A Highly Sensitive Method to Detect Avocado Sunblotch Viroid for the Maintenance of Infection-Free Avocado Germplasm Collections


Abstract
:1. Introduction
2. Materials and Methods
2.1. Avocado Trees
2.2. Viroid Indexing
2.2.1. Sampling
2.2.2. Tissue Disruption
2.3. RNA Extraction
2.4. Primers and Probes
2.5. Reverse Transcription and Pre-amplification
2.6. TaqMan Real-Time PCR Assays
2.7. Roguing Infected Trees
2.8. Creation of a Backup Germplasm Collection
3. Results
3.1. Assay Design and Testing
3.2. Sensitivity Determination
3.3. Distribution of ASBVd in A Single Tree
3.4. Pooling Samples and Multiplex of ASBVd and GAPDH
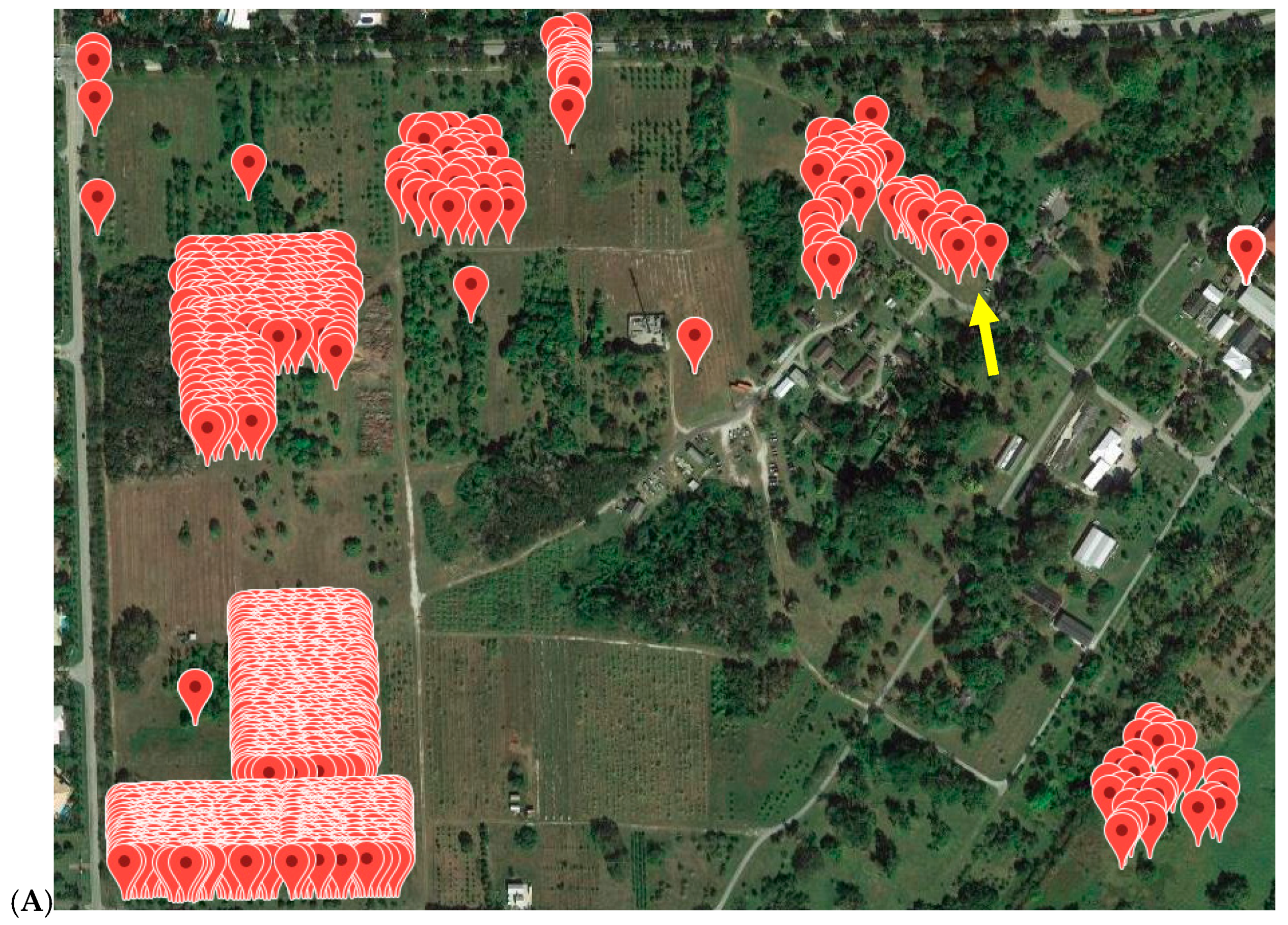
3.5. Application
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Di Serio, F.; Flores, R.; Verhoeven, J.T.J.; Li, S.-F.; Pallás, V.; Randles, J.W.; Sano, T.; Vidalakis, G.; Owens, R.A. Current status of viroid taxonomy. Arch. Virol. 2014, 159, 3467–3478. [Google Scholar] [CrossRef] [PubMed]
- Saucedo-Carabez, J.R.; Téliz-Ortiz, D.; Ochoa-Ascencio, S.; Ochoa-Martínez, D.; Vallejo-Pérez, M.R.; Beltrán-Peña, H. Effect of Avocado sunblotch viroid (ASBVd) on avocado yield in Michoacan, Mexico. Eur. J. Plant Pathol. 2014, 13, 799–805. [Google Scholar] [CrossRef]
- Kuhn, D.N.; Geering, A.D.W.; Dixon, J. Avocado sunblotch viroid. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press Ltd-Elsevier Science Ltd.: London, UK, 2017; pp. 297–305. [Google Scholar] [CrossRef]
- Desjardins, P.R.; Drake, R.J.; Sasaki, P.J.; Atkins, E.L.; Bergh, B.O. Pollen transmission of of avocado sunblotch viroid and the fate of the pollen recipient tree. Phytopathology 1984, 74, 845. [Google Scholar]
- Suarez, I.E.; Schnell, R.A.; Kuhn, D.N.; Litz, R.E. Recovery and indexing of avocado plants (persea americana) from embryogenic nucellar cultures of an avocado sunblotch viroid-infected tree. Plant Cell Tissue Organ Cult. 2006, 84, 27–37. [Google Scholar] [CrossRef]
- Suarez, I.E.; Schnell, R.A.; Kuhn, D.N.; Litz, R.E. Micrografting of asbvd-infected avocado (persea americana) plants. Plant Cell Tissue Organ Cult. 2005, 80, 179–185. [Google Scholar] [CrossRef]
- Schnell, R.J.; Tondo, C.L.; Kuhn, D.N.; Winterstein, M.C.; Ayala-Silva, T.; Moore, J.M. Spatial analysis of avocado sunblotch disease in an avocado germplasm collection. J. Phytopathol. 2011, 159, 773–781. [Google Scholar] [CrossRef]
- Olano, C.T.; Schnell, R.J.; Kuhn, D.N. Current status of asbvd infection among avocado accessions in the national germplasm collection. Proc. Fla. State Hort. Soc. 2002, 115, 280–282. [Google Scholar]
- Running, C.; Schnell, R.; Kuhn, D.N. Detection of avocado sunblotch viroid and estimation of infection among accessions in the national germplasm collection for avocado. Proc. Fla. State Hort. Soc. 1996, 109, 235–236. [Google Scholar]
- Ploetz, R.; Peña, J.; Smith, J.; Dreaden, T.; Crane, J.; Schubert, T.; Dixon, W. Laurel wilt, caused by raffaelea lauricola, is confirmed in miami-dade county, center of florida’s commercial avocado production. Plant Dis. 2011, 95, 1589. [Google Scholar] [CrossRef] [PubMed]
- Schnell, R.; Kuhn, D.; Ronning, C.; Harkins, D. Application of rt-pcr for indexing avocado sunblotch viroid. Plant Dis. 1997, 81, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Da Graça, J.; Mason, T. Detection of avocado sunblotch viroid in flower buds by polyacrylamide gel electrophoresis. J. Phytopathol. 1983, 108, 262–266. [Google Scholar] [CrossRef]
- Semancik, J.; Szychowski, J. Avocado sunblotch disease: A persistent viroid infection in which variants are associated with differential symptoms. J. Gen. Virol. 1994, 75, 1543–1549. [Google Scholar] [CrossRef] [PubMed]
- Morey-León, G.; Ortega-Ramirez, E.; Julca-Chunga, C.; Santos-Chanta, C.; Graterol-Caldera, L.; Mialhe, E. The detection of avocado sunblotch viroid in avocado using a real-time reverse transcriptase polymerase chain reaction. BioTechnologia 2018, 99, 99–107. [Google Scholar] [CrossRef]
- Olano, C.; Borrone, J.; Brown, J.S.; Violi, H.; Ploetz, R.; Schnell, R. Development of mapping populations for avocado. Proc. Fla. State Hort. Soc. 2007, 120, 26–29. [Google Scholar]
- Tondo, C.L.; Schnell, R.J.; Kuhn, D.N. Results of the 2009 asbvd survey of avocado accessions of the national germplasm collection in florida. Proc. Fla. State Hort. Soc. 2010, 123, 5–7. [Google Scholar]
- Geering, A.D.W.; Steele, V.; Kopittke, R. Final report for Horticulture Australia Limited Project AV03009: Development of Avocado sunblotch viroid indexing protocols for the avocado nursery industry; University of Queensland: St Lucia, Australia, 2006. [Google Scholar]
- Randles, J.W.; Hanold, D.; Harding, R.M.; Skrzeckzkowski, L.; Whattam, M. Viroids in Australasia. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, Australia, 2003. [Google Scholar]
- Ainsworth, C. Isolation of RNA from floral tissue ofrumex acetosa (sorrel). Plant Mol. Biol. Report. 1994, 12, 198–203. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Population | Original Source 1 | Current Location 1 | Number Individuals Sampled | Number Individuals with Inconsistent Results from Previous Assay |
|---|---|---|---|---|
| Backup germplasm collection | SHRS ARS | FDWSRU ARS Ft. Detrick, Maryland | 131 | 10 |
| Avocado germplasm collection | SHRS ARS | SHRS ARS Miami, Florida | 383 | 50 |
| Avocado mapping population 2 | SHRS ARS | SHRS ARS Miami, Florida | 50 | NPA 3 |
| Backup germplasm collection | SHRS ARS | PBARC ARS Hilo, Hawaii | 102 | NPA |
| Primer Name | Primer Color in Figure 1 | Primer Sequence (5′→ 3′) | Position in ASBVd Genome | Reaction | Reference |
|---|---|---|---|---|---|
| GAPTM-F1 | TGGAGTGGACAGTGGTCATCAG | RT and TaqMan | Geering, ADW [17] | ||
| GAPTM-R1new | CCCATTGGCCAAGGTGATC | RT and TaqMan | This study | ||
| GAPDHTM-probe | [VIC]-CCCTCAACAATGCC-MGBNFQ | TaqMan | Geering, ADW [17] | ||
| SB1-F1 | Red | TGGGAAGAACACTGATGAG | 180–198 | RT and preAmp | Semancik, JS. [13] |
| SB1-R1 | Red | TCTTTCCCTGAAGAGACGA | 179–161 | RT and preAmp | Semancik, JS. [13] |
| ASBTM-F1 | Blue | TTCCGACTCTGAGTTTCGACTT | 66–87 | TaqMan | Geering, ADW [17] |
| AVFL1 | Green | CAAGAGATTGAAGACGAGTGAACTA | 179–155 | TaqMan | Randles et al. [18] |
| ASBTM-probe | Hatched | [6FAM]TTCCGACTCTGAGTTTCGACTT-MGBNFQ | 89–107 | TaqMan | Geering, ADW [17] |
| ASB-F1 | Purple | GTGAGAGAAGGAGGAGT | 88–104 | Schnell et al. [11] | |
| ASB-R1 | Purple | AAGTCGAAACTCAGAGTCGG | 87–68 | Schnell et al. [11] | |
| AVFL2 | Green | ATCACTTCGTCTCTTCAGGGAAAGA | 130–154 | Randles et al. [18] | |
| ASBTM-R1 | Blue | GTTCTTCCCATCTTTCCCTGA | 189–168 | Geering, ADW [17] |
| Dilution | CT avg | CT SD |
|---|---|---|
| 1:103 | 17.6 | 2.0 |
| 1:104 | 24.3 | 1.9 |
| 1:105 | 25.5 | 1.4 |
| 1:106 | 29.5 | 2.4 |
| 1:107 | 31.1 | 2.4 |
| 1:108 | No amp | 0.0 |
| 1:109 | No amp | 0.0 |
| Sample ID | Avg CT | Avg CT all Locations * | FDWSRU Graft av CT | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | Location (SHRS) | Genotype ID | East | North | South | Top North | Top South | West | ||
| AYCOCK RED NO. 19 | W3-1-08-01 | PC | 22.3 | 9.3 | 7.3 | 8.4 | 8.2 | 8.0 | 10.6 | NG |
| CELLON’S HAWAII SDLG | W3-1-02-01 | CFN01 | 26.9 | 25.0 | 39.4 | 33.8 | 35.6 | No amp | 33.6 | NG |
| SHARWIL | WA2-12-37 | CFN19 | 26.9 | 25.0 | 22.9 | 23.9 | 25.2 | 21.9 | 24.3 | NG |
| JOSE ANTONIO | WA2-20-32.2 | CFN22 | No amp | No amp | No amp | 17.9 | 24.0 | No amp | 34.3 | NG |
| R06-T05 | WB4-02-13 | FtD3 | No amp | No amp | 18.0 | No amp | No amp | 11.5 | 32.2 | 18.3 |
| LA PISCINA | WB3-18-08 | FtD30 | 22.3 | 21.8 | 23.9 | No amp | 24.8 | 34.0 | 28.0 | 18.9 |
| DARIAN | WB3-19-11 | FtD36 | 20.0 | 22.3 | No amp | 28.2 | 17.3 | No amp | 28.3 | 11.0 |
| SEMIL 43 | WA2-13-41 | FtD96 | 22.2 | 20.5 | 22.2 | 16.4 | 17.3 | 12.7 | 18.6 | 29.1 |
| P. NUBIGENA | WB3-10-03 | FtD97 | 16.2 | 23.3 | 24.8 | No amp | 22.1 | 20.7 | 24.7 | 8.2 |
| PIC 9615 | WB3-13-10 | FtD105 | 19.7 | No amp | No amp | No amp | 13.4 | No amp | 32.8 | 16.2 |
| PINKERTON | WB4-09-01 | FtD107 | No amp | No amp | 23.2 | No amp | 22.2 | No amp | 34.9 | 6.1 |
| DADE SDLG | WB4-04-17 | FtD110 | No amp | 18.5 | 38.3 | No amp | No amp | No amp | 36.8 | 7.8 |
| HONALINDO 1 | WB3-10-07 | FtD117 | 19.2 | 21.6 | No amp | 9.5 | 9.0 | No amp | 23.5 | 8.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuhn, D.N.; Freeman, B.; Geering, A.; Chambers, A.H. A Highly Sensitive Method to Detect Avocado Sunblotch Viroid for the Maintenance of Infection-Free Avocado Germplasm Collections. Viruses 2019, 11, 512. https://doi.org/10.3390/v11060512
Kuhn DN, Freeman B, Geering A, Chambers AH. A Highly Sensitive Method to Detect Avocado Sunblotch Viroid for the Maintenance of Infection-Free Avocado Germplasm Collections. Viruses. 2019; 11(6):512. https://doi.org/10.3390/v11060512
Chicago/Turabian StyleKuhn, David N., Barbie Freeman, Andrew Geering, and Alan H. Chambers. 2019. "A Highly Sensitive Method to Detect Avocado Sunblotch Viroid for the Maintenance of Infection-Free Avocado Germplasm Collections" Viruses 11, no. 6: 512. https://doi.org/10.3390/v11060512
APA StyleKuhn, D. N., Freeman, B., Geering, A., & Chambers, A. H. (2019). A Highly Sensitive Method to Detect Avocado Sunblotch Viroid for the Maintenance of Infection-Free Avocado Germplasm Collections. Viruses, 11(6), 512. https://doi.org/10.3390/v11060512