Genistein Has Antiviral Activity against Herpes B Virus and Acts Synergistically with Antiviral Treatments to Reduce Effective Dose

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Media, and Virus
2.2. Chemicals and Reagents
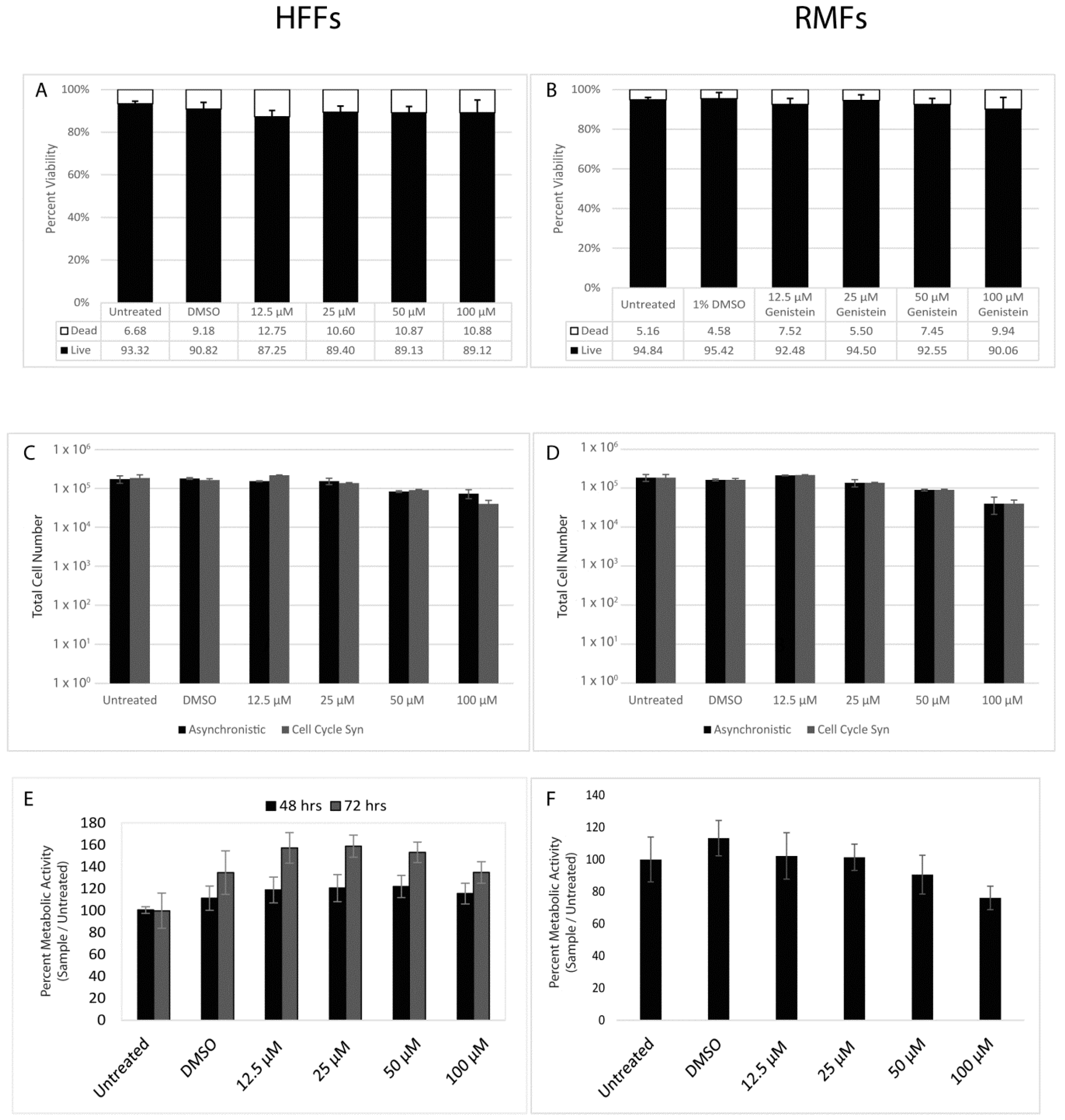
2.3. Trypan Blue Dye Exclusion Assay
2.4. Cell Proliferation Assay
2.5. Bromodeoxyuridine (BrdU) Incorporation Assay
2.6. DNA Isolation and RT-PCR
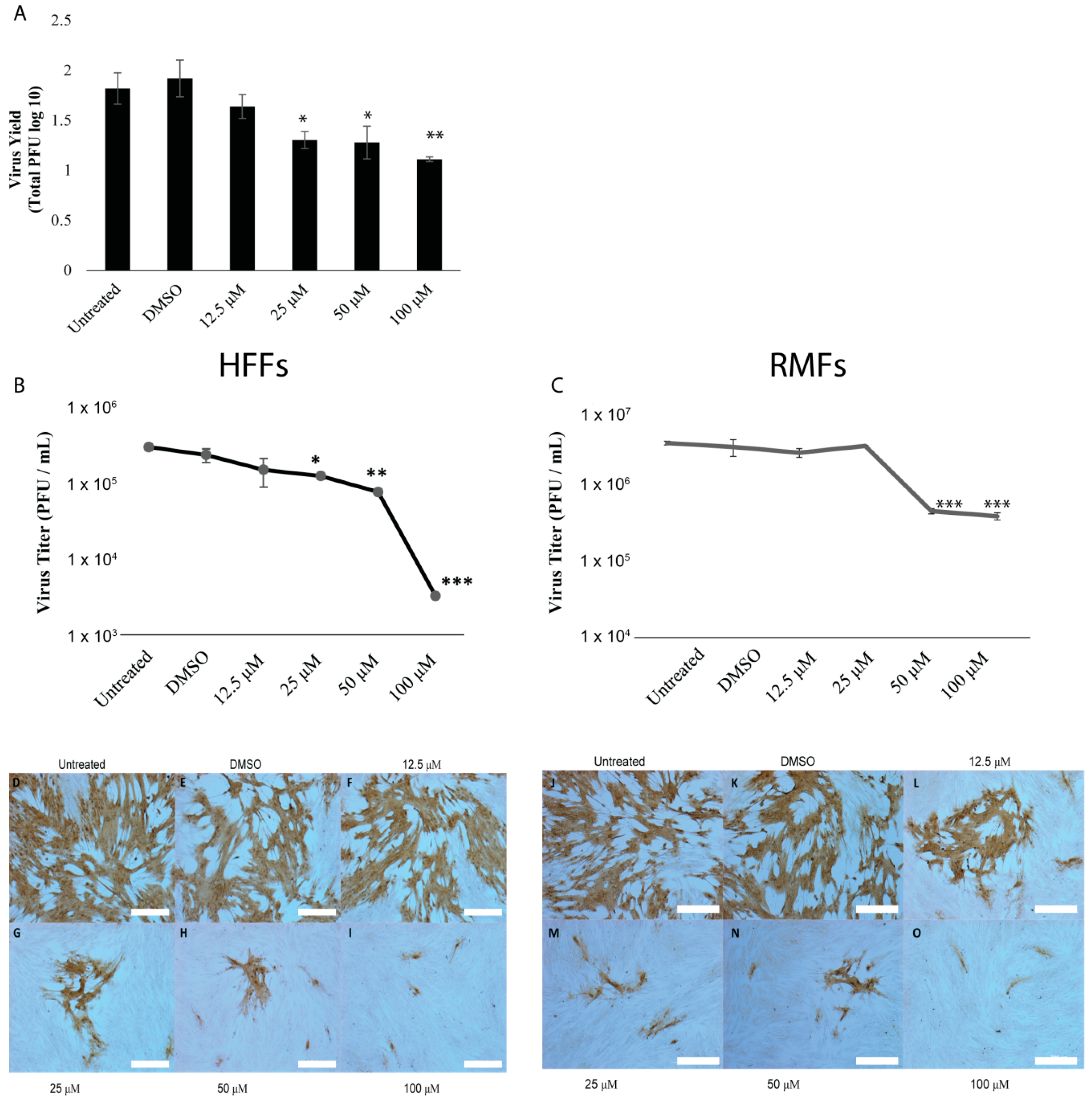
2.7. Indirect Virus Yield Assay
2.8. Plaque Reduction Assay
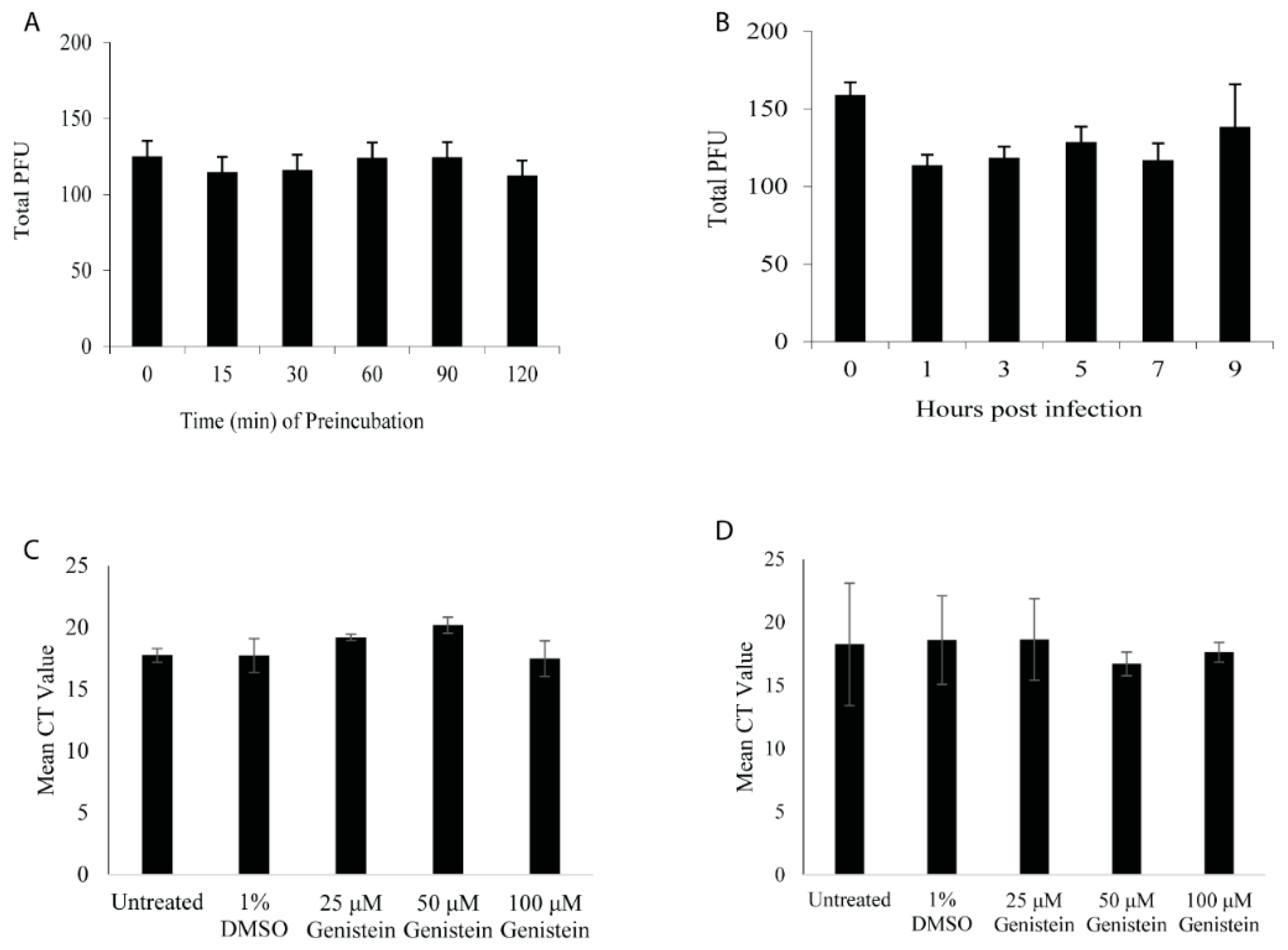
2.9. Direct Inactivation Using Modified Plaque Reduction Assay
2.10. Virus Yield Assay
2.11. Cell-Based ELISA Assay
2.12. Statistical Analysis
3. Results
3.1. Genistein Does Not Exhibit Cytotoxic Effects on Primary Fibroblasts
3.2. Genistein Reduces B Virus Spread and Replication in a Dose-Dependent Manner
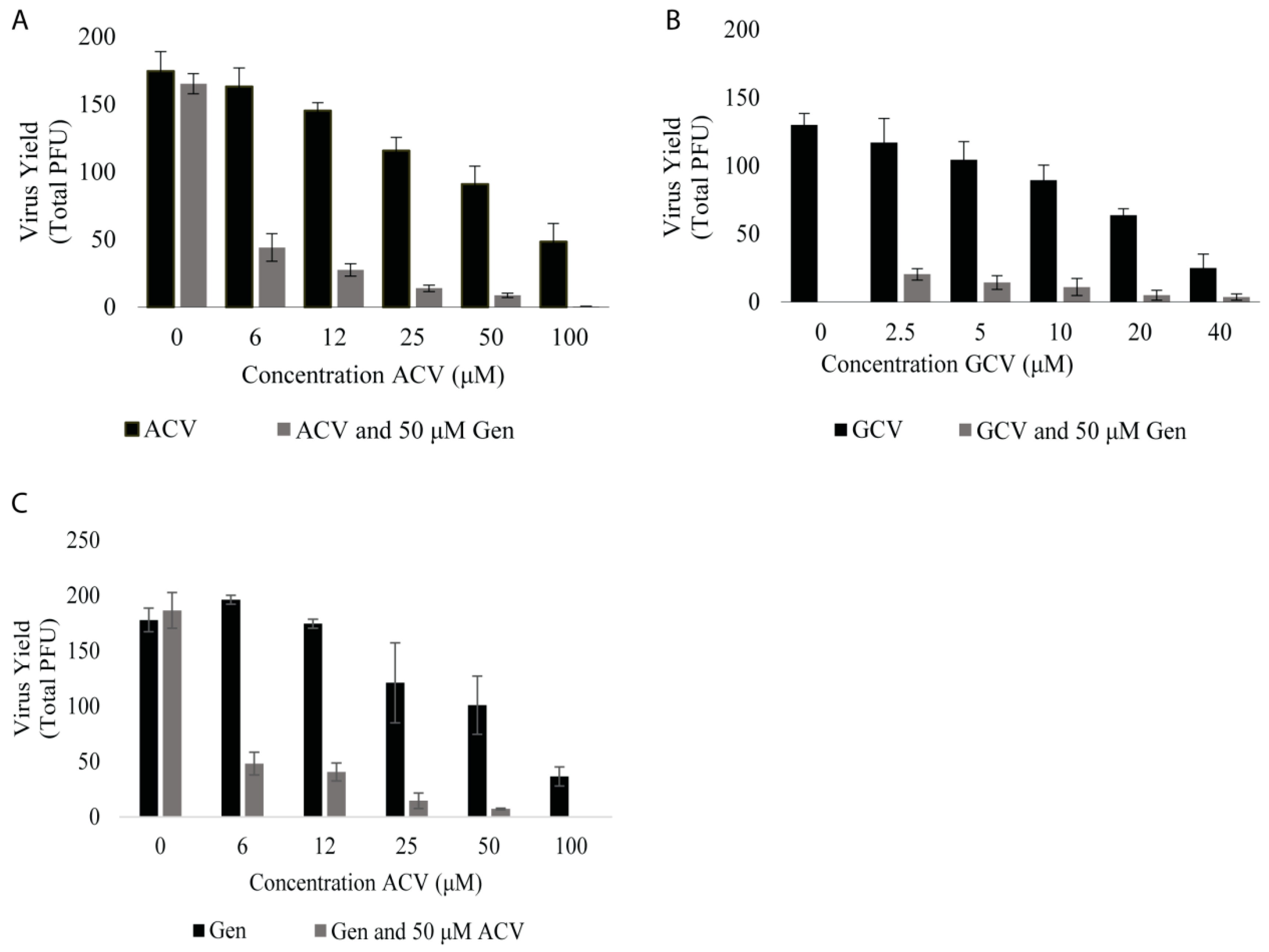
3.3. Genistein Reduces Effective Dose of Acyclovir and Ganciclovir against B Virus
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cohen, J.I.; Davenport, D.S.; Stewart, J.A.; Deitchman, S.; Hilliard, J.K.; Chapman, L.E.; the B Virus Working Group. Recommendations for Prevention of and Therapy for Exposure to B Virus (Cercopithecine Herpesvirus1). Clin. Infect. Dis. 2002, 35, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Eberle, R.; Hilliard, J. The simian herpesviruses. Infect Agents Dis. 1995, 4, 55–70. [Google Scholar] [PubMed]
- Hilliard, J.K.; Whitley, R.J. Cercopithecine Herpesvirus (B Virus). In Fields Virology; Whitley, R.J., Griffen, D., Howely, P., Knipe, D., Lamb, R., Martin, M., Philiadelphia, Lippincott Williams and Wilkins, Eds.; Philiadelphia: Linppincott Williams and Wilkins: New York, NY, USA, 2006; pp. 2835–2847. [Google Scholar]
- E Palmer, A. B virus, Herpesvirus simiae: historical perspective. J. Med Primatol. 1987, 16, 99–130. [Google Scholar]
- Weigler, B.J. Biology of B Virus in Macaque and Human Hosts: A Review. Clin. Infect. Dis. 1992, 14, 555–567. [Google Scholar] [CrossRef]
- De Clercq, E.; Field, H.J. Antiviral prodrugs–the development of successful prodrug strategies for antiviral chemotherapy. Br. J. Pharmacol. 2006, 147, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Elion, G.B.; Furman, P.A.; Fyfe, J.A.; Miranda, P.D.; Beauchamp, L.; Schaeffer, H.J. Selectivity of action of an antiherpetic agent, 9-(2-hydroxyethoxymethyl)guanine. Proc. Natl. Acad. Sci. USA 1977, 74, 5716–5720. [Google Scholar] [CrossRef] [Green Version]
- Zwartouw, H.T.; Humphreys, C.R.; Collins, P. Oral chemotherapy of fatal B virus (herpesvirus simiae) infection. Antivir. Res. 1989, 11, 275–283. [Google Scholar] [CrossRef]
- De Miranda, P.; Whitley, R.J.; Blum, M.R.; Keeney, R.E.; Barton, N.; Cocchetto, D.M.; Good, S.; Hemstreet, G.P., 3rd; Kirk, L.E.; Page, D.A.; et al. Acyclovir kinetics after intravenous infusion. Clin. Pharmacol. Ther. 1979, 26, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Svennerholm, B.; Appelgren, L.; Lycke, J.; Andersen, O.; Dahlöf, C. Acyclovir concentrations in serum and cerebrospinal fluid at steady state. J. Antimicrob. Chemother. 1989, 24, 947–954. [Google Scholar]
- Brigden, D.; Rosling, A.E.; Woods, N.C. Renal function after acyclovir intravenous injection. Am. J. Med. 1982, 73, 182–185. [Google Scholar] [CrossRef]
- Focher, F.; Lossani, A.; Verri, A.; Spadari, S.; Maioli, A.; Gambino, J.J.; Wright, G.E.; Eberle, R.; Black, D.H.; Medveczky, P.; et al. Sensitivity of Monkey B Virus (Cercopithecine herpesvirus 1) to Antiviral Drugs: Role of Thymidine Kinase in Antiviral Activities of Substrate Analogs and Acyclonucleosides. Antimicrob. Agents Chemother. 2007, 51, 2028–2034. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Morroni, J.; Wilcox, C. Lack of effect of treatment with penciclovir or acyclovir on the establishment of latent HSV-1 in primary sensory neurons in culture. Antivir. Res. 2001, 52, 19–24. [Google Scholar] [CrossRef]
- Morfin, F.; Thouvenot, D. Herpes simplex virus resistance to antiviral drugs. J. Clin. Virol. 2003, 26, 29–37. [Google Scholar] [CrossRef]
- Martin, J.; Crotty, S.; Warren, P.; Nelson, P. Does an apple a day keep the doctor away because a phytoestrogen a day keeps the virus at bay? A review of the anti-viral properties of phytoestrogens. Phytochemistry 2007, 68, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar] [PubMed]
- Food and Drug Administration. FDA Approves New Health Claim for Soy Protein and Coronary Heart Disease. FDA Talk Paper. Available online: http://www.fda.gov/bbs/topics/ANSWERS/ANS00980.html.1999 (accessed on 30 May 2019).
- U.S. National Library of Medicine. Available online: www.clinicaltrials.gov (accessed on 28 April 2019).
- Greiner, L.L.; Stahly, T.S.; Stabel, T.J. The effect of dietary soy genistein on pig growth and viral replication during a viral challenge. J. Sci. 2001, 79, 1272. [Google Scholar] [CrossRef]
- Akula, S.M.; Hurley, D.J.; Wixon, R.L.; Wang, C.; Chase, C.C. Effect of genistein on replication of bovine herpesvirus type 1. Am. J.Vet. Res. 2002, 63, 1124–1128. [Google Scholar] [CrossRef]
- Lyu, S.Y.; Rhim, J.Y.; Park, W.B. Antiherpetic activities of flavonoids against herpes simplex virus type 1 (HSV-1) and type 2 (HSV-2) in vitro. Arch. Pharmacal Res. 2005, 28, 1293–1301. [Google Scholar] [CrossRef]
- Evers, D.L.; Chao, C.F.; Wang, X.; Zhang, Z.; Huong, S.M.; Huang, E.S. Human cytomegalovirus-inhibitory flavonoids: studies on antiviral activity and mechanism of action. Antivir. Res. 2005, 68, 124–134. [Google Scholar] [CrossRef]
- Engel, S.; Heger, T.; Mancini, R.; Herzog, F.; Kartenbeck, J.; Hayer, A.; Helenius, A. Role of endosomes in simian virus 40 entry and infection. J. Virol. 2011, 85, 4198–4211. [Google Scholar] [CrossRef] [PubMed]
- Stantchev, T.S.; Markovic, I.; Telford, W.G.; Clouse, K.A.; Broder, C.C. The tyrosine kinase inhibitor genistein blocks HIV-1 infection in primary human macrophages. Virus Res. 2007, 123, 178–189. [Google Scholar] [CrossRef] [Green Version]
- Arabyan, E.; Hakobyan, A.; Kotsinyan, A.; Karalyan, Z.; Arakelov, V.; Arakelov, G.; Nazaryan, K.; Simonyan, A.; Aroutiounian, R.; Ferreira, F.; Zakaryan, H. Genistein inhibits African swine fever virus replication in vitro by disrupting viral DNA synthesis. Antivir. Res. 2018, 156, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Özçelik, B.; Kartal, M.; Orhan, I. Cytotoxicity, antiviral and antimicrobial activities of alkaloids, flavonoids, and phenolic acids. Pharm. Boil. 2011, 49, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Kubo, Y.; Ishimoto, A.; Amanuma, H. Genistein, a protein tyrosine kinase inhibitor, suppresses the fusogenicity of Moloney murine leukemia virus envelope protein in XC cells. Arch. Virol. 2003, 148, 1899–1914. [Google Scholar] [CrossRef]
- Querbes, W.; Benmerah, A.; Tosoni, D.; Di Fiore, P.P.; Atwood, W.J. A JC virus-induced signal is required for infection of glial cells by a clathrin- and eps15-dependent pathway. J. Virol. 2004, 78, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.; Braun, L.; Frew, T.; Hurley, D.; Rowland, R.; Chase, C.; Chase, C. A Role for Bovine Herpesvirus 1 (BHV-1) Glycoprotein E (gE) Tyrosine Phosphorylation in Replication of BHV-1 Wild-Type Virus but Not BHV-1 gE Deletion Mutant Virus. Virology 2000, 268, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yura, Y.; Yoshida, H.; Sato, M. Inhibition of herpes simplex virus replication by genistein, an inhibitor of protein-tyrosine kinase. Arch. Virol. 1993, 132, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, U.; Bendik, I.; Flühmann, B. Bonistein (synthetic genistein), a food component in development for a bone health nutraceutical. J. Physiol. Pharmacol. 2005, 56, 79–95. [Google Scholar] [PubMed]
- Hirasawa, K.; Kim, A.; Han, H.-S.; Han, J.; Jun, H.-S.; Yoon, J.-W. Effect of p38 Mitogen-Activated Protein Kinase on the Replication of Encephalomyocarditis Virus. J. Virol. 2003, 77, 5649–5656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangoria, N.S.; Breau, W.C.; Anderson, H.A.; Cishek, D.M.; Norkin, L.C. Extracellular Simian Virus 40 Induces an ERK/MAP Kinase-independent Signalling Pathway that Activates Primary Response Genes and Promotes Virus Entry. J. Virol. 1996, 77, 2173–2182. [Google Scholar] [CrossRef] [PubMed]
- Vela, E.M.; Bowick, G.C.; Herzog, N.K.; Aronson, J.F. Genistein treatment of cells inhibits arenavirus infection. Antivir. Res. 2008, 77, 153–156. [Google Scholar] [CrossRef] [Green Version]
- Vella, C.; Festenstein, H. Coxsackievirus B4 infection of the mouse pancreas: the role of natural killer cells in the control of virus replication and resistance to infection. J. Virol. 1992, 73, 1379–1386. [Google Scholar] [CrossRef]
- Perelygina, L.; Patrusheva, I.; Hombaiah, S.; Zurkuhlen, H.; Wildes, M.J.; Patrushev, N.; Hilliard, J. Production of Herpes B Virus Recombinant Glycoproteins and Evaluation of Their Diagnostic Potential. J. Clin. Microbiol. 2005, 43, 620–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeCher, J.C. Georgia State University: Atlanta, GA, USA; Analysis of BrdU incorporation into human and macaque fibroblasts post treatment with increasing concentrations of genestein. Unpublished data. 2015. [Google Scholar]
- Elmore, D.; Eberle, R. Monkey B Virus (Cercopithecine herpesvirus 1). Comp. Med. 2008, 58, 11–21. [Google Scholar] [PubMed]
- Hayashi, K.; Hayashi, T.; Otsuka, H.; Takeda, Y. Antiviral activity of 5,6,7-trimethoxyflavone and its potentiation of the antiherpes activity of acyclovir. J. Antimicrob. Chemother. 1997, 39, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, M.; Nagasaka, K.; Hirabayashi, T.; Uyama, S.-I.; Sato, H.; Kageyama, T.; Kadota, S.; Ohyama, H.; Hozumi, T.; Namba, T.; et al. Efficacy of traditional herbal medicines in combination with acyclovir against herpes simplex virus type 1 infection in vitro and in vivo. Antivir. Res. 1995, 27, 19–37. [Google Scholar] [CrossRef]
- Mucsi, I.; Gyulai, Z.; Béládi, I. Combined effects of flavonoids and acyclovir against herpesviruses in cell cultures. Acta Microbiol. Hung. 1992, 39, 137–147. [Google Scholar]
- Schang, L.M.; Bantly, A.; Knockaert, M.; Shaheen, F.; Meijer, L.; Malim, M.H.; Gray, N.S.; Schaffer, P.A. Pharmacological Cyclin-Dependent Kinase Inhibitors Inhibit Replication of Wild-Type and Drug-Resistant Strains of Herpes Simplex Virus and Human Immunodeficiency Virus Type 1 by Targeting Cellular, Not Viral, Proteins. J. Virol. 2002, 76, 7874–7882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.L.; Tom, P.; O’Brien, W.J. Combined effects of interferon-alpha and acyclovir on herpes simplex virus type 1 DNA polymerase and alkaline DNase. Antivir. Res. 1998, 38, 95–106. [Google Scholar] [CrossRef]
- Barnard, D.; Smee, D.; Huffman, J.; Meyerson, L.; Sidwell, R. Antiherpesvirus Activity and Mode of Action of SP-303, a Novel Plant Flavonoid. Chemotherapy 1993, 39, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Docherty, J.J.; Smith, J.S.; Fu, M.M.; Stoner, T.; Booth, T. Effect of topically applied resveratrol on cutaneous herpes simplex virus infections in hairless mice. Antivir. Res. 2004, 61, 19–26. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agent | Concentration (µM) |
|---|---|
| Genistein | 56 |
| Acyclovir | 55 |
| Acyclovir + Genistein (50 µM) | 3.9 |
| Ganciclovir | 16 |
| Ganciclovir + Genistein (50 µM) | 0.002 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
LeCher, J.C.; Diep, N.; Krug, P.W.; Hilliard, J.K. Genistein Has Antiviral Activity against Herpes B Virus and Acts Synergistically with Antiviral Treatments to Reduce Effective Dose. Viruses 2019, 11, 499. https://doi.org/10.3390/v11060499
LeCher JC, Diep N, Krug PW, Hilliard JK. Genistein Has Antiviral Activity against Herpes B Virus and Acts Synergistically with Antiviral Treatments to Reduce Effective Dose. Viruses. 2019; 11(6):499. https://doi.org/10.3390/v11060499
Chicago/Turabian StyleLeCher, Julia C., Nga Diep, Peter W. Krug, and Julia K. Hilliard. 2019. "Genistein Has Antiviral Activity against Herpes B Virus and Acts Synergistically with Antiviral Treatments to Reduce Effective Dose" Viruses 11, no. 6: 499. https://doi.org/10.3390/v11060499
APA StyleLeCher, J. C., Diep, N., Krug, P. W., & Hilliard, J. K. (2019). Genistein Has Antiviral Activity against Herpes B Virus and Acts Synergistically with Antiviral Treatments to Reduce Effective Dose. Viruses, 11(6), 499. https://doi.org/10.3390/v11060499