Virus-like Particle-Based L2 Vaccines against HPVs: Where Are We Today?

Abstract
:1. Background
- (i)
- Coat proteins that form the capsid are multivalently displayed to the immune system. Multivalent display enhances cross-linking of B-cell receptors on naïve B cells, leading to a stronger B-cell activation, proliferation, and differentiation, secretion of high-affinity antibodies, and the generation of long-lived memory B cells [6,7,8,9].
- (ii)
- VLPs have virally encoded T-helper cell epitopes, which enhance T-cell activation. Presentation of these epitopes by antigen-presenting cells (APCs) in association with major histocompatibility complex class II to T-helper cells leads to the activation of T-helper cells. In addition to this, co-stimulatory molecules from APCs help to activate the T-helper cells. Activated T-helper cells then secret cytokines, leading to the activation of B-cells, T-cells, and macrophages [6].
- (iii)
- Most VLPs are between 25 and 100 nm. This size range is very important for the following reasons. Firstly, nanoparticles between 10 and 200 nm, unlike those >200 nm, have been shown to efficiently enter the lymphatic system by direct diffusion through cell junctions. This allows VLPs to be efficiently exposed to immune cells. It is worth mentioning that VLPs can also be taken up and transported by APCs to the lymphatic system. Secondly, the small size of VLPs enables them to be transported to the lymphoid organs within a short period of time and to be efficiently taken up by APCs for presentation to T-helper cells [9].
- (iv)
- Some VLPs (especially bacteriophage VLPs) contain single-stranded (ss)RNA, which serves as endogenous adjuvant. Bacteriophage coat proteins (from MS2, PP7, etc.) have the potential to encapsudate ssRNA that codes for its capsid/coat protein during VLP assembly [10,11]. This ssRNA serves as endogenous adjuvant by directly activating immunostimulatory molecules such as toll-like receptors (TLR7 and TLR8), which in turn activate innate immune responses [9,12,13,14].
2. HPV Vaccines
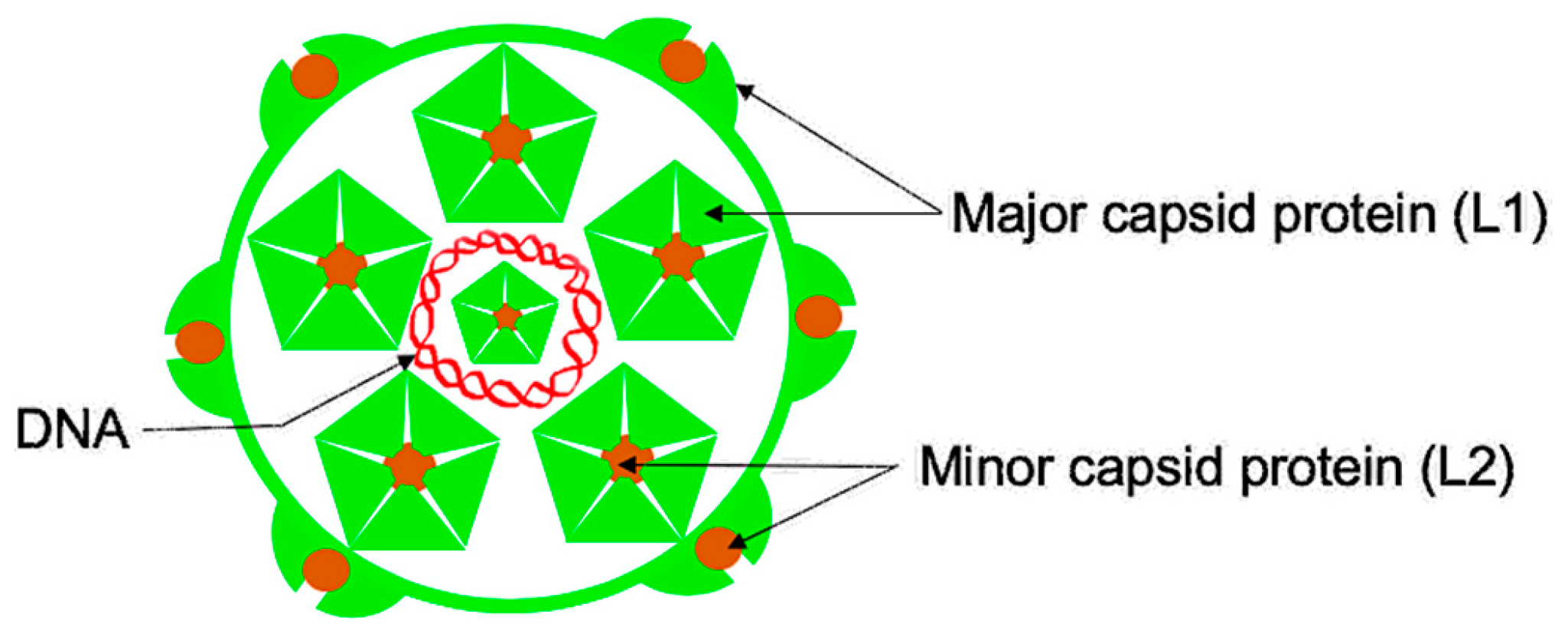
3. L2 Protein
4. Multivalent Display of HPV L2 on VLPs
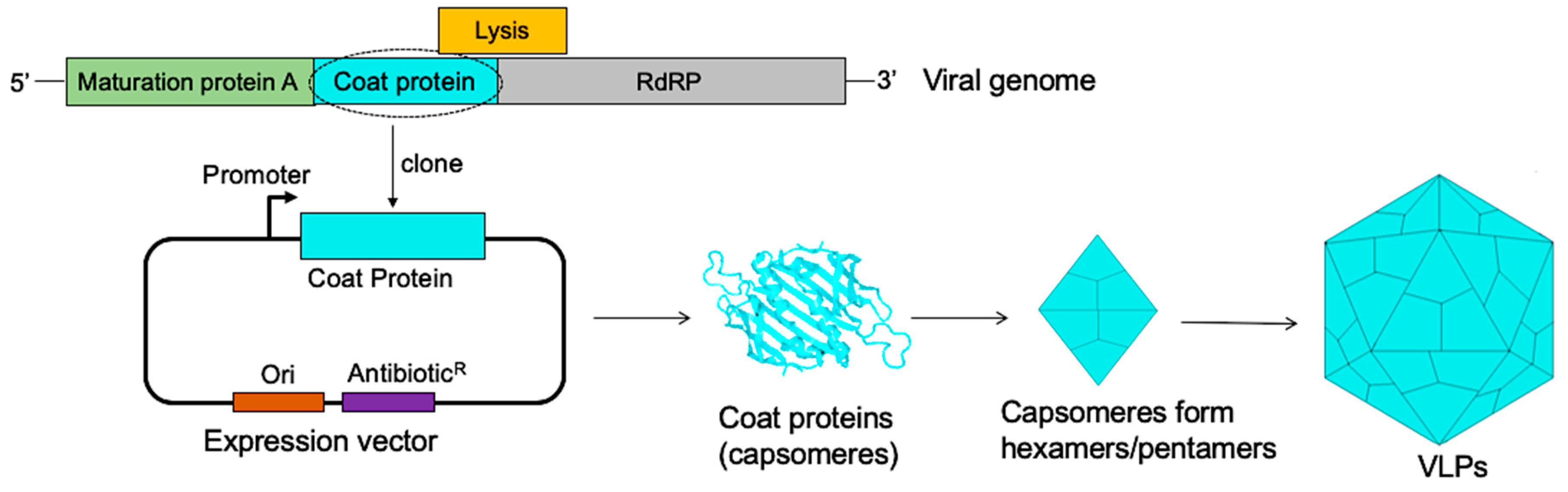
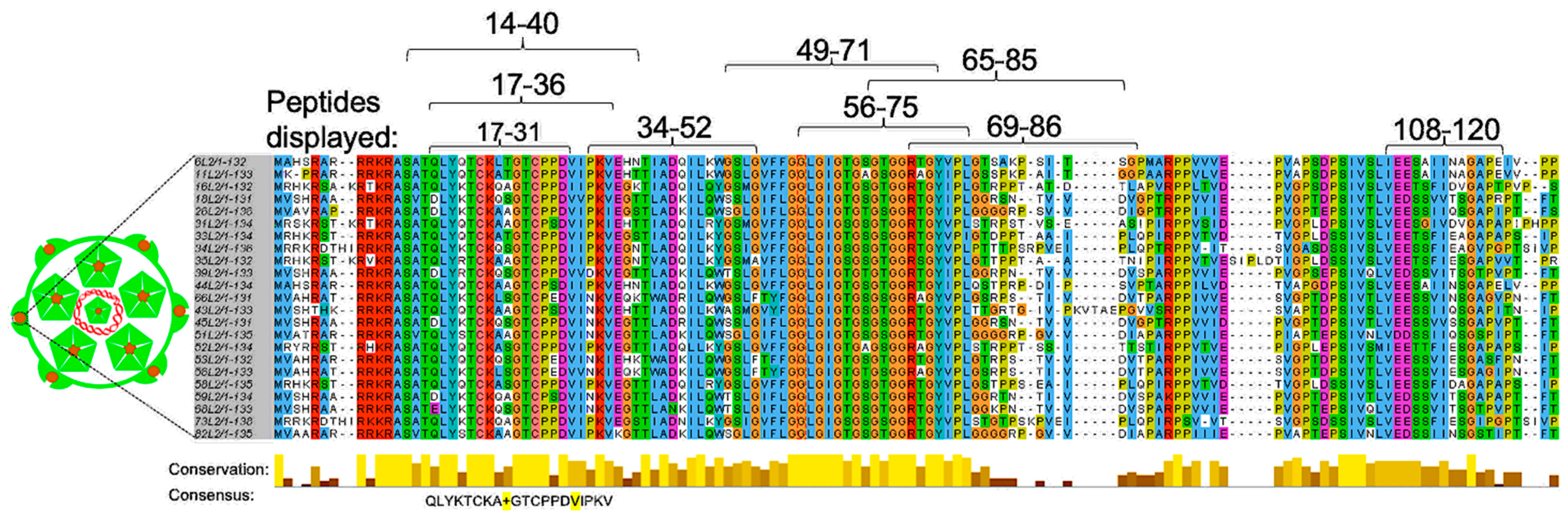
4.1. The Display of L2 Peptides on Bacteriophage VLP Platforms
- (i)
- (ii)
- (iii)
4.2. The Display of L2 Peptides on Eukaryotic VLP Platforms
5. Expert Commentary and Perspectives for the Future
Author Contributions
Funding
Conflicts of Interest
References
- Dai, S.; Wang, H.; Deng, F. Advances and challenges in enveloped virus-like particle (VLP)-based vaccines. J. Immunol. Sci. 2018, 2, 36–41. [Google Scholar]
- Chroboczek, J.; Szurgot, I.; Szolajska, E. Virus-like particles as vaccine. Acta Biochim. Pol. 2014, 61, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, B.; Uchida, M.; Douglas, T. Biomedical and Catalytic Opportunities of Virus-Like Particles in Nanotechnology. Adv. Virus Res. 2017, 97, 1–60. [Google Scholar] [PubMed] [Green Version]
- Zeltins, A. Construction and characterization of virus-like particles: A review. Mol. Biotechnol. 2013, 53, 92–107. [Google Scholar] [CrossRef]
- Tumban, E.; Peabody, J.; Tyler, M.; Peabody, D.S.; Chackerian, B. VLPs displaying a single L2 epitope induce broadly cross-neutralizing antibodies against human papillomavirus. PLoS ONE 2012, 7, e49751. [Google Scholar] [CrossRef] [Green Version]
- Chackerian, B.; Durfee, M.R.; Schiller, J.T. Virus-like display of a neo-self antigen reverses B cell anergy in a B cell receptor transgenic mouse model. J. Immunol. 2008, 180, 5816–5825. [Google Scholar] [CrossRef] [Green Version]
- Yuseff, M.I.; Pierobon, P.; Reversat, A.; Lennon-Dumenil, A.M. How B cells capture, process and present antigens: A crucial role for cell polarity. Nat. Rev. Immunol. 2013, 13, 475–486. [Google Scholar] [CrossRef]
- Zabel, F.; Kundig, T.M.; Bachmann, M.F. Virus-induced humoral immunity: On how B cell responses are initiated. Curr. Opin. Virol. 2013, 3, 357–362. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Peabody, D.S.; Manifold-Wheeler, B.; Medford, A.; Jordan, S.K.; do Carmo Caldeira, J.; Chackerian, B. Immunogenic display of diverse peptides on virus-like particles of RNA phage MS2. J. Mol. Biol. 2008, 380, 252–263. [Google Scholar] [CrossRef] [Green Version]
- Caldeira Jdo, C.; Medford, A.; Kines, R.C.; Lino, C.A.; Schiller, J.T.; Chackerian, B.; Peabody, D.S. Immunogenic display of diverse peptides, including a broadly cross-type neutralizing human papillomavirus L2 epitope, on virus-like particles of the RNA bacteriophage PP7. Vaccine 2010, 28, 4384–4393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibanez, L.I.; Roose, K.; De Filette, M.; Schotsaert, M.; De Sloovere, J.; Roels, S.; Pollard, C.; Schepens, B.; Grooten, J.; Fiers, W.; et al. M2e-displaying virus-like particles with associated RNA promote T helper 1 type adaptive immunity against influenza A. PLoS ONE 2013, 8, e59081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manolova, V.; Flace, A.; Bauer, M.; Schwarz, K.; Saudan, P.; Bachmann, M.F. Nanoparticles target distinct dendritic cell populations according to their size. Eur. J. Immunol. 2008, 38, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- Tumban, E.; Peabody, J.; Peabody, D.S.; Chackerian, B. A universal virus-like particle-based vaccine for human papillomavirus: Longevity of protection and role of endogenous and exogenous adjuvants. Vaccine 2013, 31, 4647–4654. [Google Scholar] [CrossRef] [Green Version]
- Gara, N.; Abdalla, A.; Rivera, E.; Zhao, X.; Werner, J.M.; Liang, T.J.; Hoofnagle, J.H.; Rehermann, B.; Ghany, M.G. Durability of antibody response against hepatitis B virus in healthcare workers vaccinated as adults. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2015, 60, 505–513. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Health and Human Services. Human Papillomavirus (HPV). Available online: https://www.hhs.gov/opa/reproductive-health/fact-sheets/sexually-transmitted-diseases/hpv/index.htm (accessed on 17 September 2019).
- Zhai, L.; Tumban, E. Gardasil-9: A global survey of projected efficacy. Antivir. Res. 2016, 130, 101–109. [Google Scholar] [CrossRef]
- Tumban, E. A Current Update on Human Papillomavirus-Associated Head and Neck Cancers. Viruses 2019, 11, 922. [Google Scholar] [CrossRef] [Green Version]
- Harper, D.M.; DeMars, L.R. HPV vaccines—A review of the first decade. Gynecol. Oncol. 2017, 146, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Haghshenas, M.R.; Mousavi, T.; Kheradmand, M.; Afshari, M.; Moosazadeh, M. Efficacy of Human Papillomavirus L1 Protein Vaccines (Cervarix and Gardasil) in Reducing the Risk of Cervical Intraepithelial Neoplasia: A Meta-analysis. Int. J. Prev. Med. 2017, 8, 44. [Google Scholar]
- Gilca, V.; Sauvageau, C.; Panicker, G.; De Serres, G.; Ouakki, M.; Unger, E.R. Antibody persistence after a single dose of quadrivalent HPV vaccine and the effect of a dose of nonavalent vaccine given 3–8 years later—An exploratory study. Hum. Vaccin. Immunother. 2018, 155, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Kirnbauer, R.; Booy, F.; Cheng, N.; Lowy, D.R.; Schiller, J.T. Papillomavirus L1 major capsid protein self-assembles into virus-like particles that are highly immunogenic. Proc. Natl. Acad. Sci. USA 1992, 89, 12180–12184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kines, R.C.; Thompson, C.D.; Lowy, D.R.; Schiller, J.T.; Day, P.M. The initial steps leading to papillomavirus infection occur on the basement membrane prior to cell surface binding. Proc. Natl. Acad. Sci. USA 2009, 106, 20458–20463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bywaters, S.M.; Brendle, S.A.; Tossi, K.P.; Biryukov, J.; Meyers, C.; Christensen, N.D. Antibody Competition Reveals Surface Location of HPV L2 Minor Capsid Protein Residues 17–36. Viruses 2017, 9, 336. [Google Scholar] [CrossRef] [Green Version]
- Buck, C.B.; Cheng, N.; Thompson, C.D.; Lowy, D.R.; Steven, A.C.; Schiller, J.T.; Trus, B.L. Arrangement of L2 within the papillomavirus capsid. J. Virol. 2008, 82, 5190–5197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpers, C.; Schirmacher, P.; Streeck, R.E.; Sapp, M. Assembly of the major and the minor capsid protein of human papillomavirus type 33 into virus-like particles and tubular structures in insect cells. Virology 1994, 200, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, S.C.; Patterson, N.A.; Ozbun, M.A.; Lambert, P.F. The minor capsid protein L2 contributes to two steps in the human papillomavirus type 31 life cycle. J. Virol. 2005, 79, 3938–3948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.W.; Roden, R.B. L2, the minor capsid protein of papillomavirus. Virology 2013, 445, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Kamper, N.; Day, P.M.; Nowak, T.; Selinka, H.C.; Florin, L.; Bolscher, J.; Hilbig, L.; Schiller, J.T.; Sapp, M. A membrane-destabilizing peptide in capsid protein L2 is required for egress of papillomavirus genomes from endosomes. J. Virol. 2006, 80, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Yutzy, W.H., 4th; Viscidi, R.P.; Roden, R.B. Interaction of L2 with beta-actin directs intracellular transport of papillomavirus and infection. J. Biol. Chem. 2003, 278, 12546–12553. [Google Scholar] [CrossRef] [Green Version]
- Campos, S.K. Subcellular Trafficking of the Papillomavirus Genome during Initial Infection: The Remarkable Abilities of Minor Capsid Protein L2. Viruses 2017, 9, 370. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.R.; Kjaer, S.K.; Sigurdsson, K.; Iversen, O.E.; Hernandez-Avila, M.; Wheeler, C.M.; Perez, G.; Koutsky, L.A.; Tay, E.H.; Garcia, P.; et al. The impact of quadrivalent human papillomavirus (HPV; types 6, 11, 16, and 18) L1 virus-like particle vaccine on infection and disease due to oncogenic nonvaccine HPV types in generally HPV-naive women aged 16-26 years. J. Infect. Dis. 2009, 199, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Joura, E.A.; Giuliano, A.R.; Iversen, O.E.; Bouchard, C.; Mao, C.; Mehlsen, J.; Moreira, E.D., Jr.; Ngan, Y.; Petersen, L.K.; Lazcano-Ponce, E.; et al. A 9-valent HPV vaccine against infection and intraepithelial neoplasia in women. N. Engl. J. Med. 2015, 372, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.F.; Brownlow, M.; Brown, M.; Kowalski, R.; Esser, M.T.; Ruiz, W.; Barr, E.; Brown, D.R.; Bryan, J.T. Antibodies from women immunized with Gardasil cross-neutralize HPV 45 pseudovirions. Hum. Vaccin. 2007, 3, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Toft, L.; Storgaard, M.; Muller, M.; Sehr, P.; Bonde, J.; Tolstrup, M.; Ostergaard, L.; Sogaard, O.S. Comparison of the immunogenicity and reactogenicity of Cervarix and Gardasil human papillomavirus vaccines in HIV-infected adults: A randomized, double-blind clinical trial. J. Infect. Dis. 2014, 209, 1165–1173. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, C.M.; Kjaer, S.K.; Sigurdsson, K.; Iversen, O.E.; Hernandez-Avila, M.; Perez, G.; Brown, D.R.; Koutsky, L.A.; Tay, E.H.; Garcia, P.; et al. The impact of quadrivalent human papillomavirus (HPV; types 6, 11, 16, and 18) L1 virus-like particle vaccine on infection and disease due to oncogenic nonvaccine HPV types in sexually active women aged 16-26 years. J. Infect. Dis. 2009, 199, 936–944. [Google Scholar] [CrossRef]
- Alphs, H.H.; Gambhira, R.; Karanam, B.; Roberts, J.N.; Jagu, S.; Schiller, J.T.; Zeng, W.; Jackson, D.C.; Roden, R.B. Protection against heterologous human papillomavirus challenge by a synthetic lipopeptide vaccine containing a broadly cross-neutralizing epitope of L2. Proc. Natl. Acad. Sci. USA 2008, 105, 5850–5855. [Google Scholar] [CrossRef] [Green Version]
- Christensen, N.D.; Kreider, J.W.; Kan, N.C.; DiAngelo, S.L. The open reading frame L2 of cottontail rabbit papillomavirus contains antibody-inducing neutralizing epitopes. Virology 1991, 181, 572–579. [Google Scholar] [CrossRef]
- Gambhira, R.; Jagu, S.; Karanam, B.; Gravitt, P.E.; Culp, T.D.; Christensen, N.D.; Roden, R.B. Protection of rabbits against challenge with rabbit papillomaviruses by immunization with the N terminus of human papillomavirus type 16 minor capsid antigen L2. J. Virol. 2007, 81, 11585–11592. [Google Scholar] [CrossRef] [Green Version]
- Pastrana, D.V.; Gambhira, R.; Buck, C.B.; Pang, Y.Y.; Thompson, C.D.; Culp, T.D.; Christensen, N.D.; Lowy, D.R.; Schiller, J.T.; Roden, R.B. Cross-neutralization of cutaneous and mucosal Papillomavirus types with anti-sera to the amino terminus of L2. Virology 2005, 337, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Roden, R.B.; Yutzy, W.H., 4th; Fallon, R.; Inglis, S.; Lowy, D.R.; Schiller, J.T. Minor capsid protein of human genital papillomaviruses contains subdominant, cross-neutralizing epitopes. Virology 2000, 270, 254–257. [Google Scholar] [CrossRef] [Green Version]
- Slingluff, C.L., Jr. The present and future of peptide vaccines for cancer: Single or multiple, long or short, alone or in combination? Cancer J. 2011, 17, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Joshi, M.D.; Singhania, S.; Ramsey, K.H.; Murthy, A.K. Peptide Vaccine: Progress and Challenges. Vaccines 2014, 2, 515–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, I.; Bolchi, A.; Moretto, N.; Canali, E.; Gissmann, L.; Tommasino, M.; Muller, M.; Ottonello, S. Potent anti-HPV immune responses induced by tandem repeats of the HPV16 L2 (20–38) peptide displayed on bacterial thioredoxin. Vaccine 2009, 27, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Seitz, H.; Canali, E.; Ribeiro-Muller, L.; Palfi, A.; Bolchi, A.; Tommasino, M.; Ottonello, S.; Muller, M. A three component mix of thioredoxin-L2 antigens elicits broadly neutralizing responses against oncogenic human papillomaviruses. Vaccine 2014, 32, 2610–2617. [Google Scholar] [CrossRef] [PubMed]
- Kalnin, K.; Chivukula, S.; Tibbitts, T.; Yan, Y.; Stegalkina, S.; Shen, L.; Cieszynski, J.; Costa, V.; Sabharwal, R.; Anderson, S.F.; et al. Incorporation of RG1 epitope concatemers into a self-adjuvanting Flagellin-L2 vaccine broaden durable protection against cutaneous challenge with diverse human papillomavirus genotypes. Vaccine 2017, 35, 4942–4951. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, X.; Liu, H.; Bao, Q.; Wang, Z.; Liao, G.; Xu, X. A rationally designed flagellin-L2 fusion protein induced serum and mucosal neutralizing antibodies against multiple HPV types. Vaccine 2019, 37, 4022–4030. [Google Scholar] [CrossRef] [PubMed]
- Jagu, S.; Karanam, B.; Gambhira, R.; Chivukula, S.V.; Chaganti, R.J.; Lowy, D.R.; Schiller, J.T.; Roden, R.B. Concatenated multitype L2 fusion proteins as candidate prophylactic pan-human papillomavirus vaccines. J. Natl. Cancer Inst. 2009, 101, 782–792. [Google Scholar] [CrossRef]
- Jagu, S.; Kwak, K.; Garcea, R.L.; Roden, R.B. Vaccination with multimeric L2 fusion protein and L1 VLP or capsomeres to broaden protection against HPV infection. Vaccine 2010, 28, 4478–4486. [Google Scholar] [CrossRef] [Green Version]
- Pouyanfard, S.; Spagnoli, G.; Bulli, L.; Balz, K.; Yang, F.; Odenwald, C.; Seitz, H.; Mariz, F.C.; Bolchi, A.; Ottonello, S.; et al. Minor Capsid Protein L2 Polytope Induces Broad Protection against Oncogenic and Mucosal Human Papillomaviruses. J. Virol. 2018, 92, e01930-17. [Google Scholar] [CrossRef] [Green Version]
- Seitz, H.; Ribeiro-Muller, L.; Canali, E.; Bolchi, A.; Tommasino, M.; Ottonello, S.; Muller, M. Robust In Vitro and In Vivo Neutralization against Multiple High-Risk HPV Types Induced by a Thermostable Thioredoxin-L2 Vaccine. Cancer Prev. Res. (Phila.) 2015, 8, 932–941. [Google Scholar] [CrossRef] [Green Version]
- Spagnoli, G.; Pouyanfard, S.; Cavazzini, D.; Canali, E.; Maggi, S.; Tommasino, M.; Bolchi, A.; Muller, M.; Ottonello, S. Broadly neutralizing antiviral responses induced by a single-molecule HPV vaccine based on thermostable thioredoxin-L2 multiepitope nanoparticles. Sci. Rep. 2017, 7, 18000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, V.; Arora, U.; Shukla, R.; Poddar, A.; Shanmugam, R.K.; White, L.J.; Mattocks, M.M.; Raut, R.; Perween, A.; Tyagi, P.; et al. A tetravalent virus-like particle vaccine designed to display domain III of dengue envelope proteins induces multi-serotype neutralizing antibodies in mice and macaques which confer protection against antibody dependent enhancement in AG129 mice. PLoS Negl. Trop. Dis. 2018, 12, e0006191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daly, S.M.; Joyner, J.A.; Triplett, K.D.; Elmore, B.O.; Pokhrel, S.; Frietze, K.M.; Peabody, D.S.; Chackerian, B.; Hall, P.R. VLP-based vaccine induces immune control of Staphylococcus aureus virulence regulation. Sci. Rep. 2017, 7, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crossey, E.; Amar, M.J.A.; Sampson, M.; Peabody, J.; Schiller, J.T.; Chackerian, B.; Remaley, A.T. A cholesterol-lowering VLP vaccine that targets PCSK9. Vaccine 2015, 33, 5747–5755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, H.K.; Tan, W.S.; Ho, K.L. Virus like particles as a platform for cancer vaccine development. PeerJ 2017, 5, e4053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, L.; Peabody, J.; Pang, Y.S.; Schiller, J.; Chackerian, B.; Tumban, E. A novel candidate HPV vaccine: MS2 phage VLP displaying a tandem HPV L2 peptide offers similar protection in mice to Gardasil-9. Antivir. Res. 2017, 147, 116–123. [Google Scholar] [CrossRef]
- Tumban, E.; Peabody, J.; Peabody, D.S.; Chackerian, B. A pan-HPV vaccine based on bacteriophage PP7 VLPs displaying broadly cross-neutralizing epitopes from the HPV minor capsid protein, L2. PLoS ONE 2011, 6, e23310. [Google Scholar] [CrossRef]
- Tyler, M.; Tumban, E.; Peabody, D.S.; Chackerian, B. The use of hybrid virus-like particles to enhance the immunogenicity of a broadly protective HPV vaccine. Biotechnol. Bioeng. 2014, 111, 2398–2406. [Google Scholar] [CrossRef] [Green Version]
- Janitzek, C.M.; Peabody, J.; Thrane, S.; Carlsen, P.H.; Theander, T.G.; Salanti, A.; Chackerian, B.; Nielsen, M.A.; Sander, A.F. A proof-of-concept study for the design of a VLP-based combinatorial HPV and placental malaria vaccine. Sci. Rep. 2019, 9, 5260. [Google Scholar] [CrossRef]
- Tyler, M.; Tumban, E.; Dziduszko, A.; Ozbun, M.A.; Peabody, D.S.; Chackerian, B. Immunization with a consensus epitope from human papillomavirus L2 induces antibodies that are broadly neutralizing. Vaccine 2014, 32, 4267–4274. [Google Scholar] [CrossRef] [Green Version]
- Zhai, L.; Yadav, R.; Kunda, N.K.; Anderson, D.; Bruckner, E.; Miller, E.K.; Basu, R.; Muttil, P.; Tumban, E. Oral immunization with bacteriophage MS2-L2 VLPs protects against oral and genital infection with multiple HPV types associated with head & neck cancers and cervical cancer. Antivir. Res. 2019, 166, 56–65. [Google Scholar] [PubMed]
- Kawana, K.; Kawana, Y.; Yoshikawa, H.; Taketani, Y.; Yoshiike, K.; Kanda, T. Nasal immunization of mice with peptide having a cross-neutralization epitope on minor capsid protein L2 of human papillomavirus type 16 elicit systemic and mucosal antibodies. Vaccine 2001, 19, 1496–1502. [Google Scholar] [CrossRef]
- Varsani, A.; Williamson, A.L.; de Villiers, D.; Becker, I.; Christensen, N.D.; Rybicki, E.P. Chimeric human papillomavirus type 16 (HPV-16) L1 particles presenting the common neutralizing epitope for the L2 minor capsid protein of HPV-6 and HPV-16. J. Virol. 2003, 77, 8386–8393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, K.; Ochi, H.; Matsumoto, T.; Yoshikawa, H.; Kanda, T. Modification of human papillomavirus-like particle vaccine by insertion of the cross-reactive L2-epitopes. J. Med. Virol. 2008, 80, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Slupetzky, K.; Gambhira, R.; Culp, T.D.; Shafti-Keramat, S.; Schellenbacher, C.; Christensen, N.D.; Roden, R.B.; Kirnbauer, R. A papillomavirus-like particle (VLP) vaccine displaying HPV16 L2 epitopes induces cross-neutralizing antibodies to HPV11. Vaccine 2007, 25, 2001–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schellenbacher, C.; Roden, R.; Kirnbauer, R. Chimeric L1-L2 virus-like particles as potential broad-spectrum human papillomavirus vaccines. J. Virol. 2009, 83, 10085–10095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schellenbacher, C.; Huber, B.; Skoll, M.; Shafti-Keramat, S.; Roden, R.B.S.; Kirnbauer, R. Incorporation of RG1 epitope into HPV16L1-VLP does not compromise L1-specific immunity. Vaccine 2019, 37, 3529–3534. [Google Scholar] [CrossRef] [PubMed]
- Schellenbacher, C.; Kwak, K.; Fink, D.; Shafti-Keramat, S.; Huber, B.; Jindra, C.; Faust, H.; Dillner, J.; Roden, R.B.S.; Kirnbauer, R. Efficacy of RG1-VLP vaccination against infections with genital and cutaneous human papillomaviruses. J. Investig. Dermatol. 2013, 133, 2706–2713. [Google Scholar] [CrossRef] [Green Version]
- Boxus, M.; Fochesato, M.; Miseur, A.; Mertens, E.; Dendouga, N.; Brendle, S.; Balogh, K.K.; Christensen, N.D.; Giannini, S.L. Broad Cross-Protection Is Induced in Preclinical Models by a Human Papillomavirus Vaccine Composed of L1/L2 Chimeric Virus-Like Particles. J. Virol. 2016, 90, 6314–6325. [Google Scholar] [CrossRef] [Green Version]
- Huber, B.; Schellenbacher, C.; Jindra, C.; Fink, D.; Shafti-Keramat, S.; Kirnbauer, R. A chimeric 18L1-45RG1 virus-like particle vaccine cross-protects against oncogenic alpha-7 human papillomavirus types. PLoS ONE 2015, 10, e0120152. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, T.; Liu, H.; Hao, Y.; Liao, G.; Xu, X. Displaying 31RG-1 peptide on the surface of HPV16 L1 by use of a human papillomavirus chimeric virus-like particle induces cross-neutralizing antibody responses in mice. Hum. Vaccin. Immunother. 2018, 14, 2025–2033. [Google Scholar] [CrossRef] [PubMed]
- Nieto, K.; Weghofer, M.; Sehr, P.; Ritter, M.; Sedlmeier, S.; Karanam, B.; Seitz, H.; Muller, M.; Kellner, M.; Horer, M.; et al. Development of AAVLP(HPV16/31L2) particles as broadly protective HPV vaccine candidate. PLoS ONE 2012, 7, e39741. [Google Scholar] [CrossRef] [PubMed]
- Jagu, S.; Karanam, B.; Wang, J.W.; Zayed, H.; Weghofer, M.; Brendle, S.A.; Balogh, K.K.; Tossi, K.P.; Roden, R.B.S.; Christensen, N.D. Durable immunity to oncogenic human papillomaviruses elicited by adjuvanted recombinant Adeno-associated virus-like particle immunogen displaying L2 17-36 epitopes. Vaccine 2015, 33, 5553–5563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.H.; Alkutkar, T.; Karanam, B.; Roden, R.B.; Ketner, G.; Ibeanu, O.A. Capsid display of a conserved human papillomavirus L2 peptide in the adenovirus 5 hexon protein: A candidate prophylactic hpv vaccine approach. Virol. J. 2015, 12, 140. [Google Scholar] [CrossRef] [Green Version]
- Vujadinovic, M.; Khan, S.; Oosterhuis, K.; Uil, T.G.; Wunderlich, K.; Damman, S.; Boedhoe, S.; Verwilligen, A.; Knibbe, J.; Serroyen, J.; et al. Adenovirus based HPV L2 vaccine induces broad cross-reactive humoral immune responses. Vaccine 2018, 36, 4462–4470. [Google Scholar] [CrossRef]
- Diamos, A.G.; Larios, D.; Brown, L.; Kilbourne, J.; Kim, H.S.; Saxena, D.; Palmer, K.E.; Mason, H.S. Vaccine synergy with virus-like particle and immune complex platforms for delivery of human papillomavirus L2 antigen. Vaccine 2019, 37, 137–144. [Google Scholar] [CrossRef]
- Cerovska, N.; Hoffmeisterova, H.; Pecenkova, T.; Moravec, T.; Synkova, H.; Plchova, H.; Veleminsky, J. Transient expression of HPV16 E7 peptide (aa 44–60) and HPV16 L2 peptide (aa 108–120) on chimeric potyvirus-like particles using Potato virus X-based vector. Protein Expr. Purif. 2008, 58, 154–161. [Google Scholar] [CrossRef]
- Plevka, P.; Tars, K.; Liljas, L. Structure and stability of icosahedral particles of a covalent coat protein dimer of bacteriophage MS2. Protein Sci. 2009, 18, 1653–1661. [Google Scholar] [CrossRef] [Green Version]
- Guan, J.; Bywaters, S.M.; Brendle, S.A.; Ashley, R.E.; Makhov, A.M.; Conway, J.F.; Christensen, N.D.; Hafenstein, S. Cryoelectron Microscopy Maps of Human Papillomavirus 16 Reveal L2 Densities and Heparin Binding Site. Structure 2017, 25, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Tumban, E.; Muttil, P.; Escobar, C.A.; Peabody, J.; Wafula, D.; Peabody, D.S.; Chackerian, B. Preclinical refinements of a broadly protective VLP-based HPV vaccine targeting the minor capsid protein, L2. Vaccine 2015, 33, 3346–3353. [Google Scholar] [CrossRef] [Green Version]
- Kunda, N.K.; Peabody, J.; Zhai, L.; Price, D.N.; Chackerian, B.; Tumban, E.; Muttil, P. Evaluation of the thermal stability and the protective efficacy of spray-dried HPV vaccine, Gardasil(R) 9. Hum. Vaccin. Immunother. 2019, 15, 1995–2002. [Google Scholar] [CrossRef] [PubMed]
- Caldeira Jdo, C.; Peabody, D.S. Thermal stability of RNA phage virus-like particles displaying foreign peptides. J. Nanobiotechnol. 2011, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zak, D.E.; Andersen-Nissen, E.; Peterson, E.R.; Sato, A.; Hamilton, M.K.; Borgerding, J.; Krishnamurty, A.T.; Chang, J.T.; Adams, D.J.; Hensley, T.R.; et al. Merck Ad5/HIV induces broad innate immune activation that predicts CD8(+) T-cell responses but is attenuated by preexisting Ad5 immunity. Proc. Natl. Acad. Sci. USA 2012, 109, E3503–E3512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knuchel, M.C.; Marty, R.R.; Morin, T.N.; Ilter, O.; Zuniga, A.; Naim, H.Y. Relevance of a pre-existing measles immunity prior immunization with a recombinant measles virus vector. Hum. Vaccin. Immunother. 2013, 9, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Zarnitsyna, V.I.; Lavine, J.; Ellebedy, A.; Ahmed, R.; Antia, R. Multi-epitope Models Explain How Pre-existing Antibodies Affect the Generation of Broadly Protective Responses to Influenza. PLoS Pathog. 2016, 12, e1005692. [Google Scholar] [CrossRef] [Green Version]
- Voysey, M.; Kelly, D.F.; Fanshawe, T.R.; Sadarangani, M.; O’Brien, K.L.; Perera, R.; Pollard, A.J. The Influence of Maternally Derived Antibody and Infant Age at Vaccination on Infant Vaccine Responses: An Individual Participant Meta-analysis. JAMA Pediatr. 2017, 171, 637–646. [Google Scholar] [CrossRef]
- Masat, E.; Pavani, G.; Mingozzi, F. Humoral immunity to AAV vectors in gene therapy: Challenges and potential solutions. Discov. Med. 2013, 15, 379–389. [Google Scholar]
- McCluskie, M.J.; Evans, D.M.; Zhang, N.; Benoit, M.; McElhiney, S.P.; Unnithan, M.; DeMarco, S.C.; Clay, B.; Huber, C.; Deora, A.; et al. The effect of preexisting anti-carrier immunity on subsequent responses to CRM197 or Qb-VLP conjugate vaccines. Immunopharmacol. Immunotoxicol. 2016, 38, 184–196. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of VLPs | HPV L2 Epitope Displayed (aa) | Length of Epitope | Position of Insertion on VLPs | Dose Immunized with | Neutralized or Protected against HPV Pseudoviruses (PsVs) * | Ref #. |
|---|---|---|---|---|---|---|
| MS2 | HPV16 L2 (aa 17–31) | 15 | N-terminus | Mice immunized with two doses of VLPs (5 μg/dose) | PsVs (genital): 5/6/16/33/35/39/45/51/53/58 | [5] |
| HPV16 L2 (aa 20–31) | 12 | ‘’ | ‘’ | Not tested | ||
| HPV16 L2 (aa 14–40) | 26 | ‘’ | ‘’ | ‘’ | ||
| HPV16 L2/31L2 (aa 20–31 + aa 17-31) | 27 | N-terminus | Mice immunized with two doses of a mixture of VLPs displaying 16L2/31L2 and VLPs displaying consensus epitope (10 μg/dose) | PsVs (genital): 16/18/31/33/45/58 | [57] | |
| Consensus (aa 69–86) | 18 | Mice orally immunized with three doses of a mixture of VLPs displaying 16/31L2 and VLPs displaying consensus epitope (100 μg/dose) | PsVs (genital): 11/16/53/56 | [62] | ||
| PsVs (oral): 16/35/39/52/58 | ||||||
| HPV16 L2 (aa 17–31) | 15 | 16L2 on N-terminus of one coat protein and 31L2 on the N-terminus of another coat protein | Mice immunized with two doses of hybrid VLPs (5 μg/dose) | PsVs (neutralized): 16/31/45/18/58 | [59] | |
| HPV31 L2 (aa 17–31) | 15 | |||||
| PP7 | HPVs: 1L2, 5L2, 6L2, 11L2, 16L2, 18L2, 45L2, 58L2 (aa 17–31) | 15 from each L2 on each VLP | AB-loop (internal insertion) | Mice immunized with three doses of a mixture of VLPs displaying all eight L2 peptides (10 μg/dose) | PsVs (genital): 6/16/18/31/45/52/58 PsVs (cutaneous): 5 | [58] |
| PP7 | HPV16 L2 (aa 17–31) | 15 | 16L2 on AB-loop of one coat protein and 18L2 on the AB-loop of another coat protein | Mice immunized with two doses of hybrid VLPs (5 μg/dose) | PsVs (genital): 6 | [59] |
| HPV18 L2 (aa 17–31) | 15 | |||||
| HPV16 L2 (aa 35–50) | 16 | Chemical conjugation on surface | Mice immunized with three doses each VLP (5 μg/dose) | PsVs (genital): 16 (no protection) | [61] | |
| HPV16 L2 (aa 51–65) | 15 | PsVs (genital): 16 | ||||
| Qβ | HPV16 L2 (aa 34–52) | 19 | Chemical conjugation on surface | Mice immunized with three doses of each VLP (5 μg/dose) | PsVs (genital): 16 (no protection) | [61] |
| HPV16 L2 (aa 49–71) | 23 | PsVs (genital): 16 | ||||
| Consensus (aa 65–85) | 18 | PsVs (genital): 16 | ||||
| HPV16 L2 (aa 108–120) | 13 | PsVs (genital): 6/16/31/45/58 | ||||
| AP205 | A concatemer of HPV: 16L2, 18L2, 31L2, 35L2, 52L2 (aa 17–38) | 110 | C-terminus | Mice immunized with two doses of VLPs (5 μg/dose) | PsVs (genital): 16, 31, 35, 45, 52 | [60] |
| Source of VLPs | HPV L2 Peptide Displayed (aa) | Length of Peptide | Position of Insertion on VLPs | Dose Immunized with | Neutralized or Protected against HPV Pseudoviruses (PsVs) * | Ref #. |
|---|---|---|---|---|---|---|
| HPV16 L1 | HPV16 L2 (aa 414–426) | 13 | Helix 4 loop (aa 414–426 replaced with L2) | Mice immunized with three doses of VLPs (100 μg/dose) | Assays not conducted; however, the peptide has been shown, in previous studies [63], to neutralize HPV 16 and HPV 6 | [64] |
| HPV16 L1 | HPV16 L2 (aa 18–38) | 21 | Helix 4 loop (between amino acids 430 and 433) | Rabbits immunized with five doses of VLPs (concentration not provided) | PsVs (neutralized): 16/18/31/52/58 | [65] |
| HPV16 L2 (aa 56–75) | 20 | PsVs (neutralized): 16/18/31/52/58 | ||||
| HPV16 L2 (aa 96–115) | 20 | PsVs (neutralized): 16/18/31/52/58 | ||||
| BPV1 | HPV16 L2 (aa 69–81) | 13 | Between amino acids 133 and 134 (DE loop) | Rabbits immunized with three doses of VLPs (50 μg/dose) | PsVs (neutralized): PsV11/PsV16 | [66] |
| HPV16 L2 (aa 108–120) | 13 | PsVs (neutralized): PsV11/PsV16 | ||||
| BPV1 | HPV16 L2 (aa 17–36) | 20 | Between amino acids 133 and 134 | Rabbits immunized with four doses of VLPs (25–50 μg/dose) | PsVs (neutralized): 5/16/18/45/58 | [67] |
| HPV16 L1 | HPV16 L2 (aa 17–36) | 20 | Between amino acids 136 and 137 (DE loop) | Rabbits immunized with four doses of VLPs (25–50 μg/dose) | PsVs (neutralized): 5/6/11/16/18/31/45/52/58/70 | [67,68] |
| Passive transferred sera protected mice against PsVs (genital): 6/16/18/26/31/33/34/35/39/43/44/45/51/52/53/56/58/59/66/68/73 | [69] | |||||
| Mice immunized with two doses of a mixture of 16L1 VLPs displaying 16L2 and HPV18 L1 VLPs (10 μg/dose) | PsV (genital): 58 | [68] | ||||
| HPV18 L1 | HPV33 L2 (aa 17–36) | 20 | Between amino acids 134 and 135 (DE loop) | Rabbits immunized with three doses of each VLP (20 μg/dose or 100 μg/dose) | QsV $ (neutralized): 5/6/11/18/16/31/33/45/52/58 QsVs (cutaneous): 11/18/58 | [70] |
| HPV58 L2 (aa 56–75) | 20 | C-terminus | QsV (neutralized): 31/45 | |||
| HPV18 L1 | HPV33 L2 (aa 17–36) | 20 | HPV33 L2 inserted on DE loop and HPV58 L2 inserted on C-terminus | Rabbits immunized with three doses of a mixture of 18L1 VLPs displaying 33L2 on DE loop and 58L2 on C- terminus mixed human dose of 16L1/18L1 VLPs (Cervarix vaccine). Two μg/dose of chimeric 18L1-L2 and 1/10th per dose of Cervarix was used | QsVs (cutaneous): 6/11/16/18/31/35/39/45/59 | [70] |
| HPV58 L2 (aa 56–75) | 20 | Mice immunized with two of a mixture of 18L1 VLPs displaying 33L2 on DE loop and 58L2 on C- terminus mixed human dose of 16L1/18L1 VLPs (Cervarix vaccine). Two μg/dose of chimeric 18L1-L2 and 1/10th per dose of Cervarix was used | PsVs (genital): 11/16/35/45/58/59 | |||
| HPV18 L1 | HPV45 L2 (aa 16–35) | 20 | Between amino acids 134 and 135 | Rabbits immunized with five doses of VLPs (50 μg/dose) | Passive transferred sera protected mice against PsVs (genital): 16/18/39/45/59/68 | [71] |
| HPV16 L1 | HPV31 L2 (aa 17–38) | 22 | Helix 4 loop (between amino acids 430 and 433) | Mice immunized with four doses of VLPs (10 μg/dose) | PsVs (neutralized): 2/5/6/11/16/18/27/31/33/35/39/45/52/57/58/59/68 | [72] |
| Adeno- associated virus 2 VP3 virus particles | HPV16 L2 (aa 17–36) | 20 | HPV16 L2 inserted at position 587 and HPV31 L2 inserted at position 453 of VP3 | Mice immunized with three doses of particles (1 × 1011 per dose) | PsVs (neutralized): 16/18/31/45/52/58 | [73] |
| HPV31 L2 (aa 17–36) | 20 | Rabbits immunized with four doses of particles (2 × 1012 per dose) | PsVs (neutralized): 16/18/31/45/52/58 Passive transferred sera protected mice against PsVs (genital): 16 | |||
| Adeno- associated virus 2 VP3 virus particles | HPV16 L2 (aa 17–36) | 20 | HPV16 L2 inserted at position 587 and HPV31 L2 inserted at position 453 of VP3 | Mice immunized with three doses of particles (1 × 1012 per dose) | PsVs (neutralized): 16/18/31/45/58 | [74] |
| HPV31 L2 (aa 17–36) | 20 | Rabbits immunized with three doses of particles (1 × 1019 per dose) | QsVs (cutaneous): 16/18/31/35/39/45/58/59 | |||
| Adenovirus type 5 | HPV16 L2 (12–41) | 30 | Hexon protein hypervariable regions 1 and 5 | Mice immunized with three doses of particles (1 × 1010 per dose) | PsVs (genital): 16/56 PsVs (neutralized): 16/73 | [75] |
| Adenovirus type 35 | Concatemers of HPV: 6L2, 11L2, 16L2, 18L2, 31L2, 33L2, 45L2, 52L2, 58L2 (aa 17–36) | 60, 80, 100 | C-terminus of pIX | Mice immunized with two doses of particles (1 × 1010 per dose) | PsVs (neutralized): 16/18/31/59 | [76] |
| Hepatitis B core (HBc) VLPs | HPV16 L2 (aa 14–122) | 108 | Inserted between a heterodimer of HBc | Mice immunized with three doses of VLPs (5μg/dose) | PsVs (neutralized): 16 | [77] |
| Potato virus A VLPs | HPV16 L2 (aa 108–120) | 13 | L2 inserted on the N-terminus and E7 inserted on the C-terminus | Immunization studies not conducted | No studies | [78] |
| HPV16 E7 (aa 44–60) | 17 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, R.; Zhai, L.; Tumban, E. Virus-like Particle-Based L2 Vaccines against HPVs: Where Are We Today? Viruses 2020, 12, 18. https://doi.org/10.3390/v12010018
Yadav R, Zhai L, Tumban E. Virus-like Particle-Based L2 Vaccines against HPVs: Where Are We Today? Viruses. 2020; 12(1):18. https://doi.org/10.3390/v12010018
Chicago/Turabian StyleYadav, Rashi, Lukai Zhai, and Ebenezer Tumban. 2020. "Virus-like Particle-Based L2 Vaccines against HPVs: Where Are We Today?" Viruses 12, no. 1: 18. https://doi.org/10.3390/v12010018
APA StyleYadav, R., Zhai, L., & Tumban, E. (2020). Virus-like Particle-Based L2 Vaccines against HPVs: Where Are We Today? Viruses, 12(1), 18. https://doi.org/10.3390/v12010018