Mast Cells and Natural Killer Cells—A Potentially Critical Interaction

Abstract
:1. Mast Cells
2. NK Cell–Mast Cell Interactions
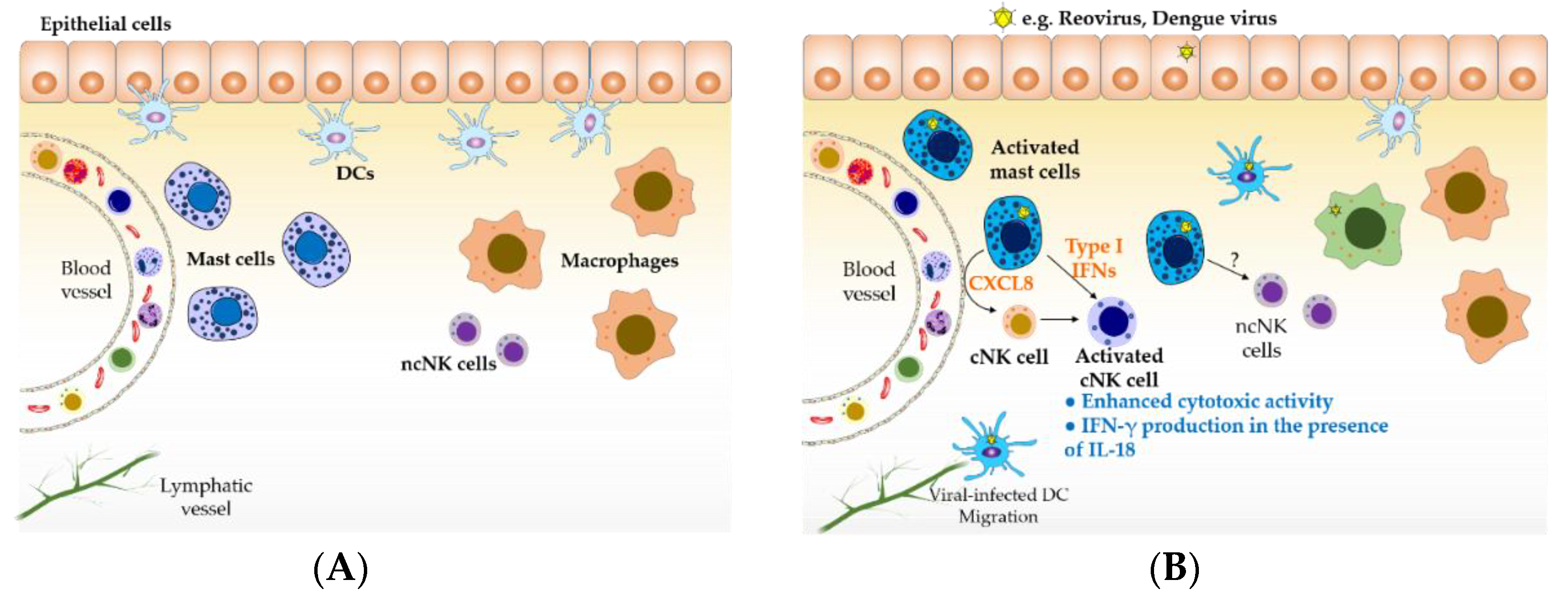
2.1. Viral Infection
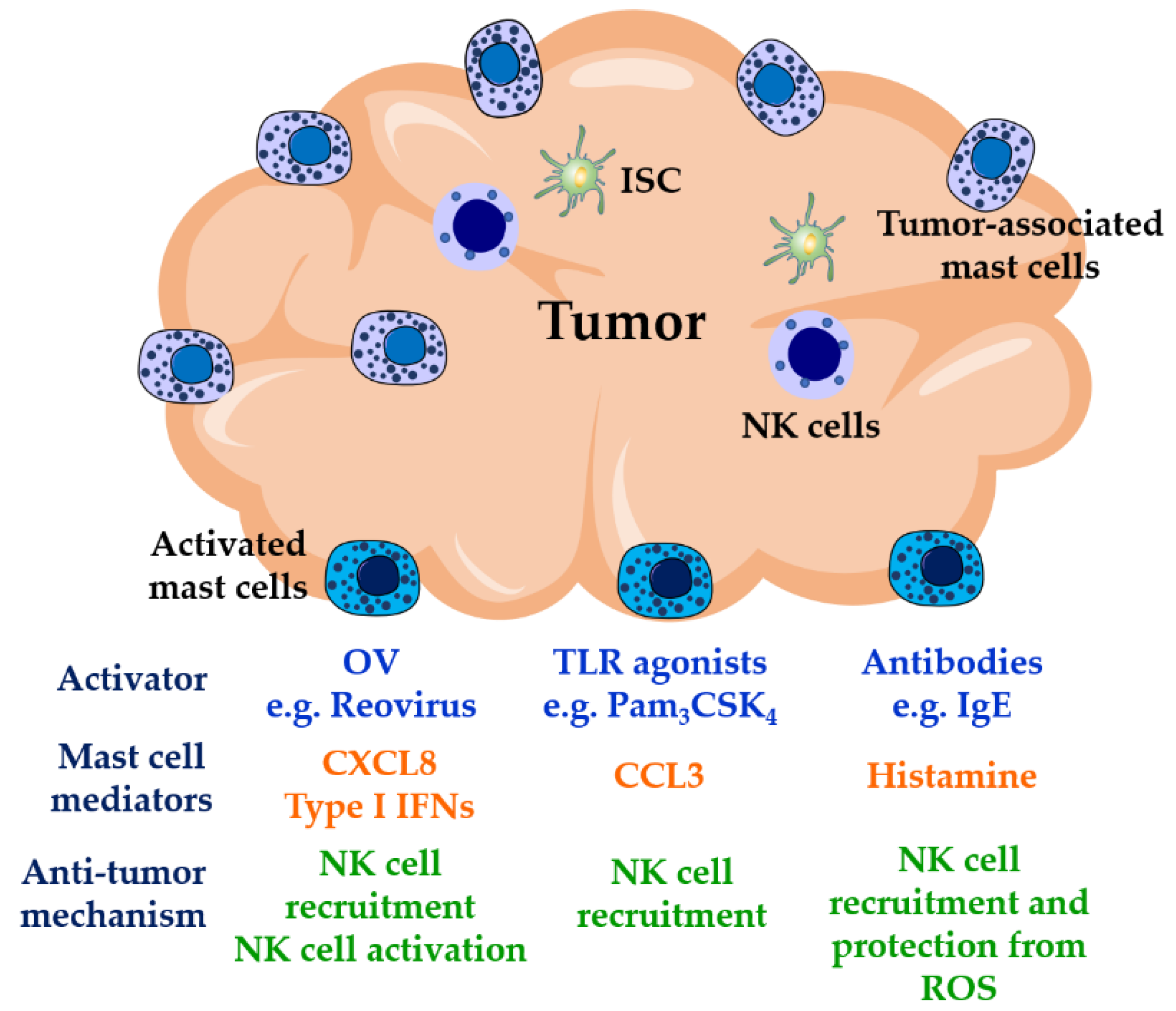
2.2. Cancer
2.3. Allergic Asthma
3. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Rivera, J.; Fierro, N.A.; Olivera, A.; Suzuki, R. New insights on mast cell activation via the high affinity receptor for ige. Adv. Immunol. 2008, 98, 85–120. [Google Scholar] [PubMed]
- Lorentz, A.; Wilke, M.; Sellge, G.; Worthmann, H.; Klempnauer, J.; Manns, M.P.; Bischoff, S.C. IL-4-induced priming of human intestinal mast cells for enhanced survival and th2 cytokine generation is reversible and associated with increased activity of erk1/2 and c-fos. J. Immunol. 2005, 174, 6751–6756. [Google Scholar] [CrossRef] [PubMed]
- Shelburne, C.P.; Ryan, J.J. The role of th2 cytokines in mast cell homeostasis. Immunol. Rev. 2001, 179, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Oldford, S.A.; Salsman, S.P.; Portales-Cervantes, L.; Alyazidi, R.; Anderson, R.; Haidl, I.D.; Marshall, J.S. Interferon alpha2 and interferon gamma induce the degranulation independent production of vegf-a and il-1 receptor antagonist and other mediators from human mast cells. Immun. Inflamm. Dis. 2018, 6, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Tore, F.; Tuncel, N. Mast cells: Target and source of neuropeptides. Curr. Pharm. Des. 2009, 15, 3433–3445. [Google Scholar] [CrossRef] [PubMed]
- Enoksson, M.; Lyberg, K.; Moller-Westerberg, C.; Fallon, P.G.; Nilsson, G.; Lunderius-Andersson, C. Mast cells as sensors of cell injury through il-33 recognition. J. Immunol. 2011, 186, 2523–2528. [Google Scholar] [CrossRef]
- Gulliksson, M.; Carvalho, R.F.; Ulleras, E.; Nilsson, G. Mast cell survival and mediator secretion in response to hypoxia. PLoS ONE 2010, 5, e12360. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M. Ige and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Amin, K. The role of mast cells in allergic inflammation. Respir. Med. 2012, 106, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Akahoshi, M.; Song, C.H.; Piliponsky, A.M.; Metz, M.; Guzzetta, A.; Abrink, M.; Schlenner, S.M.; Feyerabend, T.B.; Rodewald, H.R.; Pejler, G.; et al. Mast cell chymase reduces the toxicity of gila monster venom, scorpion venom, and vasoactive intestinal polypeptide in mice. J. Clin. Invest. 2011, 121, 4180–4191. [Google Scholar] [CrossRef]
- Grujic, M.; Paivandy, A.; Gustafson, A.M.; Thomsen, A.R.; Ohrvik, H.; Pejler, G. The combined action of mast cell chymase, tryptase and carboxypeptidase a3 protects against melanoma colonization of the lung. Oncotarget 2017, 8, 25066–25079. [Google Scholar] [CrossRef] [PubMed]
- Marichal, T.; Starkl, P.; Reber, L.L.; Kalesnikoff, J.; Oettgen, H.C.; Tsai, M.; Metz, M.; Galli, S.J. A beneficial role for immunoglobulin e in host defense against honeybee venom. Immunity 2013, 39, 963–975. [Google Scholar] [CrossRef]
- Roy, A.; Ganesh, G.; Sippola, H.; Bolin, S.; Sawesi, O.; Dagalv, A.; Schlenner, S.M.; Feyerabend, T.; Rodewald, H.R.; Kjellen, L.; et al. Mast cell chymase degrades the alarmins heat shock protein 70, biglycan, hmgb1, and interleukin-33 (IL-33) and limits danger-induced inflammation. J. Biol. Chem. 2014, 289, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.Y.; Reed, N.D.; Crowle, P.K. Delayed expulsion of adult trichinella spiralis by mast cell-deficient w/wv mice. Infect. Immun. 1983, 41, 445–447. [Google Scholar] [PubMed]
- Hepworth, M.R.; Danilowicz-Luebert, E.; Rausch, S.; Metz, M.; Klotz, C.; Maurer, M.; Hartmann, S. Mast cells orchestrate type 2 immunity to helminths through regulation of tissue-derived cytokines. Proc. Natl. Acad. Sci. USA 2012, 109, 6644–6649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronnberg, E.; Guss, B.; Pejler, G. Infection of mast cells with live streptococci causes a toll-like receptor 2- and cell-cell contact-dependent cytokine and chemokine response. Infect. Immun. 2010, 78, 854–864. [Google Scholar] [CrossRef] [PubMed]
- Supajatura, V.; Ushio, H.; Nakao, A.; Okumura, K.; Ra, C.; Ogawa, H. Protective roles of mast cells against enterobacterial infection are mediated by toll-like receptor 4. J. Immunol. 2001, 167, 2250–2256. [Google Scholar] [CrossRef] [PubMed]
- Malaviya, R.; Ikeda, T.; Ross, E.; Abraham, S.N. Mast cell modulation of neutrophil influx and bacterial clearance at sites of infection through tnf-alpha. Nature 1996, 381, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Dawicki, W.; Jawdat, D.W.; Xu, N.; Marshall, J.S. Mast cells, histamine, and il-6 regulate the selective influx of dendritic cell subsets into an inflamed lymph node. J. Immunol. 2010, 184, 2116–2123. [Google Scholar] [CrossRef]
- Merluzzi, S.; Frossi, B.; Gri, G.; Parusso, S.; Tripodo, C.; Pucillo, C. Mast cells enhance proliferation of b lymphocytes and drive their differentiation toward iga-secreting plasma cells. Blood 2010, 115, 2810–2817. [Google Scholar] [CrossRef]
- Shelburne, C.P.; Nakano, H.; St John, A.L.; Chan, C.; McLachlan, J.B.; Gunn, M.D.; Staats, H.F.; Abraham, S.N. Mast cells augment adaptive immunity by orchestrating dendritic cell trafficking through infected tissues. Cell Host Microbe 2009, 6, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Nakae, S.; Suto, H.; Kakurai, M.; Sedgwick, J.D.; Tsai, M.; Galli, S.J. Mast cells enhance t cell activation: Importance of mast cell-derived tnf. Proc. Natl. Acad. Sci. USA 2005, 102, 6467–6472. [Google Scholar] [CrossRef] [PubMed]
- Zarnegar, B.; Westin, A.; Evangelidou, S.; Hallgren, J. Innate immunity induces the accumulation of lung mast cells during influenza infection. Front. Immunol. 2018, 9, 2288. [Google Scholar] [CrossRef]
- St John, A.L.; Rathore, A.P.; Raghavan, B.; Ng, M.L.; Abraham, S.N. Contributions of mast cells and vasoactive products, leukotrienes and chymase, to dengue virus-induced vascular leakage. Elife 2013, 2, e00481. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Li, W.; She, R.; Wang, D.; Han, D.; Li, R.; Ding, Y.; Yue, Z. Evidence for a role of mast cells in the mucosal injury induced by newcastle disease virus. Poult. Sci. 2009, 88, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.G.; Hermann, L.L.; Issekutz, A.C.; Marshall, J.S.; Rowter, D.; Al-Afif, A.; Anderson, R. Dengue virus infection of mast cells triggers endothelial cell activation. J. Virol. 2011, 85, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- St John, A.L.; Rathore, A.P.; Yap, H.; Ng, M.L.; Metcalfe, D.D.; Vasudevan, S.G.; Abraham, S.N. Immune surveillance by mast cells during dengue infection promotes natural killer (nk) and nkt-cell recruitment and viral clearance. Proc. Natl. Acad. Sci. USA 2011, 108, 9190–9195. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.M.; Issekutz, T.B.; Mohan, K.; Lee, P.W.; Shmulevitz, M.; Marshall, J.S. Human mast cell activation with virus-associated stimuli leads to the selective chemotaxis of natural killer cells by a cxcl8-dependent mechanism. Blood 2008, 111, 5467–5476. [Google Scholar] [CrossRef] [PubMed]
- Henney, C.S.; Kuribayashi, K.; Kern, D.E.; Gillis, S. Interleukin-2 augments natural killer cell activity. Nature 1981, 291, 335–338. [Google Scholar] [CrossRef]
- Boieri, M.; Ulvmoen, A.; Sudworth, A.; Lendrem, C.; Collin, M.; Dickinson, A.M.; Kveberg, L.; Inngjerdingen, M. Il-12, il-15, and il-18 pre-activated nk cells target resistant t cell acute lymphoblastic leukemia and delay leukemia development in vivo. Oncoimmunology 2017, 6, e1274478. [Google Scholar] [CrossRef]
- Borg, C.; Jalil, A.; Laderach, D.; Maruyama, K.; Wakasugi, H.; Charrier, S.; Ryffel, B.; Cambi, A.; Figdor, C.; Vainchenker, W.; et al. Nk cell activation by dendritic cells (dcs) requires the formation of a synapse leading to il-12 polarization in dcs. Blood 2004, 104, 3267–3275. [Google Scholar] [CrossRef] [PubMed]
- Ferlazzo, G.; Tsang, M.L.; Moretta, L.; Melioli, G.; Steinman, R.M.; Munz, C. Human dendritic cells activate resting natural killer (nk) cells and are recognized via the nkp30 receptor by activated nk cells. J. Exp. Med. 2002, 195, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Portales-Cervantes, L.; Haidl, I.D.; Lee, P.W.; Marshall, J.S. Virus-infected human mast cells enhance natural killer cell functions. J. Innate. Immun. 2017, 9, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Zwirner, N.W.; Domaica, C.I. Cytokine regulation of natural killer cell effector functions. Biofactors 2010, 36, 274–288. [Google Scholar] [CrossRef] [PubMed]
- Chun, Y.H.; Park, J.Y.; Lee, H.; Kim, H.S.; Won, S.; Joe, H.J.; Chung, W.J.; Yoon, J.S.; Kim, H.H.; Kim, J.T.; et al. Rhinovirus-infected epithelial cells produce more IL-8 and rantes compared with other respiratory viruses. Allergy Asthma Immunol. Res. 2013, 5, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Matsukura, S.; Kokubu, F.; Noda, H.; Tokunaga, H.; Adachi, M. Expression of IL-6, IL-8, and rantes on human bronchial epithelial cells, nci-h292, induced by influenza virus a. J. Allergy Clin. Immunol. 1996, 98, 1080–1087. [Google Scholar] [CrossRef]
- Vosskuhl, K.; Greten, T.F.; Manns, M.P.; Korangy, F.; Wedemeyer, J. Lipopolysaccharide-mediated mast cell activation induces ifn-gamma secretion by nk cells. J. Immunol. 2010, 185, 119–125. [Google Scholar] [CrossRef]
- Erick, T.K.; Brossay, L. Phenotype and functions of conventional and non-conventional nk cells. Curr. Opin. Immunol. 2016, 38, 67–74. [Google Scholar] [CrossRef]
- Ivanova, D.; Krempels, R.; Ryfe, J.; Weitzman, K.; Stephenson, D.; Gigley, J.P. Nk cells in mucosal defense against infection. Biomed. Res. Int. 2014, 2014, 413982. [Google Scholar] [CrossRef]
- Sojka, D.K.; Plougastel-Douglas, B.; Yang, L.; Pak-Wittel, M.A.; Artyomov, M.N.; Ivanova, Y.; Zhong, C.; Chase, J.M.; Rothman, P.B.; Yu, J.; et al. Tissue-resident natural killer (nk) cells are cell lineages distinct from thymic and conventional splenic nk cells. Elife 2014, 3, e01659. [Google Scholar] [CrossRef]
- Esposito, I.; Menicagli, M.; Funel, N.; Bergmann, F.; Boggi, U.; Mosca, F.; Bevilacqua, G.; Campani, D. Inflammatory cells contribute to the generation of an angiogenic phenotype in pancreatic ductal adenocarcinoma. J. Clin. Pathol. 2004, 57, 630–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Qian, J.; Zeng, F.; Li, S.; Guo, W.; Chen, L.; Li, G.; Zhang, Z.; Wang, Q.J.; Deng, F. Protein kinase ds promote tumor angiogenesis through mast cell recruitment and expression of angiogenic factors in prostate cancer microenvironment. J. Exp. Clin. Cancer Res 2019, 38, 114. [Google Scholar] [CrossRef] [PubMed]
- Gulubova, M.; Manolova, I.; Kyurkchiev, D.; Julianov, A.; Altunkova, I. Decrease in intrahepatic cd56+ lymphocytes in gastric and colorectal cancer patients with liver metastases. APMIS 2009, 117, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Takanami, I.; Takeuchi, K.; Naruke, M. Mast cell density is associated with angiogenesis and poor prognosis in pulmonary adenocarcinoma. Cancer 2000, 88, 2686–2692. [Google Scholar] [CrossRef]
- Ribatti, D.; Ennas, M.G.; Vacca, A.; Ferreli, F.; Nico, B.; Orru, S.; Sirigu, P. Tumor vascularity and tryptase-positive mast cells correlate with a poor prognosis in melanoma. Eur. J. Clin. Invest. 2003, 33, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Sammarco, G.; Gadaleta, C.D.; Zuccala, V.; Albayrak, E.; Patruno, R.; Milella, P.; Sacco, R.; Ammendola, M.; Ranieri, G. Tumor-associated macrophages and mast cells positive to tryptase are correlated with angiogenesis in surgically-treated gastric cancer patients. Int. J. Mol. Sci. 2018, 19, 1176. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, L.; Xia, Y.; Qi, Y.; Chen, Y.; Chen, L.; Zhang, P.; Kong, Y.; Qu, Y.; Wang, Z.; et al. Tumor infiltrating mast cells determine oncogenic hif-2alpha-conferred immune evasion in clear cell renal cell carcinoma. Cancer Immunol. Immunother. 2019, 68, 731–741. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, Y.; Xu, L.; Zhang, J.; Xie, H.; Fu, H.; Zhou, Q.; Chang, Y.; Dai, B.; Xu, J. Tumor stroma-infiltrating mast cells predict prognosis and adjuvant chemotherapeutic benefits in patients with muscle invasive bladder cancer. Oncoimmunology 2018, 7, e1474317. [Google Scholar] [CrossRef] [Green Version]
- Tu, J.F.; Pan, H.Y.; Ying, X.H.; Lou, J.; Ji, J.S.; Zou, H. Mast cells comprise the major of interleukin 17-producing cells and predict a poor prognosis in hepatocellular carcinoma. Medicine (Baltimore) 2016, 95, e3220. [Google Scholar] [CrossRef]
- Lv, Y.P.; Peng, L.S.; Wang, Q.H.; Chen, N.; Teng, Y.S.; Wang, T.T.; Mao, F.Y.; Zhang, J.Y.; Cheng, P.; Liu, Y.G.; et al. Degranulation of mast cells induced by gastric cancer-derived adrenomedullin prompts gastric cancer progression. Cell Death Dis. 2018, 9, 1034. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, B.; Li, D.; Lv, M.; Huang, C.; Shen, G.X.; Huang, B. Mast cells mobilize myeloid-derived suppressor cells and treg cells in tumor microenvironment via il-17 pathway in murine hepatocarcinoma model. PLoS ONE 2010, 5, e8922. [Google Scholar] [CrossRef] [PubMed]
- Danelli, L.; Frossi, B.; Gri, G.; Mion, F.; Guarnotta, C.; Bongiovanni, L.; Tripodo, C.; Mariuzzi, L.; Marzinotto, S.; Rigoni, A.; et al. Mast cells boost myeloid-derived suppressor cell activity and contribute to the development of tumor-favoring microenvironment. Cancer Immunol. Res. 2015, 3, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Siebenhaar, F.; Metz, M.; Maurer, M. Mast cells protect from skin tumor development and limit tumor growth during cutaneous de novo carcinogenesis in a kit-dependent mouse model. Exp. Dermatol. 2014, 23, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Rajput, A.B.; Turbin, D.A.; Cheang, M.C.; Voduc, D.K.; Leung, S.; Gelmon, K.A.; Gilks, C.B.; Huntsman, D.G. Stromal mast cells in invasive breast cancer are a marker of favourable prognosis: A study of 4,444 cases. Breast Cancer Res. Treat. 2008, 107, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Zhu, Y.; Wang, Y.; Liu, Z.; Zhang, J.; Wang, Z.; Xie, H.; Dai, B.; Xu, J.; Ye, D. Tumor infiltrating mast cells (tims) confers a marked survival advantage in nonmetastatic clear-cell renal cell carcinoma. Ann. Surg. Oncol. 2017, 24, 1435–1442. [Google Scholar] [CrossRef]
- Wang, B.; Li, L.; Liao, Y.; Li, J.; Yu, X.; Zhang, Y.; Xu, J.; Rao, H.; Chen, S.; Zhang, L.; et al. Mast cells expressing interleukin 17 in the muscularis propria predict a favorable prognosis in esophageal squamous cell carcinoma. Cancer Immunol. Immunother. 2013, 62, 1575–1585. [Google Scholar] [CrossRef]
- Chan, J.K.; Magistris, A.; Loizzi, V.; Lin, F.; Rutgers, J.; Osann, K.; DiSaia, P.J.; Samoszuk, M. Mast cell density, angiogenesis, blood clotting, and prognosis in women with advanced ovarian cancer. Gynecol. Oncol. 2005, 99, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Hedstrom, G.; Berglund, M.; Molin, D.; Fischer, M.; Nilsson, G.; Thunberg, U.; Book, M.; Sundstrom, C.; Rosenquist, R.; Roos, G.; et al. Mast cell infiltration is a favourable prognostic factor in diffuse large b-cell lymphoma. Br. J. Haematol. 2007, 138, 68–71. [Google Scholar] [CrossRef]
- Ali, G.; Boldrini, L.; Lucchi, M.; Mussi, A.; Corsi, V.; Fontanini, G. Tryptase mast cells in malignant pleural mesothelioma as an independent favorable prognostic factor. J. Thorac. Oncol. 2009, 4, 348–354. [Google Scholar] [CrossRef]
- Glajcar, A.; Szpor, J.; Pacek, A.; Tyrak, K.E.; Chan, F.; Streb, J.; Hodorowicz-Zaniewska, D.; Okon, K. The relationship between breast cancer molecular subtypes and mast cell populations in tumor microenvironment. Virchows Arch. 2017, 470, 505–515. [Google Scholar] [CrossRef]
- Rojas, I.G.; Spencer, M.L.; Martinez, A.; Maurelia, M.A.; Rudolph, M.I. Characterization of mast cell subpopulations in lip cancer. J. Oral. Pathol. Med. 2005, 34, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Belloni, A.S.; Nico, B.; Sala, G.; Longo, V.; Mangieri, D.; Crivellato, E.; Nussdorfer, G.G. Tryptase- and leptin-positive mast cells correlate with vascular density in uterine leiomyomas. Am. J. Obstet. Gynecol. 2007, 196, 470.e1–470.e7. [Google Scholar] [CrossRef] [PubMed]
- Rao, Q.; Chen, Y.; Yeh, C.R.; Ding, J.; Li, L.; Chang, C.; Yeh, S. Recruited mast cells in the tumor microenvironment enhance bladder cancer metastasis via modulation of erbeta/ccl2/ccr2 emt/mmp9 signals. Oncotarget 2016, 7, 7842–7855. [Google Scholar] [CrossRef] [PubMed]
- Soucek, L.; Lawlor, E.R.; Soto, D.; Shchors, K.; Swigart, L.B.; Evan, G.I. Mast cells are required for angiogenesis and macroscopic expansion of myc-induced pancreatic islet tumors. Nat. Med. 2007, 13, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Soucek, L.; Buggy, J.J.; Kortlever, R.; Adimoolam, S.; Monclus, H.A.; Allende, M.T.; Swigart, L.B.; Evan, G.I. Modeling pharmacological inhibition of mast cell degranulation as a therapy for insulinoma. Neoplasia 2011, 13, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Oldford, S.A.; Haidl, I.D.; Howatt, M.A.; Leiva, C.A.; Johnston, B.; Marshall, J.S. A critical role for mast cells and mast cell-derived il-6 in tlr2-mediated inhibition of tumor growth. J. Immunol. 2010, 185, 7067–7076. [Google Scholar] [CrossRef] [PubMed]
- Bodduluri, S.R.; Mathis, S.; Maturu, P.; Krishnan, E.; Satpathy, S.R.; Chilton, P.M.; Mitchell, T.C.; Lira, S.; Locati, M.; Mantovani, A.; et al. Mast cell-dependent cd8(+) t-cell recruitment mediates immune surveillance of intestinal tumors in apc(min/+) mice. Cancer Immunol. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Forward, N.A.; Furlong, S.J.; Yang, Y.; Lin, T.J.; Hoskin, D.W. Mast cells down-regulate cd4+cd25+ t regulatory cell suppressor function via histamine h1 receptor interaction. J. Immunol. 2009, 183, 3014–3022. [Google Scholar] [CrossRef]
- Roder, J.C.; Haliotis, T.; Klein, M.; Korec, S.; Jett, J.R.; Ortaldo, J.; Heberman, R.B.; Katz, P.; Fauci, A.S. A new immunodeficiency disorder in humans involving nk cells. Nature 1980, 284, 553–555. [Google Scholar] [CrossRef]
- Sullivan, J.L.; Byron, K.S.; Brewster, F.E.; Purtilo, D.T. Deficient natural killer cell activity in x-linked lymphoproliferative syndrome. Science 1980, 210, 543–545. [Google Scholar] [CrossRef]
- Imai, K.; Matsuyama, S.; Miyake, S.; Suga, K.; Nakachi, K. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: An 11-year follow-up study of a general population. Lancet 2000, 356, 1795–1799. [Google Scholar] [CrossRef]
- Strayer, D.R.; Carter, W.A.; Brodsky, I. Familial occurrence of breast cancer is associated with reduced natural killer cytotoxicity. Breast Cancer Res. Treat. 1986, 7, 187–192. [Google Scholar] [CrossRef]
- Hersey, P.; Edwards, A.; Honeyman, M.; McCarthy, W.H. Low natural-killer-cell activity in familial melanoma patients and their relatives. Br. J. Cancer 1979, 40, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef]
- Caretto, D.; Katzman, S.D.; Villarino, A.V.; Gallo, E.; Abbas, A.K. Cutting edge: The th1 response inhibits the generation of peripheral regulatory t cells. J. Immunol. 2010, 184, 30–34. [Google Scholar] [CrossRef]
- Olalekan, S.A.; Cao, Y.; Hamel, K.M.; Finnegan, A. B cells expressing ifn-gamma suppress treg-cell differentiation and promote autoimmune experimental arthritis. Eur. J. Immunol. 2015, 45, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Paik, P.K.; Chen, J.; Yarilina, A.; Kockeritz, L.; Lu, T.T.; Woodgett, J.R.; Ivashkiv, L.B. IFN-gamma suppresses il-10 production and synergizes with tlr2 by regulating gsk3 and creb/ap-1 proteins. Immunity 2006, 24, 563–574. [Google Scholar] [CrossRef]
- Wen, F.Q.; Liu, X.; Kobayashi, T.; Abe, S.; Fang, Q.; Kohyama, T.; Ertl, R.; Terasaki, Y.; Manouilova, L.; Rennard, S.I. Interferon-gamma inhibits transforming growth factor-beta production in human airway epithelial cells by targeting smads. Am. J. Respir. Cell Mol. Biol. 2004, 30, 816–822. [Google Scholar] [CrossRef]
- Kochupurakkal, B.S.; Wang, Z.C.; Hua, T.; Culhane, A.C.; Rodig, S.J.; Rajkovic-Molek, K.; Lazaro, J.B.; Richardson, A.L.; Biswas, D.K.; Iglehart, J.D. Rela-induced interferon response negatively regulates proliferation. PLoS ONE 2015, 10, e0140243. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Song, Z.; Chu, J.; Qu, X. Deficiency of interferon-gamma or its receptor promotes colorectal cancer development. J. Interferon Cytokine Res. 2015, 35, 273–280. [Google Scholar] [CrossRef]
- Zaidi, M.R. The interferon-gamma paradox in cancer. J. Interferon Cytokine Res. 2019, 39, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Salih, H.R.; Antropius, H.; Gieseke, F.; Lutz, S.Z.; Kanz, L.; Rammensee, H.G.; Steinle, A. Functional expression and release of ligands for the activating immunoreceptor nkg2d in leukemia. Blood 2003, 102, 1389–1396. [Google Scholar] [CrossRef] [PubMed]
- Doubrovina, E.S.; Doubrovin, M.M.; Vider, E.; Sisson, R.B.; O’Reilly, R.J.; Dupont, B.; Vyas, Y.M. Evasion from nk cell immunity by mhc class i chain-related molecules expressing colon adenocarcinoma. J. Immunol. 2003, 171, 6891–6899. [Google Scholar] [CrossRef] [PubMed]
- Vasievich, E.A.; Huang, L. The suppressive tumor microenvironment: A challenge in cancer immunotherapy. Mol. Pharm. 2011, 8, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Halama, N.; Braun, M.; Kahlert, C.; Spille, A.; Quack, C.; Rahbari, N.; Koch, M.; Weitz, J.; Kloor, M.; Zoernig, I.; et al. Natural killer cells are scarce in colorectal carcinoma tissue despite high levels of chemokines and cytokines. Clin. Cancer Res. 2011, 17, 678–689. [Google Scholar] [CrossRef]
- Takanami, I.; Takeuchi, K.; Giga, M. The prognostic value of natural killer cell infiltration in resected pulmonary adenocarcinoma. J. Thorac. Cardiovasc. Surg. 2001, 121, 1058–1063. [Google Scholar] [CrossRef] [Green Version]
- Ishigami, S.; Natsugoe, S.; Tokuda, K.; Nakajo, A.; Che, X.; Iwashige, H.; Aridome, K.; Hokita, S.; Aikou, T. Prognostic value of intratumoral natural killer cells in gastric carcinoma. Cancer 2000, 88, 577–583. [Google Scholar] [CrossRef]
- Sznurkowski, J.J.; Zawrocki, A.; Biernat, W. Subtypes of cytotoxic lymphocytes and natural killer cells infiltrating cancer nests correlate with prognosis in patients with vulvar squamous cell carcinoma. Cancer Immunol. Immunother. 2014, 63, 297–303. [Google Scholar] [CrossRef]
- Prestwich, R.J.; Errington, F.; Steele, L.P.; Ilett, E.J.; Morgan, R.S.; Harrington, K.J.; Pandha, H.S.; Selby, P.J.; Vile, R.G.; Melcher, A.A. Reciprocal human dendritic cell-natural killer cell interactions induce antitumor activity following tumor cell infection by oncolytic reovirus. J. Immunol. 2009, 183, 4312–4321. [Google Scholar] [CrossRef]
- Bhat, R.; Dempe, S.; Dinsart, C.; Rommelaere, J. Enhancement of nk cell antitumor responses using an oncolytic parvovirus. Int. J. Cancer 2011, 128, 908–919. [Google Scholar] [CrossRef]
- Chen, X.; Han, J.; Chu, J.; Zhang, L.; Zhang, J.; Chen, C.; Chen, L.; Wang, Y.; Wang, H.; Yi, L.; et al. A combinational therapy of egfr-car nk cells and oncolytic herpes simplex virus 1 for breast cancer brain metastases. Oncotarget 2016, 7, 27764–27777. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Rommelaere, J. Nk-cell-dependent killing of colon carcinoma cells is mediated by natural cytotoxicity receptors (ncrs) and stimulated by parvovirus infection of target cells. BMC Cancer 2013, 13, 367. [Google Scholar] [CrossRef] [PubMed]
- Bachanova, V.; Cooley, S.; Defor, T.E.; Verneris, M.R.; Zhang, B.; McKenna, D.H.; Curtsinger, J.; Panoskaltsis-Mortari, A.; Lewis, D.; Hippen, K.; et al. Clearance of acute myeloid leukemia by haploidentical natural killer cells is improved using il-2 diphtheria toxin fusion protein. Blood 2014, 123, 3855–3863. [Google Scholar] [CrossRef] [PubMed]
- Curti, A.; Ruggeri, L.; D’Addio, A.; Bontadini, A.; Dan, E.; Motta, M.R.; Trabanelli, S.; Giudice, V.; Urbani, E.; Martinelli, G.; et al. Successful transfer of alloreactive haploidentical kir ligand-mismatched natural killer cells after infusion in elderly high risk acute myeloid leukemia patients. Blood 2011, 118, 3273–3279. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.S.; Soignier, Y.; Panoskaltsis-Mortari, A.; McNearney, S.A.; Yun, G.H.; Fautsch, S.K.; McKenna, D.; Le, C.; Defor, T.E.; Burns, L.J.; et al. Successful adoptive transfer and in vivo expansion of human haploidentical nk cells in patients with cancer. Blood 2005, 105, 3051–3057. [Google Scholar] [CrossRef]
- Rubnitz, J.E.; Inaba, H.; Ribeiro, R.C.; Pounds, S.; Rooney, B.; Bell, T.; Pui, C.H.; Leung, W. Nkaml: A pilot study to determine the safety and feasibility of haploidentical natural killer cell transplantation in childhood acute myeloid leukemia. J. Clin. Oncol. 2010, 28, 955–959. [Google Scholar] [CrossRef]
- Brentjens, R.J.; Davila, M.L.; Riviere, I.; Park, J.; Wang, X.; Cowell, L.G.; Bartido, S.; Stefanski, J.; Taylor, C.; Olszewska, M.; et al. Cd19-targeted t cells rapidly induce molecular remissions in adults with chemotherapy-refractory acute lymphoblastic leukemia. Sci. Transl. Med. 2013, 5, 177ra138. [Google Scholar] [CrossRef]
- Maude, S.L.; Frey, N.; Shaw, P.A.; Aplenc, R.; Barrett, D.M.; Bunin, N.J.; Chew, A.; Gonzalez, V.E.; Zheng, Z.; Lacey, S.F.; et al. Chimeric antigen receptor t cells for sustained remissions in leukemia. N. Engl. J. Med. 2014, 371, 1507–1517. [Google Scholar] [CrossRef]
- Yu, Z.; Chan, M.K.; O-charoenrat, P.; Eisenberg, D.P.; Shah, J.P.; Singh, B.; Fong, Y.; Wong, R.J. Enhanced nectin-1 expression and herpes oncolytic sensitivity in highly migratory and invasive carcinoma. Clin. Cancer Res. 2005, 11, 4889–4897. [Google Scholar] [CrossRef]
- Anderson, B.D.; Nakamura, T.; Russell, S.J.; Peng, K.W. High cd46 receptor density determines preferential killing of tumor cells by oncolytic measles virus. Cancer Res. 2004, 64, 4919–4926. [Google Scholar] [CrossRef]
- Mansour, M.; Palese, P.; Zamarin, D. Oncolytic specificity of newcastle disease virus is mediated by selectivity for apoptosis-resistant cells. J. Virol. 2011, 85, 6015–6023. [Google Scholar] [CrossRef] [PubMed]
- Sborov, D.W.; Nuovo, G.J.; Stiff, A.; Mace, T.; Lesinski, G.B.; Benson, D.M., Jr.; Efebera, Y.A.; Rosko, A.E.; Pichiorri, F.; Grever, M.R.; et al. A phase i trial of single-agent reolysin in patients with relapsed multiple myeloma. Clin. Cancer Res. 2014, 20, 5946–5955. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, P.; Roldan, G.; George, D.; Wallace, C.; Palmer, C.A.; Morris, D.; Cairncross, G.; Matthews, M.V.; Markert, J.; Gillespie, Y.; et al. A phase i trial of intratumoral administration of reovirus in patients with histologically confirmed recurrent malignant gliomas. Mol. Ther. 2008, 16, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Markovic, S.N.; Suman, V.J.; Nuovo, G.J.; Vile, R.G.; Kottke, T.J.; Nevala, W.K.; Thompson, M.A.; Lewis, J.E.; Rumilla, K.M.; et al. Phase ii trial of intravenous administration of reolysin((r)) (reovirus serotype-3-dearing strain) in patients with metastatic melanoma. Mol. Ther. 2012, 20, 1998–2003. [Google Scholar] [CrossRef] [PubMed]
- White, C.L.; Twigger, K.R.; Vidal, L.; De Bono, J.S.; Coffey, M.; Heinemann, L.; Morgan, R.; Merrick, A.; Errington, F.; Vile, R.G.; et al. Characterization of the adaptive and innate immune response to intravenous oncolytic reovirus (dearing type 3) during a phase i clinical trial. Gene Ther. 2008, 15, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Errington, F.; Steele, L.; Prestwich, R.; Harrington, K.J.; Pandha, H.S.; Vidal, L.; de Bono, J.; Selby, P.; Coffey, M.; Vile, R.; et al. Reovirus activates human dendritic cells to promote innate antitumor immunity. J. Immunol. 2008, 180, 6018–6026. [Google Scholar] [CrossRef] [PubMed]
- Prestwich, R.J.; Errington, F.; Ilett, E.J.; Morgan, R.S.; Scott, K.J.; Kottke, T.; Thompson, J.; Morrison, E.E.; Harrington, K.J.; Pandha, H.S.; et al. Tumor infection by oncolytic reovirus primes adaptive antitumor immunity. Clin. Cancer Res. 2008, 14, 7358–7366. [Google Scholar] [CrossRef] [PubMed]
- Gujar, S.A.; Pan, D.A.; Marcato, P.; Garant, K.A.; Lee, P.W. Oncolytic virus-initiated protective immunity against prostate cancer. Mol. Ther. 2011, 19, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Gomez, G.; Ramirez, C.D.; Rivera, J.; Patel, M.; Norozian, F.; Wright, H.V.; Kashyap, M.V.; Barnstein, B.O.; Fischer-Stenger, K.; Schwartz, L.B.; et al. Tgf-beta 1 inhibits mast cell fc epsilon ri expression. J. Immunol. 2005, 174, 5987–5993. [Google Scholar] [CrossRef] [PubMed]
- Ndaw, V.S.; Abebayehu, D.; Spence, A.J.; Paez, P.A.; Kolawole, E.M.; Taruselli, M.T.; Caslin, H.L.; Chumanevich, A.P.; Paranjape, A.; Baker, B.; et al. Tgf-beta1 suppresses il-33-induced mast cell function. J. Immunol. 2017, 199, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Zaiatz-Bittencourt, V.; Finlay, D.K.; Gardiner, C.M. Canonical tgf-beta signaling pathway represses human nk cell metabolism. J. Immunol. 2018, 200, 3934–3941. [Google Scholar] [CrossRef] [PubMed]
- Taipale, J.; Lohi, J.; Saarinen, J.; Kovanen, P.T.; Keski-Oja, J. Human mast cell chymase and leukocyte elastase release latent transforming growth factor-beta 1 from the extracellular matrix of cultured human epithelial and endothelial cells. J. Biol. Chem. 1995, 270, 4689–4696. [Google Scholar] [CrossRef] [PubMed]
- Lindstedt, K.A.; Wang, Y.; Shiota, N.; Saarinen, J.; Hyytiainen, M.; Kokkonen, J.O.; Keski-Oja, J.; Kovanen, P.T. Activation of paracrine tgf-beta1 signaling upon stimulation and degranulation of rat serosal mast cells: A novel function for chymase. FASEB J. 2001, 15, 1377–1388. [Google Scholar] [CrossRef] [PubMed]
- Hutzen, B.; Chen, C.Y.; Wang, P.Y.; Sprague, L.; Swain, H.M.; Love, J.; Conner, J.; Boon, L.; Cripe, T.P. Tgf-beta inhibition improves oncolytic herpes viroimmunotherapy in murine models of rhabdomyosarcoma. Mol. Ther. Oncolytics 2017, 7, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Esaki, S.; Nigim, F.; Moon, E.; Luk, S.; Kiyokawa, J.; Curry, W., Jr.; Cahill, D.P.; Chi, A.S.; Iafrate, A.J.; Martuza, R.L.; et al. Blockade of transforming growth factor-beta signaling enhances oncolytic herpes simplex virus efficacy in patient-derived recurrent glioblastoma models. Int. J. Cancer 2017, 141, 2348–2358. [Google Scholar] [CrossRef] [PubMed]
- Geevarghese, S.K.; Geller, D.A.; de Haan, H.A.; Horer, M.; Knoll, A.E.; Mescheder, A.; Nemunaitis, J.; Reid, T.R.; Sze, D.Y.; Tanabe, K.K.; et al. Phase i/ii study of oncolytic herpes simplex virus nv1020 in patients with extensively pretreated refractory colorectal cancer metastatic to the liver. Hum. Gene. Ther. 2010, 21, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Cary, Z.D.; Willingham, M.C.; Lyles, D.S. Oncolytic vesicular stomatitis virus induces apoptosis in u87 glioblastoma cells by a type ii death receptor mechanism and induces cell death and tumor clearance in vivo. J. Virol. 2011, 85, 5708–5717. [Google Scholar] [CrossRef] [PubMed]
- Silberhumer, G.R.; Brader, P.; Wong, J.; Serganova, I.S.; Gonen, M.; Gonzalez, S.J.; Blasberg, R.; Zamarin, D.; Fong, Y. Genetically engineered oncolytic newcastle disease virus effectively induces sustained remission of malignant pleural mesothelioma. Mol. Cancer Ther. 2010, 9, 2761–2769. [Google Scholar] [CrossRef]
- Heo, J.; Reid, T.; Ruo, L.; Breitbach, C.J.; Rose, S.; Bloomston, M.; Cho, M.; Lim, H.Y.; Chung, H.C.; Kim, C.W.; et al. Randomized dose-finding clinical trial of oncolytic immunotherapeutic vaccinia jx-594 in liver cancer. Nat Med 2013, 19, 329–336. [Google Scholar] [CrossRef]
- Aoki, R.; Kawamura, T.; Goshima, F.; Ogawa, Y.; Nakae, S.; Nakao, A.; Moriishi, K.; Nishiyama, Y.; Shimada, S. Mast cells play a key role in host defense against herpes simplex virus infection through tnf-alpha and il-6 production. J. Invest. Dermatol. 2013, 133, 2170–2179. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Wang, D.; She, R.; Li, W.; Liu, S.; Han, D.; Wang, Y.; Ding, Y. Increased mast cell density during the infection with velogenic newcastle disease virus in chickens. Avian Pathol. 2008, 37, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Ushio, H.; Kawasaki, J.; Niyonsaba, F.; Takeuchi, M.; Baba, T.; Hiramatsu, K.; Okumura, K.; Ogawa, H. Expression and functional characterization of retinoic acid-inducible gene-i-like receptors of mast cells in response to viral infection. J. Innate. Immun. 2013, 5, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lai, Y.; Bernard, J.J.; Macleod, D.T.; Cogen, A.L.; Moss, B.; Di Nardo, A. Skin mast cells protect mice against vaccinia virus by triggering mast cell receptor s1pr2 and releasing antimicrobial peptides. J. Immunol. 2012, 188, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.R.; Bastin, D.; Chen, L.; Nguyen, A.; Storbeck, C.J.; Lefebvre, C.; Stojdl, D.; Bramson, J.L.; Bell, J.C.; Wan, Y. Type i ifn blockade uncouples immunotherapy-induced antitumor immunity and autoimmune toxicity. J. Clin. Invest. 2019, 129, 518–530. [Google Scholar] [CrossRef] [PubMed]
- Jawdat, D.M.; Rowden, G.; Marshall, J.S. Mast cells have a pivotal role in tnf-independent lymph node hypertrophy and the mobilization of langerhans cells in response to bacterial peptidoglycan. J. Immunol. 2006, 177, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Helby, J.; Bojesen, S.E.; Nielsen, S.F.; Nordestgaard, B.G. Ige and risk of cancer in 37 747 individuals from the general population. Ann. Oncol. 2015, 26, 1784–1790. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Jarolim, E.; Achatz, G.; Turner, M.C.; Karagiannis, S.; Legrand, F.; Capron, M.; Penichet, M.L.; Rodriguez, J.A.; Siccardi, A.G.; Vangelista, L.; et al. Allergooncology: The role of ige-mediated allergy in cancer. Allergy 2008, 63, 1255–1266. [Google Scholar] [CrossRef]
- Nigro, E.A.; Brini, A.T.; Yenagi, V.A.; Ferreira, L.M.; Achatz-Straussberger, G.; Ambrosi, A.; Sanvito, F.; Soprana, E.; van Anken, E.; Achatz, G.; et al. Cutting edge: Ige plays an active role in tumor immunosurveillance in mice. J. Immunol. 2016, 197, 2583–2588. [Google Scholar] [CrossRef]
- Neuchrist, C.; Kornfehl, J.; Grasl, M.; Lassmann, H.; Kraft, D.; Ehrenberger, K.; Scheiner, O. Distribution of immunoglobulins in squamous cell carcinoma of the head and neck. Int. Arch. Allergy Immunol. 1994, 104, 97–100. [Google Scholar] [CrossRef]
- Fu, S.L.; Pierre, J.; Smith-Norowitz, T.A.; Hagler, M.; Bowne, W.; Pincus, M.R.; Mueller, C.M.; Zenilman, M.E.; Bluth, M.H. Immunoglobulin e antibodies from pancreatic cancer patients mediate antibody-dependent cell-mediated cytotoxicity against pancreatic cancer cells. Clin. Exp. Immunol. 2008, 153, 401–409. [Google Scholar] [CrossRef]
- Karagiannis, S.N.; Bracher, M.G.; Beavil, R.L.; Beavil, A.J.; Hunt, J.; McCloskey, N.; Thompson, R.G.; East, N.; Burke, F.; Sutton, B.J.; et al. Role of ige receptors in ige antibody-dependent cytotoxicity and phagocytosis of ovarian tumor cells by human monocytic cells. Cancer Immunol. Immunother. 2008, 57, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Mommert, S.; Dittrich-Breiholz, O.; Stark, H.; Gutzmer, R.; Werfel, T. The histamine h4 receptor regulates chemokine production in human natural killer cells. Int. Arch. Allergy Immunol. 2015, 166, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Damaj, B.B.; Becerra, C.B.; Esber, H.J.; Wen, Y.; Maghazachi, A.A. Functional expression of h4 histamine receptor in human natural killer cells, monocytes, and dendritic cells. J. Immunol. 2007, 179, 7907–7915. [Google Scholar] [CrossRef] [PubMed]
- Aydin, E.; Johansson, J.; Nazir, F.H.; Hellstrand, K.; Martner, A. Role of nox2-derived reactive oxygen species in nk cell-mediated control of murine melanoma metastasis. Cancer Immunol. Res. 2017, 5, 804–811. [Google Scholar] [CrossRef]
- Hansson, M.; Hermodsson, S.; Brune, M.; Mellqvist, U.H.; Naredi, P.; Betten, A.; Gehlsen, K.R.; Hellstrand, K. Histamine protects t cells and natural killer cells against oxidative stress. J. Interferon Cytokine Res. 1999, 19, 1135–1144. [Google Scholar] [CrossRef] [PubMed]
- Betten, A.; Dahlgren, C.; Hermodsson, S.; Hellstrand, K. Histamine inhibits neutrophil nadph oxidase activity triggered by the lipoxin a4 receptor-specific peptide agonist trp-lys-tyr-met-val-met. Scand. J. Immunol. 2003, 58, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.; Henriksson, R.; Bergenheim, A.T.; Koskinen, L.O. Interleukin-2 and histamine in combination inhibit tumour growth and angiogenesis in malignant glioma. Br. J. Cancer 2000, 83, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Cuapio, A.; Post, M.; Cerny-Reiterer, S.; Gleixner, K.V.; Stefanzl, G.; Basilio, J.; Herndlhofer, S.; Sperr, W.R.; Brons, N.H.; Casanova, E.; et al. Maintenance therapy with histamine plus il-2 induces a striking expansion of two cd56bright nk cell subpopulations in patients with acute myeloid leukemia and supports their activation. Oncotarget 2016, 7, 46466–46481. [Google Scholar] [CrossRef] [PubMed]
- Brune, M.; Castaigne, S.; Catalano, J.; Gehlsen, K.; Ho, A.D.; Hofmann, W.K.; Hogge, D.E.; Nilsson, B.; Or, R.; Romero, A.I.; et al. Improved leukemia-free survival after postconsolidation immunotherapy with histamine dihydrochloride and interleukin-2 in acute myeloid leukemia: Results of a randomized phase 3 trial. Blood 2006, 108, 88–96. [Google Scholar] [CrossRef] [PubMed]
- McAlpine, S.M.; Issekutz, T.B.; Marshall, J.S. Virus stimulation of human mast cells results in the recruitment of cd56(+) t cells by a mechanism dependent on ccr5 ligands. FASEB J. 2012, 26, 1280–1289. [Google Scholar] [CrossRef] [PubMed]
- Becker, Y. Respiratory syncytial virus (rsv) evades the human adaptive immune system by skewing the th1/th2 cytokine balance toward increased levels of th2 cytokines and ige, markers of allergy--a review. Virus Genes 2006, 33, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Sigurs, N.; Bjarnason, R.; Sigurbergsson, F.; Kjellman, B. Respiratory syncytial virus bronchiolitis in infancy is an important risk factor for asthma and allergy at age 7. Am. J. Respir. Crit. Care Med. 2000, 161, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.J.; Gangnon, R.E.; Evans, M.D.; Roberg, K.A.; Anderson, E.L.; Pappas, T.E.; Printz, M.C.; Lee, W.M.; Shult, P.A.; Reisdorf, E.; et al. Wheezing rhinovirus illnesses in early life predict asthma development in high-risk children. Am. J. Respir. Crit. Care Med. 2008, 178, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Dakhama, A.; Lee, Y.M.; Ohnishi, H.; Jing, X.; Balhorn, A.; Takeda, K.; Gelfand, E.W. Virus-specific ige enhances airway responsiveness on reinfection with respiratory syncytial virus in newborn mice. J. Allergy Clin. Immunol. 2009, 123, 138–145.e5. [Google Scholar] [CrossRef] [PubMed]
- Kimman, T.G.; Terpstra, G.K.; Daha, M.R.; Westenbrink, F. Pathogenesis of naturally acquired bovine respiratory syncytial virus infection in calves: Evidence for the involvement of complement and mast cell mediators. Am. J. Vet. Res. 1989, 50, 694–700. [Google Scholar] [PubMed]
- Everard, M.L.; Fox, G.; Walls, A.F.; Quint, D.; Fifield, R.; Walters, C.; Swarbrick, A.; Milner, A.D. Tryptase and ige concentrations in the respiratory tract of infants with acute bronchiolitis. Arch. Dis. Child 1995, 72, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Oymar, K.; Halvorsen, T.; Aksnes, L. Mast cell activation and leukotriene secretion in wheezing infants. Relation to respiratory syncytial virus and outcome. Pediatr. Allergy Immunol. 2006, 17, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Al-Afif, A.; Alyazidi, R.; Oldford, S.A.; Huang, Y.Y.; King, C.A.; Marr, N.; Haidl, I.D.; Anderson, R.; Marshall, J.S. Respiratory syncytial virus infection of primary human mast cells induces the selective production of type i interferons, cxcl10, and ccl4. J. Allergy Clin. Immunol. 2015, 136, 1346–1354.e1. [Google Scholar] [CrossRef] [PubMed]
- Peritt, D.; Robertson, S.; Gri, G.; Showe, L.; Aste-Amezaga, M.; Trinchieri, G. Differentiation of human nk cells into nk1 and nk2 subsets. J. Immunol. 1998, 161, 5821–5824. [Google Scholar] [PubMed]
- Kaiko, G.E.; Phipps, S.; Angkasekwinai, P.; Dong, C.; Foster, P.S. Nk cell deficiency predisposes to viral-induced th2-type allergic inflammation via epithelial-derived il-25. J. Immunol. 2010, 185, 4681–4690. [Google Scholar] [CrossRef] [PubMed]
- Ple, C.; Barrier, M.; Amniai, L.; Marquillies, P.; Bertout, J.; Tsicopoulos, A.; Walzer, T.; Lassalle, P.; Duez, C. Natural killer cells accumulate in lung-draining lymph nodes and regulate airway eosinophilia in a murine model of asthma. Scand. J. Immunol. 2010, 72, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Wingett, D.; Nielson, C.P. Divergence in nk cell and cyclic amp regulation of t cell cd40l expression in asthmatic subjects. J. Leukoc. Biol. 2003, 74, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Mathias, C.B.; Guernsey, L.A.; Zammit, D.; Brammer, C.; Wu, C.A.; Thrall, R.S.; Aguila, H.L. Pro-inflammatory role of natural killer cells in the development of allergic airway disease. Clin. Exp. Allergy 2014, 44, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Zhang, J.; Xiao, W.; Feng, J.; Sun, R.; Tian, Z. Involvement of human natural killer cells in asthma pathogenesis: Natural killer 2 cells in type 2 cytokine predominance. J. Allergy Clin. Immunol. 2005, 115, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Aktas, E.; Akdis, M.; Bilgic, S.; Disch, R.; Falk, C.S.; Blaser, K.; Akdis, C.; Deniz, G. Different natural killer (nk) receptor expression and immunoglobulin e (ige) regulation by nk1 and nk2 cells. Clin. Exp. Immunol. 2005, 140, 301–309. [Google Scholar] [CrossRef]
- Mesdaghi, M.; Vodjgani, M.; Salehi, E.; Hadjati, J.; Sarrafnejad, A.; Bidad, K.; Berjisian, F. Natural killer cells in allergic rhinitis patients and nonatopic controls. Int. Arch. Allergy Immunol. 2010, 153, 234–238. [Google Scholar] [CrossRef]
- Raundhal, M.; Morse, C.; Khare, A.; Oriss, T.B.; Milosevic, J.; Trudeau, J.; Huff, R.; Pilewski, J.; Holguin, F.; Kolls, J.; et al. High ifn-gamma and low slpi mark severe asthma in mice and humans. J. Clin. Invest. 2015, 125, 3037–3050. [Google Scholar] [CrossRef]
- Krishnan, V.; Diette, G.B.; Rand, C.S.; Bilderback, A.L.; Merriman, B.; Hansel, N.N.; Krishnan, J.A. Mortality in patients hospitalized for asthma exacerbations in the united states. Am. J. Respir. Crit. Care Med. 2006, 174, 633–638. [Google Scholar] [CrossRef]
- Busse, W.W.; Lemanske, R.F., Jr.; Gern, J.E. Role of viral respiratory infections in asthma and asthma exacerbations. Lancet 2010, 376, 826–834. [Google Scholar] [CrossRef]
- Edwards, M.R.; Regamey, N.; Vareille, M.; Kieninger, E.; Gupta, A.; Shoemark, A.; Saglani, S.; Sykes, A.; Macintyre, J.; Davies, J.; et al. Impaired innate interferon induction in severe therapy resistant atopic asthmatic children. Mucosal. Immunol. 2013, 6, 797–806. [Google Scholar] [CrossRef]
- Dunican, E.M.; Fahy, J.V. The role of type 2 inflammation in the pathogenesis of asthma exacerbations. Ann. Am. Thorac. Soc. 2015, 12 Suppl 2, S144–149. [Google Scholar]
- Tel, J.; Torensma, R.; Figdor, C.G.; de Vries, I.J. Il-4 and il-13 alter plasmacytoid dendritic cell responsiveness to cpg DNA and herpes simplex virus-1. J. Invest. Dermatol. 2011, 131, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Moriwaki, A.; Matsumoto, K.; Matsunaga, Y.; Fukuyama, S.; Matsumoto, T.; Kan-o, K.; Noda, N.; Asai, Y.; Nakanishi, Y.; Inoue, H. Il-13 suppresses double-stranded rna-induced ifn-lambda production in lung cells. Biochem. Biophys. Res. Commun. 2011, 404, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Contoli, M.; Ito, K.; Padovani, A.; Poletti, D.; Marku, B.; Edwards, M.R.; Stanciu, L.A.; Gnesini, G.; Pastore, A.; Spanevello, A.; et al. Th2 cytokines impair innate immune responses to rhinovirus in respiratory epithelial cells. Allergy 2015, 70, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Jordan, W.J.; Eskdale, J.; Srinivas, S.; Pekarek, V.; Kelner, D.; Rodia, M.; Gallagher, G. Human interferon lambda-1 (ifn-lambda1/il-29) modulates the th1/th2 response. Genes Immun. 2007, 8, 254–261. [Google Scholar] [CrossRef]
- Huber, J.P.; Ramos, H.J.; Gill, M.A.; Farrar, J.D. Cutting edge: Type i ifn reverses human th2 commitment and stability by suppressing gata3. J. Immunol. 2010, 185, 813–817. [Google Scholar] [CrossRef]
- Djukanovic, R.; Harrison, T.; Johnston, S.L.; Gabbay, F.; Wark, P.; Thomson, N.C.; Niven, R.; Singh, D.; Reddel, H.K.; Davies, D.E.; et al. The effect of inhaled ifn-beta on worsening of asthma symptoms caused by viral infections. A randomized trial. Am. J. Respir. Crit. Care Med. 2014, 190, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Cerboni, C.; Zingoni, A.; Cippitelli, M.; Piccoli, M.; Frati, L.; Santoni, A. Antigen-activated human t lymphocytes express cell-surface nkg2d ligands via an atm/atr-dependent mechanism and become susceptible to autologous nk- cell lysis. Blood 2007, 110, 606–615. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tumor | Role | Biological Action or Clinical Observation |
|---|---|---|
| Pancreatic adenocarcinoma [41] * Prostate [42] | Pro-tumorigenic | Angiogenesis via VEGF production |
| Colorectal cancer [43] Lung adenocarcinoma [44] Melanoma [45] Gastric cancer [46] | Pro-tumorigenic | Angiogenesis |
| Renal carcinoma [47] | Pro-tumorigenic | Impair anti-tumoral responses via IL-10 and TGF-β production |
| Muscle invasive bladder cancer [48] | Pro-tumorigenic | Negative correlation between TIMC numbers and patient survival |
| Hepatocellular cancer [49] * Gastric cancer [50] | Pro-tumorigenic | IL-17 expression |
| * Hepatocarcinoma [51] * Colon cancer [52] | Pro-tumorigenic | Recruitment of MDSC Increase the suppressive role of MDSC via IFN-γ and NO production |
| * Skin carcinogenesis [53] | Anti-tumorigenic | Recruitment of effector immune cells to the tumor site |
| Breast cancer [54] | Anti-tumorigenic | Presence of mast cells is a good prognosis marker |
| Renal cell carcinoma [55] | Anti-tumorigenic | Positive correlation between TIMC numbers and patient survival |
| Esophageal squamous cell carcinoma [56] | Anti-tumorigenic | Negative correlation between IL-17+ mast cells and tumor invasion |
| Ovarian cancer [57] | Anti-tumorigenic | Mast cell infiltration in tumors with high vessel density was associated with improved survival |
| B cell lymphoma [58] Pleural mesothelioma [59] | Anti-tumorigenic | TIMC associated to favorable clinical outcome |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Portales-Cervantes, L.; Dawod, B.; Marshall, J.S. Mast Cells and Natural Killer Cells—A Potentially Critical Interaction. Viruses 2019, 11, 514. https://doi.org/10.3390/v11060514
Portales-Cervantes L, Dawod B, Marshall JS. Mast Cells and Natural Killer Cells—A Potentially Critical Interaction. Viruses. 2019; 11(6):514. https://doi.org/10.3390/v11060514
Chicago/Turabian StylePortales-Cervantes, Liliana, Bassel Dawod, and Jean S. Marshall. 2019. "Mast Cells and Natural Killer Cells—A Potentially Critical Interaction" Viruses 11, no. 6: 514. https://doi.org/10.3390/v11060514
APA StylePortales-Cervantes, L., Dawod, B., & Marshall, J. S. (2019). Mast Cells and Natural Killer Cells—A Potentially Critical Interaction. Viruses, 11(6), 514. https://doi.org/10.3390/v11060514