Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Influenza Viruses
2.2. Serum Samples
2.3. Haemagglutination Inhibition Assay
2.4. Virus Neutralization Assay
2.5. Statistical Analysis
3. Results
3.1. Investigations on Potential Cross-Reactivity between IDV- and ICV-Positive Serum Samples
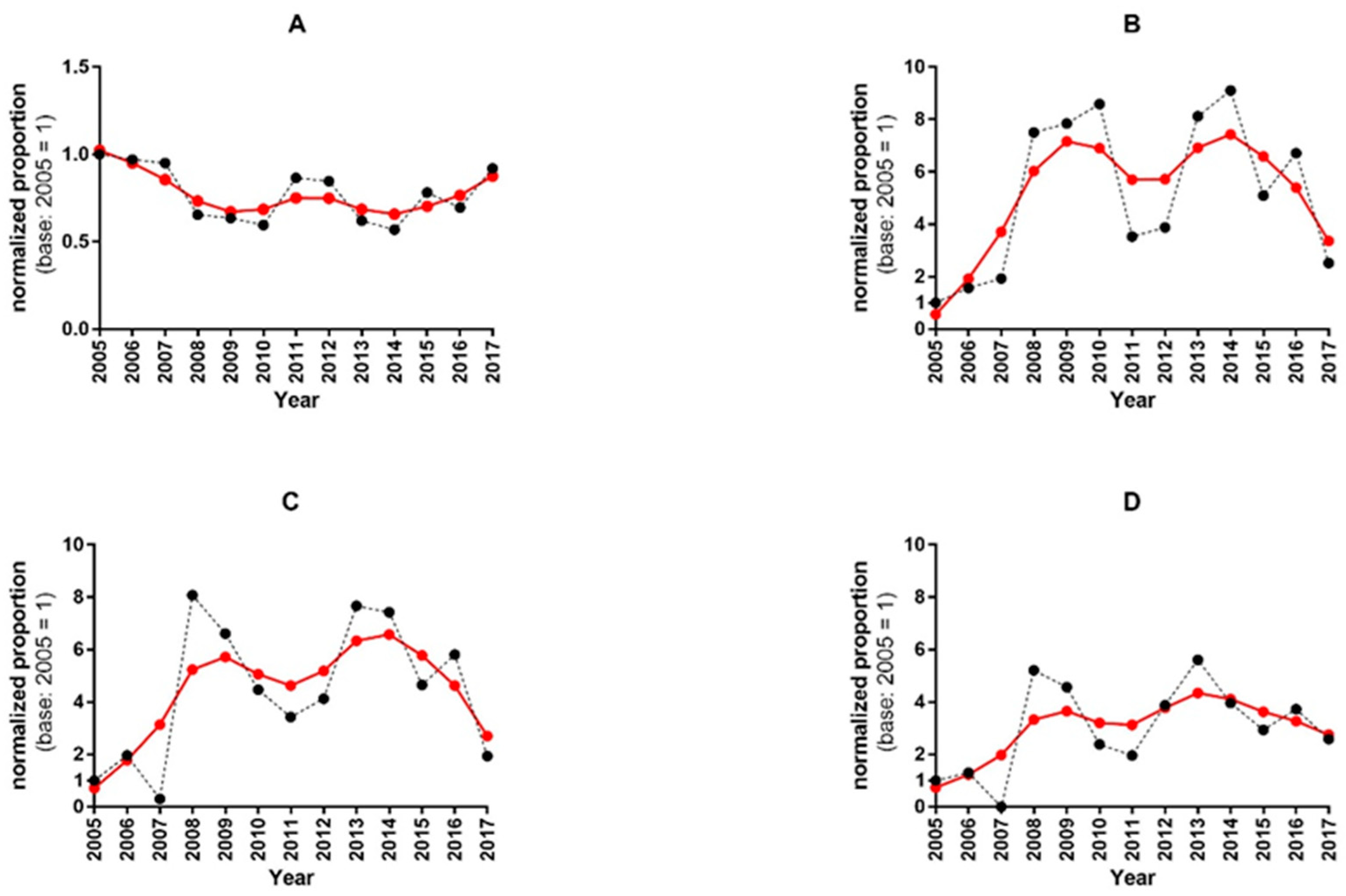
3.2. Analysis of Human Serum Samples for the Presence of IDV-Specific Antibodies
3.3. Pearson’s Chi-Squared Test for Multi-Proportions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.; Liu, R.; Sheng, Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a novel swine influenza virus from oklahoma in 2011 which is distantly related to human influenza c viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Excellence for Influenza Research and Surveillance. Introducing Influenza D: International Committee on Taxonomy of Viruses Names New Virus. Available online: http://www.niaidceirs.org/news/2017/02/introducing-influenza-d-international-committee-on-taxonomy-of-viruses-names-new-virus/ (accessed on 27 November 2019).
- Hause, B.M.; Collin, E.A.; Liu, R.; Huang, B.; Sheng, Z.; Lu, W.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a novel influenza virus in cattle and swine: Proposal for a new genus in the orthomyxoviridae family. MBio 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, Z.; Ran, Z.; Wang, D.; Hoppe, A.D.; Simonson, R.; Chakravarty, S.; Hause, B.M.; Li, F. Genomic and evolutionary characterization of a novel influenza-c-like virus from swine. Arch. Virol. 2014, 159, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asha, K.; Kumar, B. Emerging influenza d virus threat: What we know so far! J. Clin. Med. 2019, 8, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.M.; Wang, S.C.; Peng, C.; Yu, J.M.; Zhuang, Q.Y.; Hou, G.Y.; Liu, S.; Li, J.P.; Chen, J.M. Identification of a potential novel type of influenza virus in bovine in china. Virus Genes 2014, 49, 493–496. [Google Scholar] [CrossRef]
- Ducatez, M.F.; Pelletier, C.; Meyer, G. Influenza D virus in cattle, France, 2011–2014. Emerg. Infect. Dis. 2015, 21, 368–371. [Google Scholar] [CrossRef]
- Collin, E.A.; Sheng, Z.; Lang, Y.; Ma, W.; Hause, B.M.; Li, F. Cocirculation of two distinct genetic and antigenic lineages of proposed influenza d virus in cattle. J. Virol. 2015, 89, 1036–1042. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, L.; Eckard, L.; Epperson, W.B.; Long, L.P.; Smith, D.; Huston, C.; Genova, S.; Webby, R.; Wan, X.F. Influenza d virus infection in mississippi beef cattle. Virology 2015, 486, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, L.; Olivier, A.K.; Genova, S.; Epperson, W.B.; Smith, D.R.; Schneider, L.; Barton, K.; McCuan, K.; Webby, R.J.; Wan, X.F. Pathogenesis of influenza d virus in cattle. J. Virol. 2016, 90, 5636–5642. [Google Scholar] [CrossRef] [Green Version]
- Murakami, S.; Endoh, M.; Kobayashi, T.; Takenaka-Uema, A.; Chambers, J.K.; Uchida, K.; Nishihara, M.; Hause, B.; Horimoto, T. Influenza d virus infection in herd of cattle, Japan. Emerg. Infect. Dis. 2016, 22, 1517–1519. [Google Scholar] [CrossRef]
- Horimoto, T.; Hiono, T.; Mekata, H.; Odagiri, T.; Lei, Z.; Kobayashi, T.; Norimine, J.; Inoshima, Y.; Hikono, H.; Murakami, K.; et al. Nationwide distribution of bovine influenza d virus infection in japan. PLoS ONE 2016, 11, e0163828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Ferguson, L.; Smith, D.R.; Woolums, A.R.; Epperson, W.B.; Wan, X.F. Serological evidence for high prevalence of influenza d viruses in cattle, nebraska, united states, 2003–2004. Virology 2017, 501, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Hill, J.E.; Godson, D.L.; Ngeleka, M.; Fernando, C.; Huang, Y. The pulmonary virome, bacteriological and histopathological findings in bovine respiratory disease from western Canada. Transbound. Emerg. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Dane, H.; Duffy, C.; Guelbenzu, M.; Hause, B.; Fee, S.; Forster, F.; McMenamy, M.J.; Lemon, K. Detection of influenza d virus in bovine respiratory disease samples, UK. Transbound. Emerg. Dis. 2019, 66, 2184–2187. [Google Scholar] [CrossRef]
- Flynn, O.; Gallagher, C.; Mooney, J.; Irvine, C.; Ducatez, M.; Hause, B.; McGrath, G.; Ryan, E. Influenza d virus in cattle, Ireland. Emerg. Infect. Dis. 2018, 24, 389–391. [Google Scholar] [CrossRef] [Green Version]
- Mitra, N.; Cernicchiaro, N.; Torres, S.; Li, F.; Hause, B.M. Metagenomic characterization of the virome associated with bovine respiratory disease in feedlot cattle identified novel viruses and suggests an etiologic role for influenza d virus. J. Gen. Virol. 2016, 97, 1771–1784. [Google Scholar] [CrossRef]
- Snoeck, C.J.; Oliva, J.; Pauly, M.; Losch, S.; Wildschutz, F.; Muller, C.P.; Hubschen, J.M.; Ducatez, M.F. Influenza d virus circulation in cattle and swine, Luxembourg, 2012–2016. Emerg. Infect. Dis. 2018, 24, 1388–1389. [Google Scholar] [CrossRef] [Green Version]
- Quast, M.; Sreenivasan, C.; Sexton, G.; Nedland, H.; Singrey, A.; Fawcett, L.; Miller, G.; Lauer, D.; Voss, S.; Pollock, S.; et al. Serological evidence for the presence of influenza d virus in small ruminants. Vet. Microbiol. 2015, 180, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Salem, E.; Cook, E.A.J.; Lbacha, H.A.; Oliva, J.; Awoume, F.; Aplogan, G.L.; Hymann, E.C.; Muloi, D.; Deem, S.L.; Alali, S.; et al. Serologic evidence for influenza c and d virus among ruminants and camelids, Africa, 1991–2015. Emerg. Infect. Dis. 2017, 23, 1556–1559. [Google Scholar] [CrossRef] [Green Version]
- Zhai, S.L.; Zhang, H.; Chen, S.N.; Zhou, X.; Lin, T.; Liu, R.; Lv, D.H.; Wen, X.H.; Wei, W.K.; Wang, D.; et al. Influenza d virus in animal species in Guangdong province, Southern China. Emerg. Infect. Dis. 2017, 23, 1392–1396. [Google Scholar] [CrossRef]
- Nedland, H.; Wollman, J.; Sreenivasan, C.; Quast, M.; Singrey, A.; Fawcett, L.; Christopher-Hennings, J.; Nelson, E.; Kaushik, R.S.; Wang, D.; et al. Serological evidence for the co-circulation of two lineages of influenza d viruses in equine populations of the Midwest United States. Zoonoses Public Health 2018, 65, e148–e154. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Odagiri, T.; Melaku, S.K.; Bazartseren, B.; Ishida, H.; Takenaka-Uema, A.; Muraki, Y.; Sentsui, H.; Horimoto, T. Influenza d virus infection in dromedary camels, Ethiopia. Emerg. Infect. Dis. 2019, 25, 1224–1226. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, T.; Donohoe, L.; Ducatez, M.F.; Meyer, G.; Ryan, E. Seroprevalence of influenza d virus in selected sample groups of irish cattle, sheep and pigs. Ir. Vet. J. 2019, 72, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foni, E.; Chiapponi, C.; Baioni, L.; Zanni, I.; Merenda, M.; Rosignoli, C.; Kyriakis, C.S.; Luini, M.V.; Mandola, M.L.; Bolzoni, L.; et al. Influenza d in Italy: Towards a better understanding of an emerging viral infection in swine. Sci. Rep. 2017, 7, 11660. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel influenza d virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, L.; Luo, K.; Olivier, A.K.; Cunningham, F.L.; Blackmon, S.; Hanson-Dorr, K.; Sun, H.; Baroch, J.; Lutman, M.W.; Quade, B.; et al. Influenza d virus infection in feral swine populations, United States. Emerg. Infect. Dis. 2018, 24, 1020–1028. [Google Scholar] [CrossRef] [Green Version]
- Chiapponi, C.; Faccini, S.; De Mattia, A.; Baioni, L.; Barbieri, I.; Rosignoli, C.; Nigrelli, A.; Foni, E. Detection of influenza d virus among swine and cattle, Italy. Emerg. Infect. Dis. 2016, 22, 352–354. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.B.; Gaunt, E.R.; Digard, P.; Templeton, K.; Simmonds, P. Detection of influenza c virus but not influenza d virus in Scottish respiratory samples. J. Clin. Virol. 2016, 74, 50–53. [Google Scholar] [CrossRef] [Green Version]
- White, S.K.; Ma, W.; McDaniel, C.J.; Gray, G.C.; Lednicky, J.A. Serologic evidence of exposure to influenza d virus among persons with occupational contact with cattle. J. Clin. Virol. 2016, 81, 31–33. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Perini, D.; Mather, S.; Temperton, N.; Montomoli, E. Overview of serological techniques for influenza vaccine evaluation: Past, present and future. Vaccines 2014, 2, 707–734. [Google Scholar] [CrossRef] [Green Version]
- Clopper, C.J.; Pearson, E.S. The use of confidence or fiducial limits illustrated in the case of the binomial. Biometrika 1934, 26, 404–413. [Google Scholar] [CrossRef]
- Hodrick, R.J.; Prescott, E.C. Postwar U.S. Business cycles: An empirical investigation. J. Money Credit Bank. 1997, 29, 1–16. [Google Scholar] [CrossRef]
- Borkenhagen, L.K.; Mallinson, K.A.; Tsao, R.W.; Ha, S.J.; Lim, W.H.; Toh, T.H.; Anderson, B.D.; Fieldhouse, J.K.; Philo, S.E.; Chong, K.S.; et al. Surveillance for respiratory and diarrheal pathogens at the human-pig interface in Sarawak, Malaysia. PLoS ONE 2018, 13, e0201295. [Google Scholar] [CrossRef] [PubMed]
- Bailey, E.S.; Choi, J.Y.; Zemke, J.; Yondon, M.; Gray, G.C. Molecular surveillance of respiratory viruses with bioaerosol sampling in an airport. Trop. Dis. Travel Med. Vaccines 2018, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, C.; Thomas, M.; Sheng, Z.; Hause, B.M.; Collin, E.A.; Knudsen, D.E.; Pillatzki, A.; Nelson, E.; Wang, D.; Kaushik, R.S.; et al. Replication and transmission of the novel bovine influenza d virus in a guinea pig model. J. Virol. 2015, 89, 11990–12001. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Qi, J.; Khedri, Z.; Diaz, S.; Yu, H.; Chen, X.; Varki, A.; Shi, Y.; Gao, G.F. Correction: An open receptor-binding cavity of hemagglutinin-esterase-fusion glycoprotein from newly-identified influenza d virus: Basis for its broad cell tropism. PLoS Pathog. 2016, 12, e1005505. [Google Scholar] [CrossRef] [Green Version]
- Holwerda, M.; Kelly, J.; Laloli, L.; Sturmer, I.; Portmann, J.; Stalder, H.; Dijkman, R. Determining the replication kinetics and cellular tropism of influenza d virus on primary well-differentiated human airway epithelial cells. Viruses 2019, 11, 337. [Google Scholar] [CrossRef] [Green Version]
- Chiapponi, C.; Faccini, S.; Fusaro, A.; Moreno, A.; Prosperi, A.; Merenda, M.; Baioni, L.; Gabbi, V.; Rosignoli, C.; Alborali, G.L.; et al. Detection of a new genetic cluster of influenza d virus in Italian cattle. Viruses 2019, 11, 1110. [Google Scholar] [CrossRef] [Green Version]
- Odagiri, T.; Ishida, H.; Li, J.Y.; Endo, M.; Kobayashi, T.; Kamiki, H.; Matsugo, H.; Takenaka-Uema, A.; Murakami, S.; Horimoto, T. Antigenic heterogeneity among phylogenetic clusters of influenza d viruses. J. Vet. Med Sci. 2018, 80, 1241–1244. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Influenza Viruses | IDV Antiserum HI Titre | ICV Antiserum HI Titre |
|---|---|---|
| D/bovine/Oklahoma/660/2013 | 1:10,240 | 5 |
| C/Victoria/2/2012 | 5 | 1:640 |
| HI Assay | |||||||||||||||
| Year | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | Total | |
| Samples | 99 | 101 | 82 | 95 | 101 | 83 | 101 | 102 | 100 | 100 | 101 | 115 | 101 | 1281 | |
| Titre | 5 | 94 | 93 | 74 | 59 | 61 | 47 | 83 | 82 | 59 | 54 | 75 | 76 | 89 | 946 |
| ≥1:10 | 5 | 8 | 8 | 36 | 40 | 36 | 18 | 20 | 41 | 46 | 26 | 39 | 12 | 335 | |
| ≥1:20 | 4 | 8 | 1 | 31 | 27 | 15 | 14 | 17 | 31 | 30 | 19 | 27 | 12 | 236 | |
| ≥1:40 | 3 | 4 | 0 | 15 | 14 | 6 | 6 | 12 | 17 | 12 | 9 | 13 | 8 | 119 | |
| ≥1:80 | 2 | 1 | 0 | 7 | 9 | 3 | 2 | 8 | 7 | 2 | 2 | 4 | 5 | 52 | |
| ≥1:160 | 1 | 0 | 0 | 5 | 2 | 0 | 2 | 0 | 2 | 0 | 0 | 3 | 3 | 18 | |
| N≥1:10 | 5 | 8 | 8 | 36 | 40 | 36 | 18 | 20 | 41 | 46 | 26 | 39 | 12 | 335 | |
| %≥1:10 | 5.1% | 7.9% | 9.8% | 37.9% | 39.6% | 43.4% | 17.8% | 19.6% | 41.0% | 46.0% | 25.7% | 33.9% | 11.9% | 26.2% | |
| CI 95% lower | 1.7% | 3.5% | 4.3% | 28.1% | 30.0% | 32.5% | 10.9% | 12.4% | 31.3% | 36.0% | 17.6% | 25.3% | 6.3% | 23.8% | |
| CI 95% upper | 11.4% | 15.0% | 18.3% | 48.4% | 49.8% | 54.7% | 26.7% | 28.6% | 51.3% | 56.3% | 35.4% | 43.3% | 19.8% | 28.6% | |
| (A) | |||||||||||||||
| VN Assay | |||||||||||||||
| Year | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | Total | |
| Samples | 5 | 8 | 8 | 36 | 40 | 36 | 18 | 20 | 41 | 46 | 26 | 39 | 12 | 335 | |
| Titre | 5 | 0 | 1 | 0 | 2 | 7 | 16 | 1 | 5 | 2 | 4 | 4 | 9 | 4 | 55 |
| ≥1:10 | 5 | 7 | 8 | 34 | 33 | 20 | 17 | 15 | 39 | 42 | 22 | 30 | 8 | 280 | |
| ≥1:20 | 5 | 5 | 8 | 33 | 18 | 17 | 15 | 13 | 29 | 35 | 35 | 20 | 7 | 240 | |
| ≥1:40 | 4 | 4 | 2 | 10 | 2 | 5 | 9 | 9 | 12 | 14 | 14 | 7 | 4 | 96 | |
| ≥1:80 | 1 | 1 | 0 | 1 | 0 | 1 | 2 | 0 | 4 | 0 | 0 | 0 | 1 | 11 | |
| ≥1:160 | 0 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 4 | |
| N≥1:10 | 5 | 7 | 8 | 34 | 33 | 20 | 17 | 15 | 39 | 42 | 22 | 30 | 8 | 280 | |
| %≥1:10 | 5.1% | 6.9% | 9.8% | 35.8% | 32.7% | 24.1% | 16.8% | 14.7% | 39.0% | 42.0% | 21.8% | 26.1% | 7.9% | 21.9% | |
| CI 95% lower | 1.7% | 2.8% | 4.3% | 26.2% | 23.7% | 15.4% | 10.1% | 8.5% | 29.4% | 32.2% | 14.2% | 18.3% | 3.5% | 19.6% | |
| CI 95% upper | 11.4% | 13.8% | 18.3% | 46.3% | 42.7% | 34.7% | 25.6% | 23.1% | 49.3% | 52.3% | 31.1% | 35.1% | 15.0% | 24.2% | |
| (B) | |||||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trombetta, C.M.; Marchi, S.; Manini, I.; Kistner, O.; Li, F.; Piu, P.; Manenti, A.; Biuso, F.; Sreenivasan, C.; Druce, J.; et al. Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017. Viruses 2020, 12, 30. https://doi.org/10.3390/v12010030
Trombetta CM, Marchi S, Manini I, Kistner O, Li F, Piu P, Manenti A, Biuso F, Sreenivasan C, Druce J, et al. Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017. Viruses. 2020; 12(1):30. https://doi.org/10.3390/v12010030
Chicago/Turabian StyleTrombetta, Claudia M., Serena Marchi, Ilaria Manini, Otfried Kistner, Feng Li, Pietro Piu, Alessandro Manenti, Fabrizio Biuso, Chithra Sreenivasan, Julian Druce, and et al. 2020. "Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017" Viruses 12, no. 1: 30. https://doi.org/10.3390/v12010030
APA StyleTrombetta, C. M., Marchi, S., Manini, I., Kistner, O., Li, F., Piu, P., Manenti, A., Biuso, F., Sreenivasan, C., Druce, J., & Montomoli, E. (2020). Influenza D Virus: Serological Evidence in the Italian Population from 2005 to 2017. Viruses, 12(1), 30. https://doi.org/10.3390/v12010030