Occurrence and Molecular Phylogeny of Honey Bee Viruses in Vespids

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
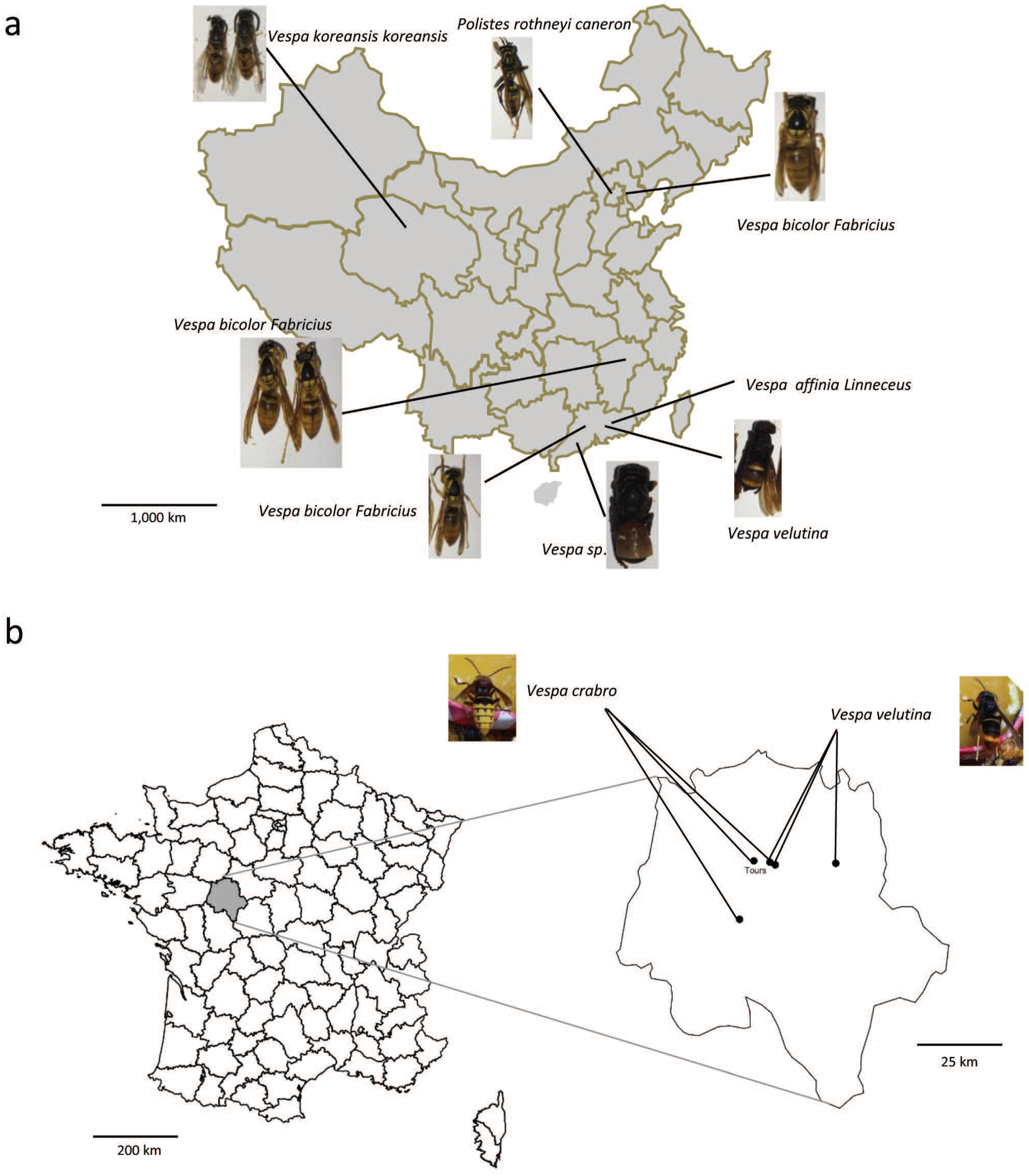
2.1. Samples
2.2. Identification of Vespid Species
2.3. Extraction of DNA and RNA for Virus Identification
2.4. Identification of Honey Bee Viruses via PCR
2.5. Sequencing and Phylogenetic Analyses
3. Results
3.1. Virus Occurrence
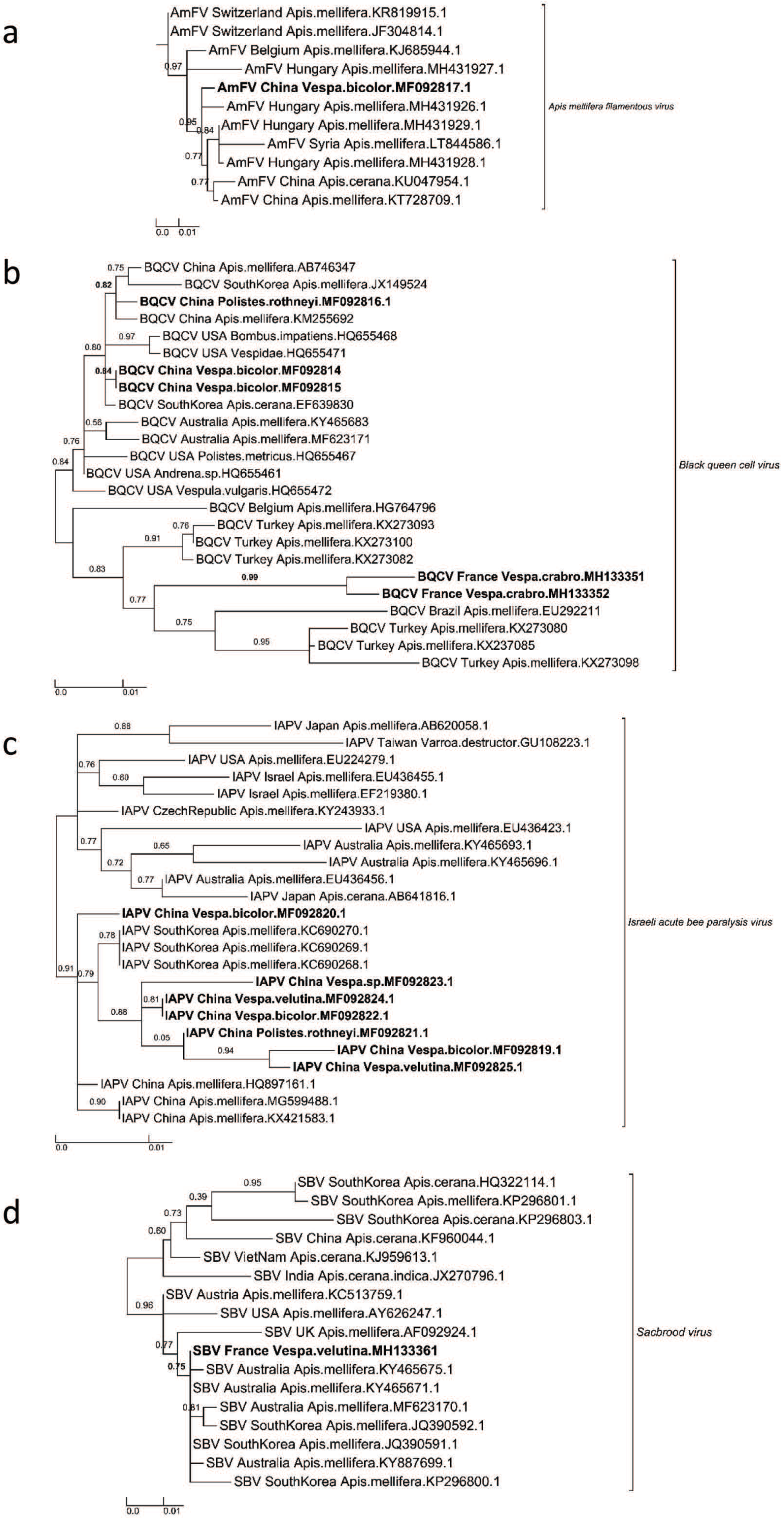
3.2. Phylogenetic Relationships Among the Virus Sequences Found in Vespids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klein, A.-M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Brutscher, L.M.; McMenamin, A.J.; Flenniken, M.L. The buzz about honey bee viruses. PLoS Pathog. 2016, 12, e1005757. [Google Scholar] [CrossRef]
- Wilfert, L.; Long, G.; Leggett, H.C.; Schmid-Hempel, P.; Butlin, R.; Martin, S.J.M.; Boots, M. Deformed wing virus is a recent global epidemic in honey bees driven by Varroa mites. Science 2016, 351, 594–597. [Google Scholar] [CrossRef] [Green Version]
- Di Prisco, G.; Pennacchio, F.; Caprio, E.; Boncristiani, H.F.; Evans, J.D.; Chen, Y.P. Varroa destructor is an effective vector of Israeli acute paralysis virus in the honey bee, Apis mellifera. J. Gen. Virol. 2011, 92, 151–155. [Google Scholar] [CrossRef]
- Takeuchi, T.; Takahashi, R.; Kiyoshi, T.; Nakamura, M.; Minoshima, Y.; Takahahi, J. The origin and genetic diversity of the yellow-legged hornet, Vespa velutina introduced in Japan. Insectes Sociaux 2017, 3, 313–320. [Google Scholar] [CrossRef]
- Perrard, A.; Haxaire, J.; Rortais, A.; Villemant, C. Observations on the colony activity of the Asian hornet Vespa velutina Lepeletier 1836 (Hymenoptera: Vespidae: Vespinae) in France. Ann. Soc. Entomol. Fr. 2009, 45, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Choi, M.; Moon, T.Y. Occurrence of Vespa velutina Lepeletier from Korea, and a revised key for Korean Vespa species (Hymenoptera:Vespidae). Entomol. Res. 2006, 2, 112–115. [Google Scholar] [CrossRef]
- Choi, M.B.; Martin, S.J.; Lee, J.W. Distribution, spread, and impact of the invasive hornet Vespa velutina in South Korea. Entomol. Res. 2012, 15, 473–477. [Google Scholar] [CrossRef]
- Villemant, C.; Zuccon, D.; Rome, Q.; Muller, F.; Poinar, G.O., Jr.; Justine, J.L. Can parasites halt the invader? Mermithid nematodes parasitizing the yellow-legged Asian hornet in France. PeerJ 2015, 3, e947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haxaire, J.; Bouguet, J.P.; Tamisier, J.P. Vespa velutina Lepeletier, 1836, une redoutable nouveauté pour la faune de France (Hym., Vespidae). Bull. Soc. Entomol. Fr. 2006, 111, 194. [Google Scholar]
- Villemant, C.; Barbet-Massin, M.; Perrard, A.; Muller, F.; Gargominy, O.; Jiguet, F.; Rome, Q. Predicting the invasion risk by the alien bee-hawking Yellow-legged hornet Vespa velutina nigrithorax across Europe and other continents with niche models. Biol. Conserv. 2011, 144, 2142–2150. [Google Scholar] [CrossRef]
- Robinet, C.; Darrouzet, E.; Suppo, C. Spread modelling: A suitable tool to explore the role of human-mediated dispersal in the range expansion of the yellow-legged hornet in Europe. Int. J. Pest Manag. 2018, 1, 1–10. [Google Scholar] [CrossRef]
- Rome, Q.; Dambrine, L.; Onate, C.; Muller, F.; Villemant, C.; Pérez, A.L.G.; Maia, M.; Carvalho, P.; Bruneau, E. Spread of the invasive hornet Vespa velutina Lepeletier, 1836, in Europe in 2012 (Hym., Vespidae). Bull. Soc. Entomol. Fr. 2013, 118, 21–22. [Google Scholar]
- Demichelis, S.; Manino, A.; Minuto, G.; Mariotti, M.; Porporato, M. Social wasp trapping in north west Italy: Comparison of different bait-traps and first detection of Vespa velutina. Bull. Insectol. 2014, 67, 307–317. [Google Scholar]
- Tan, K.; Wu, J.; Liu, Q.; Zhou, D.Y. Reduction of foraging activity by A.cerana colonies attacked by Vespa Velutina. J. Bee 2004, 2, 7–8. [Google Scholar]
- Requier, F.; Rome, Q.; Chiron, G.; Decante, D.; Marion, S.; Menard, M.; Muller, F.; Villemant, C.; Henry, M. Predation of the invasive Asian hornet affects foraging activity and survival probability of honey bees in Western Europe. J. Pest Sci. 2019, 92, 567–578. [Google Scholar] [CrossRef]
- Yañez, O.; Zheng, H.Q.; Hu, F.L.; Neumann, P.; Dietemann, V. A scientific note on Israeli acute paralysis virus infection of Eastern honeybee Apis cerana and vespine predator Vespa velutina. Apidologie 2012, 5, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Garigliany, M.; Taminiau, B.; El Agrebi, N.; Cadar, D.; Gilliaux, G.; Hue, M.; Desmecht, D.; Daube, G.; Linden, A.; Farnir, F. Moku virus in invasive Asian hornets, Belgium, 2016. Emerg. Infect. Dis. 2017, 12, 2109. [Google Scholar] [CrossRef] [Green Version]
- Mazzei, M.; Forzan, M.; Cilia, G.; Sagona, S.; Bortolotti, L.; Felicioli, A. First detection of replicative deformed wing virus (DWV) in Vespa velutina nigrithorax. Bull. Insectol. 2018, 2, 211–216. [Google Scholar]
- Yue, Q.Y.; Qiu, D.Y.; Huang, Y.W.; Liu, G.X. Application of DNA barcoding to species identification of an unknown insect larva. Chin. J. Health Lab. Technol. 2011, 3, 615–617. [Google Scholar]
- Matsuura, M. Vespa and Provespa. In The Social Biology of Wasps; Ross, K.G., Matthews, R.W., Eds.; Cornell University Press: Ithaca, NY, USA, 1991; p. 30. [Google Scholar]
- Bigot, D.; Dalmon, A.; Roy, B.; Hou, C.; Germain, M.; Romary, M. The discovery of Halictivirus resolves the Sinaivirus phylogeny. J. Gen. Virol. 2017, 98, 2864–2875. [Google Scholar] [CrossRef] [PubMed]
- Sguazza, G.H.; Reynaldi, F.J.; Galosi, C.M.; Pecoraro, M.R. Simultaneous detection of bee viruses by multiplex PCR. J. Virol. Methods. 2013, 194, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 3, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart model selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Chen, G.; Wang, Z.W.; Qin, Y.; Sun, W.B. Seed dispersal by hornets: An unusual insect-plant mutualism. J. Integr. Plant Biol. 2017, 59, 792–796. [Google Scholar] [CrossRef] [Green Version]
- López, S.; González, M.; Goldarazena, A. Vespa velutina Lepeletier, 1836 (Hymenoptera: Vespidae): First records in Iberian Peninsula. Eppo Bull. 2011, 41, 439–441. [Google Scholar] [CrossRef]
- Boncristiani, H.F.; Evans, J.D.; Chen, Y.; Pettis, J.; Murphy, C.; Lopez, D.L.; Simone-Finstrom, M.; Strand, M.; Tarpy, D.R.; Rueppell, O. In vitro infection of pupae with Israeli acute paralysis virus suggests disturbance of transcriptional homeostasis in honey bees (Apis mellifera). PLoS ONE 2013, 9, e73429. [Google Scholar] [CrossRef] [PubMed]
- Ai, H.; Yan, X.; Han, R. Occurrence and prevalence of seven bee viruses in Apis mellifera and Apis cerana apiaries in China. J. Invertebr. Pathol. 2012, 1, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Dalmon, A.; Gayral, P.; Decante, D.; Klopp, C.; Bigot, D.; Thomasson, M.; Herniou, E.A.; Alaux, C.; Conte, Y.L. Viruses in the invasive hornet Vespa velutina. Viruses 2019, 11, 1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maori, E.; Lavi, S.; Mozes-Koch, R.; Gantman, Y.; Peretz, Y.; Edelbaum, O.; Tanne, E.; Sela, I. Isolation and characterization of Israeli acute paralysis virus, a dicistrovirus affecting honey bees in Israel: Evidence for diversity due to intra-and inter-species recombination. J. Gen. Virol. 2008, 88, 3428–3438. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Hui, J.; Quan, P.L.; Kalkstein, A.; Honkavuori, K.S.; Bussetti, A.V.; Conlan, S.; Evans, J.; Chen, Y.P.; VanEngelsdorp, D.; et al. Genetic analysis of Israel acute paralysis virus: Distinct clusters are circulating in the United States. J. Virol. 2008, 13, 6209–6217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diao, Q.; Li, B.; Zhao, H.; Wu, Y.; Guo, R.; Dai, P.; Chen, D.; Wang, Q.; Hou, C. Enhancement of chronic bee paralysis virus levels in honeybees acute exposed to imidacloprid: A Chinese case study. Sci. Total Environ. 2018, 630, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Clark, T.B. A filamentous virus of the honey bee. J. Invertebr. Pathol. 1978, 32, 332–340. [Google Scholar] [CrossRef]
- Gauthier, L.; Cornman, S.; Hartmann, U.; Cousserans, F.; Evans, J.D.; De Miranda, J.R.; Neumann, P. The Apis mellifera filamentous virus genome. Viruses 2015, 7, 3798–3815. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, U.; Forsgren, E.; Charrière, J.D.; Neumann, P.; Gauthier, L. Dynamics of Apis mellifera filamentous virus (AmFV) infections in honey bees and relationships with other parasites. Viruses 2015, 7, 2654–2667. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Country | Developmental Stage | Colony ID/Province (Species) | AmFV | IAPV | DWV-A | DWV-B | DWV-A/KV | CSBV | SBV | BQCV | LSV |
|---|---|---|---|---|---|---|---|---|---|---|---|
| China | Adult | B1/Beijing (Vespa bicolor) | nt | nt | + | nt | + | nt | nt | nt | nt |
| Pupa | + | + | + | nt | + | nt | nt | nt | nt | ||
| Adult | B2/Beijing (Vespa bicolor) | nt | nt | + | nt | nt | nt | nt | + | nt | |
| Pupa | + | + | + | nt | + | + | nt | + | + | ||
| Adult | B3/Beijing (Vespa bicolor) | nt | nt | nt | nt | nt | nt | nt | nt | nt | |
| Pupa | nt | nt | nt | nt | nt | nt | nt | nt | nt | ||
| Adult | B4/Beijing (Polistes rothneyi) | + | nt | nt | nt | nt | nt | nt | nt | nt | |
| Pupa | + | nt | nt | nt | nt | nt | nt | + | nt | ||
| Adult | B5/Beijing (Polistes rothneyi) | nt | nt | nt | nt | nt | nt | nt | nt | nt | |
| Pupa | + | + | nt | nt | nt | nt | nt | nt | nt | ||
| Adult | Q1/Qinghai (Vespa koreansis) | nt | nt | nt | nt | nt | nt | nt | nt | nt | |
| Adult | GD1/Guangdong (Vespa affinia) | nt | nt | + | nt | nt | nt | nt | nt | nt | |
| Pupa | nt | + | + | nt | nt | nt | nt | nt | nt | ||
| Adult | GD2/Guangdong (Vespa velutina) | nt | nt | nt | nt | nt | nt | nt | nt | nt | |
| Pupa | nt | + | nt | nt | nt | nt | nt | nt | nt | ||
| Adult | GD3/Guangdong (Vespa velutina) | nt | nt | nt | nt | nt | nt | nt | nt | nt | |
| Pupa | nt | + | nt | nt | nt | nt | nt | nt | nt | ||
| Adult | GD4/Guangdong (Vespa sp.) | nt | nt | nt | nt | nt | nt | nt | nt | nt | |
| Pupa | nt | + | + | + | + | nt | nt | + | nt | ||
| Adult | GD5/Guangdong (Vespa bicolor) | nt | + | + | nt | nt | nt | nt | nt | nt | |
| Pupa | nt | + | + | nt | nt | nt | nt | nt | nt | ||
| Adult | JX/Jiangxi (Vespa bicolor) | nt | nt | nt | nt | nt | nt | nt | nt | nt | |
| Pupa | nt | nt | nt | nt | nt | nt | nt | nt | nt | ||
| France | Adult | Fondettes (Vespa crabro) | nt | nt | + | nt | nt | nt | nt | nt | nt |
| Adult | Saché (Vespa crabro) | nt | nt | + | nt | nt | nt | nt | nt | nt | |
| Adult | Saché (Vespa crabro) | nt | nt | + | nt | nt | nt | nt | nt | nt | |
| Adult | Lussault-sur-Loire (Vespa velutina) | nt | nt | + | nt | nt | nt | nt | nt | nt | |
| Adult | Lussault-sur-Loire (Vespa velutina) | nt | nt | nt | nt | nt | nt | + | nt | nt | |
| Adult | Fondettes (Vespa crabro) | nt | nt | nt | nt | nt | nt | nt | + | nt | |
| Adult | Tours (Vespa crabro) | nt | nt | nt | nt | nt | nt | nt | + | nt |
| Colony ID | Vespid Species | Developmental Stage | Viruses Present |
|---|---|---|---|
| B2 | Vespa bicolor | Adult | DWV+BQCV |
| GD5 | Vespa bicolor | Adult | IAPV+DWV |
| B4 | Polistes rothneyi | Pupa | AmFV+BQCV |
| B5 | Polistes rothneyi | Pupa | AmFV+IAPV |
| GD1 | Vespa affinia | Pupa | DWV+IAPV |
| B1 | Vespa bicolor | Pupa | AmFV+IAPV+DWV |
| GD4 | Vespa sp. | Pupa | IAPV+DWV+BQCV |
| B2 | Vespa bicolor | Pupa | AmFV+IAPV+DWV+CSBV+BQCV+LSV |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Gayral, P.; Zhao, H.; Wu, Y.; Jiang, X.; Wu, Y.; Bigot, D.; Wang, X.; Yang, D.; Herniou, E.A.; et al. Occurrence and Molecular Phylogeny of Honey Bee Viruses in Vespids. Viruses 2020, 12, 6. https://doi.org/10.3390/v12010006
Yang S, Gayral P, Zhao H, Wu Y, Jiang X, Wu Y, Bigot D, Wang X, Yang D, Herniou EA, et al. Occurrence and Molecular Phylogeny of Honey Bee Viruses in Vespids. Viruses. 2020; 12(1):6. https://doi.org/10.3390/v12010006
Chicago/Turabian StyleYang, Sa, Philippe Gayral, Hongxia Zhao, Yaojun Wu, Xuejian Jiang, Yanyan Wu, Diane Bigot, Xinling Wang, Dahe Yang, Elisabeth A. Herniou, and et al. 2020. "Occurrence and Molecular Phylogeny of Honey Bee Viruses in Vespids" Viruses 12, no. 1: 6. https://doi.org/10.3390/v12010006
APA StyleYang, S., Gayral, P., Zhao, H., Wu, Y., Jiang, X., Wu, Y., Bigot, D., Wang, X., Yang, D., Herniou, E. A., Deng, S., Li, F., Diao, Q., Darrouzet, E., & Hou, C. (2020). Occurrence and Molecular Phylogeny of Honey Bee Viruses in Vespids. Viruses, 12(1), 6. https://doi.org/10.3390/v12010006