; Gniel, D.; Petermann, R.; Pfeffer, M. Epidemiology and distribution of tick-borne encephalitis. Wien. Med. Wochenschr. 2012, 162, 230–238. [Google Scholar] [CrossRef]
- SurvStat@RKI 2.0. In Robert Koch Institut. 2018. Available online: https://survstat.rki.de/Content/Instruction/Main.aspx (accessed on 15 March 2019).
- Schwaiger, M.; Cassinotti, P. Development of a quantitative real-time RT-PCR assay with internal control for the laboratory detection of tick borne encephalitis virus (TBEV) RNA. J. Clin. Virol. 2003, 27, 136–145. [Google Scholar] [CrossRef]
- Andersen, N.S.; Bestehorn, M.; Chitimia-Dobler, L.; Kolmos, H.J.; Jensen, P.M.; Dobler, G.; Skarphédinsson, S. Phylogenetic characterization of tick-borne encephalitis virus from Bornholm, Denmark. Ticks Tick-Borne Dis. 2018, 10, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling Genomes and Mini-metagenomes from Highly Chimeric Reads, Berlin, Heidelberg, 2013; Springer: Berlin, Heidelberg, 2013; pp. 158–170. [Google Scholar]
- Pettersson, J.H.; Golovljova, I.; Vene, S.; Jaenson, T.G. Prevalence of tick-borne encephalitis virus in Ixodes ricinus ticks in northern Europe with particular reference to Southern Sweden. Parasites Vectors 2014, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Katargina, O.; Russakova, S.; Geller, J.; Kondrusik, M.; Zajkowska, J.; Zygutiene, M.; Bormane, A.; Trofimova, J.; Golovljova, I. Detection and characterization of tick-borne encephalitis virus in Baltic countries and eastern Poland. Plos ONE 2013, 8, e61374. [Google Scholar] [CrossRef]
- Süss, J.; Schrader, C.; Falk, U.; Wohanka, N. Tick-borne encephalitis (TBE) in Germany — Epidemiological data, development of risk areas and virus prevalence in field-collected ticks and in ticks removed from humans. Int. J. Med. Microbiol. Suppl. 2004, 293, 69–79. [Google Scholar] [CrossRef]
- Suss, J.; Schrader, C.; Abel, U.; Voigt, W.P.; Schosser, R. Annual and seasonal variation of tick-borne encephalitis virus (TBEV) prevalence in ticks in selected hot spot areas in Germany using a nRT-PCR: results from 1997 and 1998. Zent. Bakteriol. 1999, 289, 564–578. [Google Scholar] [CrossRef]
- Jovani, R.; Tella, J.L. Parasite prevalence and sample size: Misconceptions and solutions. Trends Parasitol. 2006, 22, 214–218. [Google Scholar] [CrossRef]
- Kovalev, S.Y.; Mukhacheva, T.A. Tick-borne encephalitis virus subtypes emerged through rapid vector switches rather than gradual evolution. Ecol. Evol. 2014, 4, 4307–4316. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, M.; Růžek, D.; Křivanec, K.; Zöller, G.; Essbauer, S.; Pfeffer, M.; Zanotto, P.M.d.A.; Hufert, F.T.; Dobler, G. Relation of genetic phylogeny and geographical distance of tick-borne encephalitis virus in central Europe. J. Gen. Virol. 2011, 92, 1906–1916. [Google Scholar] [CrossRef]
- Chitimia-Dobler, L.; Lemhöfer, G.; Król, N.; Bestehorn, M.; Dobler, G.; Pfeffer, M. Repeated isolation of tick-borne encephalitis virus from adult Dermacentor reticulatus ticks in an endemic area in Germany. Parasites Vectors 2019, 12, 90. [Google Scholar] [CrossRef]
- Zanotto, P.M.; Gao, G.F.; Gritsun, T.; Marin, M.S.; Jiang, W.R.; Venugopal, K.; Reid, H.W.; Gould, E.A. An arbovirus cline across the northern hemisphere. Virology 1995, 210, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Heinze, D.M.; Gould, E.A.; Forrester, N.L. Revisiting the clinal concept of evolution and dispersal for the tick-borne flaviviruses by using phylogenetic and biogeographic analyses. J. Virol. 2012, 86, 8663–8671. [Google Scholar] [CrossRef]
- Weidmann, M.; Frey, S.; Freire, C.C.; Essbauer, S.; Ruzek, D.; Klempa, B.; Zubrikova, D.; Vogerl, M.; Pfeffer, M.; Hufert, F.T.; et al. Molecular phylogeography of tick-borne encephalitis virus in central Europe. J. Gen. Virol. 2013, 94 (Pt 9), 2129–2139. [Google Scholar] [CrossRef]
- Kriz, B.; Daniel, M.; Benes, C.; Maly, M. The role of game (wild boar and roe deer) in the spread of tick-borne encephalitis in the Czech Republic. Vector Borne Zoonotic Dis. (Larchmt. N.Y.) 2014, 14, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Balling, A.; Plessow, U.; Beer, M.; Pfeffer, M. Prevalence of antibodies against tick-borne encephalitis virus in wild game from Saxony, Germany. Ticks Tick-Borne Dis. 2014, 5, 805–809. [Google Scholar] [CrossRef]
- Waldenstrom, J.; Lundkvist, A.; Falk, K.I.; Garpmo, U.; Bergstrom, S.; Lindegren, G.; Sjostedt, A.; Mejlon, H.; Fransson, T.; Haemig, P.D.; et al. Migrating birds and tickborne encephalitis virus. Emerg. Infect. Dis. 2007, 13, 1215–1218. [Google Scholar] [CrossRef]
- Bestehorn, M.; Weigold, S.; Kern, W.V.; Chitimia-Dobler, L.; Mackenstedt, U.; Dobler, G.; Borde, J.P. Phylogenetics of tick-borne encephalitis virus in endemic foci in the upper Rhine region in France and Germany. PLoS ONE 2018, 13, e0204790. [Google Scholar] [CrossRef] [PubMed]
- Hütter, A. Güterverkehr in Deutschland 2014. In Statistisches Bundesamt Wista; 2016; Vol. WISTA 1/2016, pp. 47–62. Available online: https://www.destatis.de/DE/Publikationen/WirtschaftStatistik/2016/01/Wista_1_2016.pdf?__blob=publicationFile (accessed on 15 March 2019).
- Kovalev, S.Y.; Mukhacheva, T.A. Clusterons as a tool for monitoring populations of tick-borne encephalitis virus. J. Med. Virol. 2014, 86, 283–289. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boelke, M.; Bestehorn, M.; Marchwald, B.; Kubinski, M.; Liebig, K.; Glanz, J.; Schulz, C.; Dobler, G.; Monazahian, M.; Becker, S.C. First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany. Viruses 2019, 11, 462. https://doi.org/10.3390/v11050462
Boelke M, Bestehorn M, Marchwald B, Kubinski M, Liebig K, Glanz J, Schulz C, Dobler G, Monazahian M, Becker SC. First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany. Viruses. 2019; 11(5):462. https://doi.org/10.3390/v11050462
Chicago/Turabian StyleBoelke, Mathias, Malena Bestehorn, Birgit Marchwald, Mareike Kubinski, Katrin Liebig, Julien Glanz, Claudia Schulz, Gerhard Dobler, Masyar Monazahian, and Stefanie C. Becker. 2019. "First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany" Viruses 11, no. 5: 462. https://doi.org/10.3390/v11050462
APA StyleBoelke, M., Bestehorn, M., Marchwald, B., Kubinski, M., Liebig, K., Glanz, J., Schulz, C., Dobler, G., Monazahian, M., & Becker, S. C. (2019). First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany. Viruses, 11(5), 462. https://doi.org/10.3390/v11050462