Construction of Stable Reporter Flaviviruses and Their Applications


Abstract
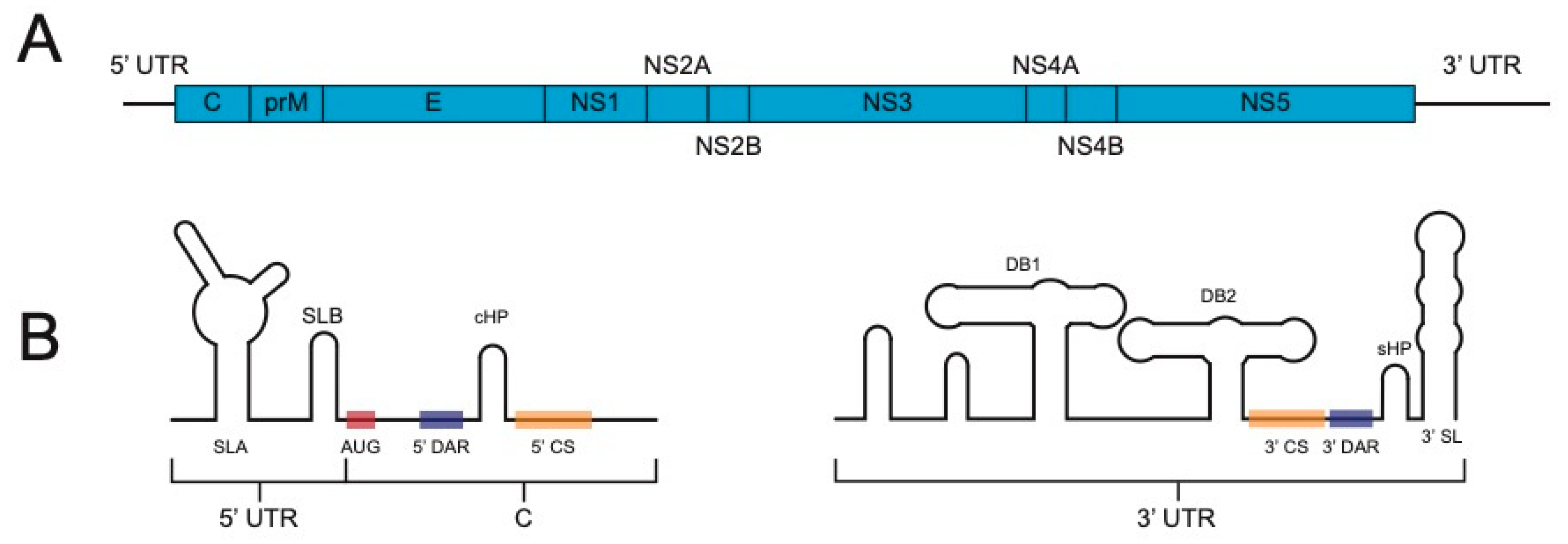
:1. Introduction
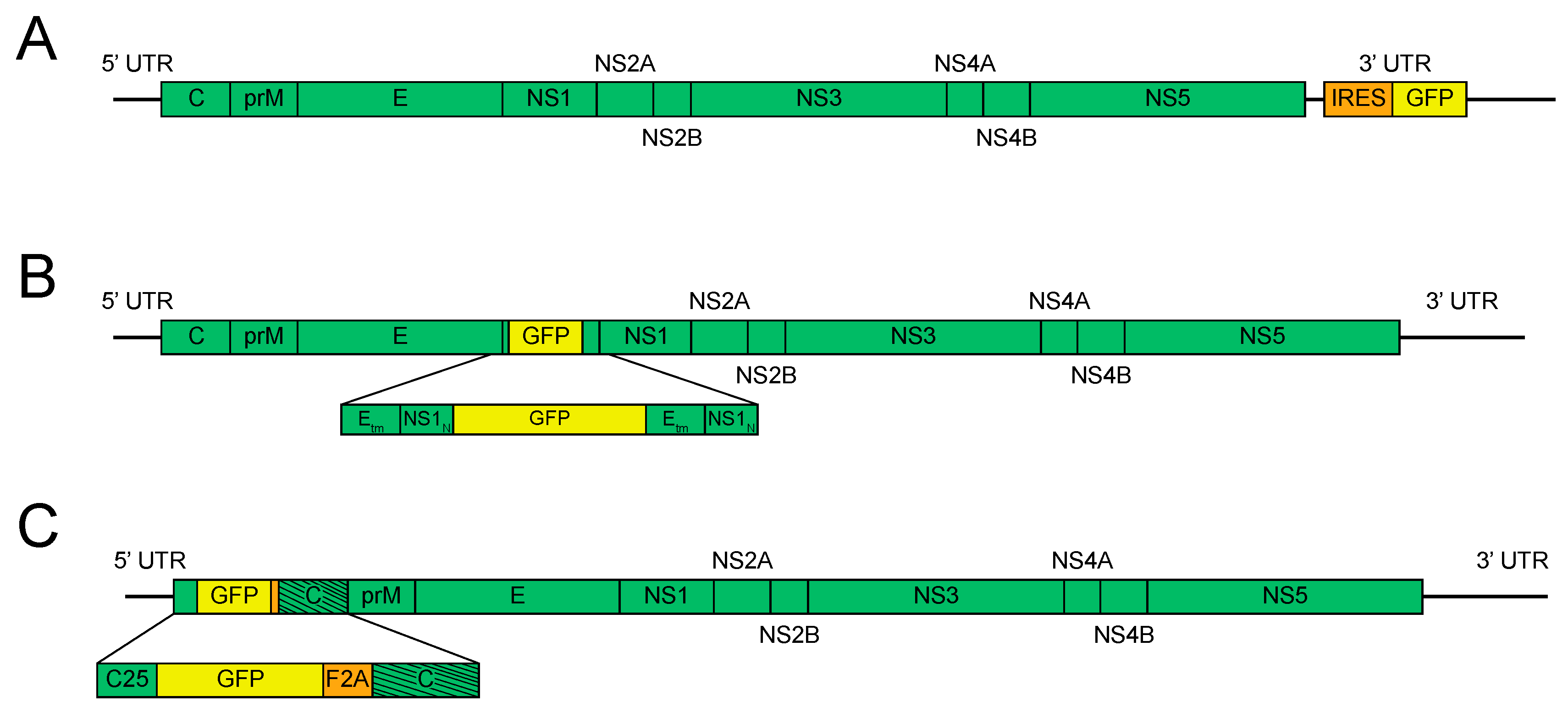
2. Timeline of Reporter Flavivirus Design
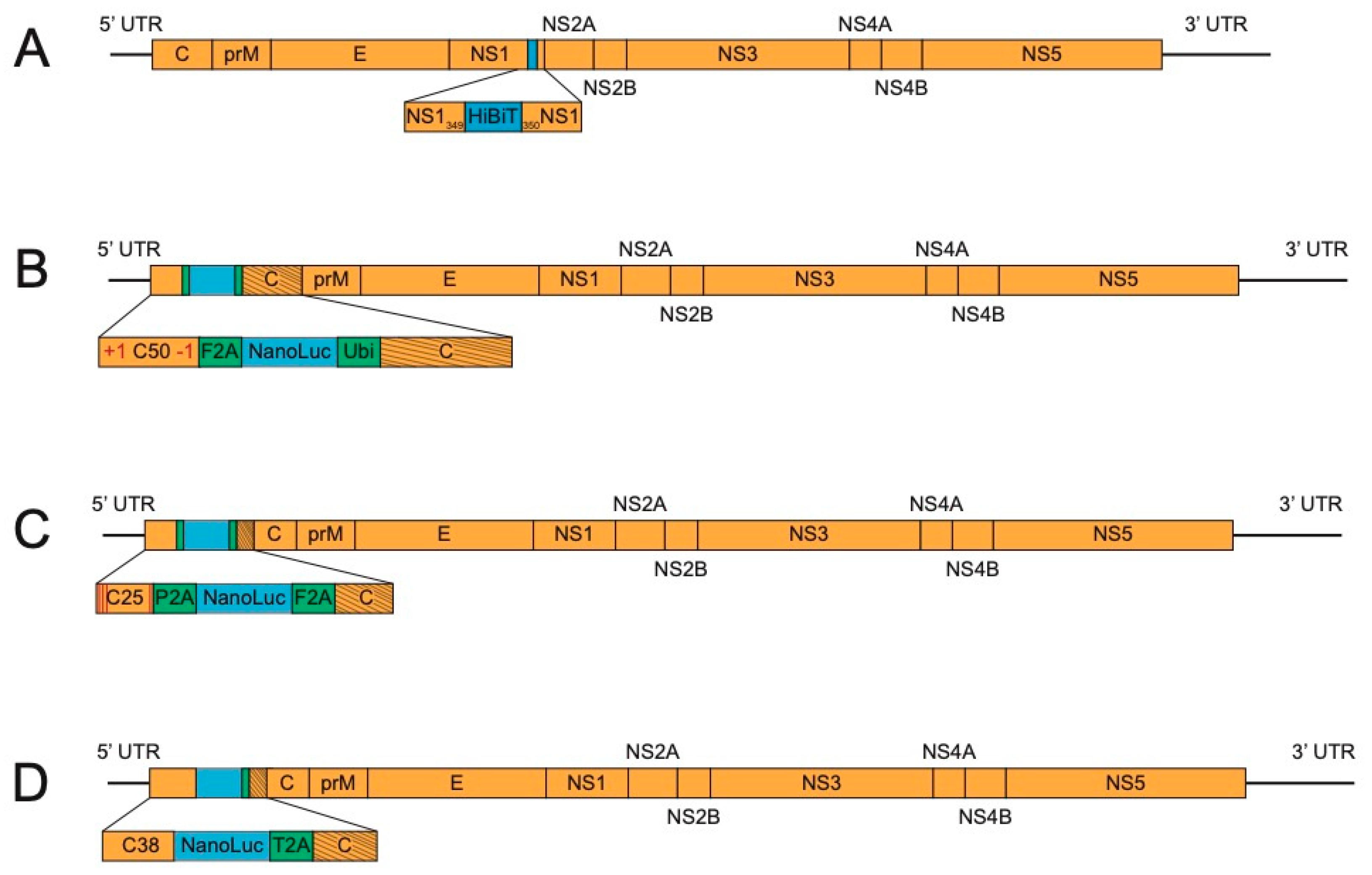
3. Methods for Further Stabilization of Reporter Flaviviruses
4. Applications of Reporter Flaviviruses-Present and Future
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 1–17. [Google Scholar] [CrossRef]
- De Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Dawood, F.S.; Iuliano, A.D.; Reed, C.; Meltzer, M.I.; Shay, D.K.; Cheng, P.-Y.; Bandaranayake, D.; Breiman, R.F.; Brooks, W.A.; Buchy, P.; et al. Estimated global mortality associated with the first 12 months of 2009 pandemic influenza A H1N1 virus circulation: A modelling study. Lancet Infect. Dis. 2012, 12, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Malvy, D.; McElroy, A.K.; de Clerck, H.; Günther, S.; van Griensven, J. Ebola virus disease. Lancet Lond. Engl. 2019, 393, 936–948. [Google Scholar] [CrossRef] [Green Version]
- Baud, D.; Gubler, D.J.; Schaub, B.; Lanteri, M.C.; Musso, D. An update on Zika virus infection. Lancet 2017, 390, 2099–2109. [Google Scholar] [CrossRef] [Green Version]
- Dong, E.; Du, H.; Gardner, L. An interactive web-based dashboard to track COVID-19 in real time. Lancet Infect. Dis. 2020, 20, 533–534. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504. [Google Scholar] [CrossRef]
- Ishikawa, T.; Yamanaka, A.; Konishi, E. A review of successful flavivirus vaccines and the problems with those flaviviruses for which vaccines are not yet available. Vaccine 2014, 32, 1326–1337. [Google Scholar] [CrossRef]
- Brinton, M.A.; Basu, M. Functions of the 3’ and 5’ genome RNA regions of members of the genus Flavivirus. Virus Res. 2015, 206, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Aubry, F.; Nougairède, A.; Gould, E.A.; de Lamballerie, X. Flavivirus reverse genetic systems, construction techniques and applications: A historical perspective. Antivir. Res. 2015, 114, 67–85. [Google Scholar] [CrossRef] [Green Version]
- Guy, B.; Guirakhoo, F.; Barban, V.; Higgs, S.; Monath, T.P.; Lang, J. Preclinical and clinical development of YFV 17D-based chimeric vaccines against dengue, West Nile and Japanese encephalitis viruses. Vaccine 2010, 28, 632–649. [Google Scholar] [CrossRef] [PubMed]
- Fontes-Garfias, C.R.; Baker, C.K.; Shi, P.-Y. Reserve genetic approaches for the development of Zika vaccines and therapeutics. Curr. Opin. Virol. 2020, 44, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kümmerer, B.M. Establishment and application of flavivirus replicons. Adv. Exp. Med. Biol. 2018, 1062, 165–173. [Google Scholar] [CrossRef]
- Khromykh, A.A.; Westaway, E.G. Subgenomic replicons of the flavivirus Kunjin: Construction and applications. J. Virol. 1997, 71, 1497–1505. [Google Scholar] [CrossRef] [Green Version]
- Varnavski, A.N.; Khromykh, A.A. Noncytopathic flavivirus replicon RNA-based system for expression and delivery of heterologous genes. Virology 1999, 255, 366–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.-I.; Kim, S.-Y.; Rice, C.M.; Lee, Y.-M. Development and application of a reverse genetics system for Japanese encephalitis virus. J. Virol. 2003, 77, 6450–6465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierson, T.C.; Diamond, M.S.; Ahmed, A.A.; Valentine, L.E.; Davis, C.W.; Samuel, M.A.; Hanna, S.L.; Puffer, B.A.; Doms, R.W. An infectious West Nile Virus that expresses a GFP reporter gene. Virology 2005, 334, 28–40. [Google Scholar] [CrossRef] [Green Version]
- Deas, T.S.; Binduga-Gajewska, I.; Tilgner, M.; Ren, P.; Stein, D.A.; Moulton, H.M.; Iversen, P.L.; Kauffman, E.B.; Kramer, L.D.; Shi, P.-Y. Inhibition of flavivirus infections by antisense oligomers specifically suppressing viral translation and RNA replication. J. Virol. 2005, 79, 4599–4609. [Google Scholar] [CrossRef] [Green Version]
- Mondotte, J.A.; Lozach, P.-Y.; Amara, A.; Gamarnik, A.V. Essential role of dengue virus envelope protein N glycosylation at asparagine-67 during viral propagation. J. Virol. 2007, 81, 7136–7148. [Google Scholar] [CrossRef] [Green Version]
- Bonaldo, M.C.; Mello, S.M.; Trindade, G.F.; Rangel, A.A.; Duarte, A.S.; Oliveira, P.J.; Freire, M.S.; Kubelka, C.F.; Galler, R. Construction and characterization of recombinant flaviviruses bearing insertions between E and NS1 genes. Virol. J. 2007, 4, 115. [Google Scholar] [CrossRef] [Green Version]
- Shustov, A.V.; Mason, P.W.; Frolov, I. Production of Pseudoinfectious yellow fever virus with a two-component genome. J. Virol. 2007, 81, 11737–11748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samsa, M.M.; Mondotte, J.A.; Iglesias, N.G.; Assunção-Miranda, I.; Barbosa-Lima, G.; Poian, A.T.D.; Bozza, P.T.; Gamarnik, A.V. Dengue virus capsid protein usurps lipid droplets for viral particle formation. PLoS Pathog. 2009, 5, e1000632. [Google Scholar] [CrossRef] [PubMed]
- Samsa, M.M.; Mondotte, J.A.; Caramelo, J.J.; Gamarnik, A.V. Uncoupling cis-acting RNA elements from coding sequences revealed a requirement of the N-terminal region of dengue virus capsid protein in virus particle formation. J. Virol. 2012, 86, 1046–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaptein, S.J.F.; Burghgraeve, T.D.; Froeyen, M.; Pastorino, B.; Alen, M.M.F.; Mondotte, J.A.; Herdewijn, P.; Jacobs, M.; de Lamballerie, X.; Schols, D.; et al. A derivate of the antibiotic doxorubicin is a selective inhibitor of dengue and yellow fever virus replication in vitro. Antimicrob. Agents Chemother. 2010, 54, 5269–5280. [Google Scholar] [CrossRef] [Green Version]
- Zou, G.; Xu, H.Y.; Qing, M.; Wang, Q.-Y.; Shi, P.-Y. Development and characterization of a stable luciferase dengue virus for high-throughput screening. Antivir. Res. 2011, 91, 11–19. [Google Scholar] [CrossRef]
- Schoggins, J.W.; Dorner, M.; Feulner, M.; Imanaka, N.; Murphy, M.Y.; Ploss, A.; Rice, C.M. Dengue reporter viruses reveal viral dynamics in interferon receptor-deficient mice and sensitivity to interferon effectors in vitro. Proc. Natl. Acad. Sci. USA 2012, 109, 14610–14615. [Google Scholar] [CrossRef] [Green Version]
- de Santana, M.G.V.; Neves, P.C.C.; dos Santos, J.R.; Lima, N.S.; dos Santos, A.A.C.; Watkins, D.I.; Galler, R.; Bonaldo, M.C. Improved genetic stability of recombinant yellow fever 17D virus expressing a lentiviral Gag gene fragment. Virology 2014, 452–453, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Shan, C.; Xie, X.; Muruato, A.E.; Rossi, S.L.; Roundy, C.M.; Azar, S.R.; Yang, Y.; Tesh, R.B.; Bourne, N.; Barrett, A.D.; et al. An infectious cDNA CLONE of zika virus to study viral virulence, mosquito transmission, and antiviral inhibitors. Cell Host Microbe 2016, 19, 891–900. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.-T.; Shan, C.; Li, X.-D.; Liu, S.-Q.; Deng, C.-L.; Ye, H.-Q.; Shang, B.-D.; Shi, P.-Y.; Lv, M.; Shen, B.-F.; et al. Generation of a recombinant West Nile virus stably expressing the Gaussia luciferase for neutralization assay. Virus Res. 2016, 211, 17–24. [Google Scholar] [CrossRef]
- Gadea, G.; Bos, S.; Krejbich-Trotot, P.; Clain, E.; Viranaicken, W.; El-Kalamouni, C.; Mavingui, P.; Desprès, P. A robust method for the rapid generation of recombinant Zika virus expressing the GFP reporter gene. Virology 2016, 497, 157–162. [Google Scholar] [CrossRef]
- Grabowski, J.M.; Tsetsarkin, K.A.; Long, D.; Scott, D.P.; Rosenke, R.; Schwan, T.G.; Mlera, L.; Offerdahl, D.K.; Pletnev, A.G.; Bloom, M.E. Flavivirus infection of Ixodes Scapularis (black-legged tick) ex vivo organotypic cultures and applications for disease control. mBio 2017, 8, e01255-17. [Google Scholar] [CrossRef] [Green Version]
- Mutso, M.; Saul, S.; Rausalu, K.; Susova, O.; Žusinaite, E.; Mahalingam, S.; Merits, A. Reverse genetic system, genetically stable reporter viruses and packaged Subgenomic replicon based on a Brazilian Zika virus isolate. J. Gen. Virol. 2017, 98, 2712–2724. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-F.; Li, X.-D.; Deng, C.-L.; Dong, H.-L.; Zhang, Q.-Y.; Ye, Q.; Ye, H.-Q.; Huang, X.-Y.; Deng, Y.-Q.; Zhang, B.; et al. Visualization of a neurotropic flavivirus infection in mouse reveals unique Viscerotropism controlled by host type I interferon signaling. Theranostics 2017, 7, 912–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, T.; Fukuhara, T.; Uchida, T.; Ono, C.; Mori, H.; Sato, A.; Fauzyah, Y.; Okamoto, T.; Kurosu, T.; Setoh, Y.X.; et al. Characterization of recombinant Flaviviridae viruses possessing a small reporter tag. J. Virol. 2017, 92, e01582-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suphatrakul, A.; Duangchinda, T.; Jupatanakul, N.; Prasittisa, K.; Onnome, S.; Pengon, J.; Siridechadilok, B. Multi-color fluorescent reporter dengue viruses with improved stability for analysis of a multi-virus infection. PLoS ONE 2018, 13, e0194399. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Liu, P.; Wang, T.; Wu, Y.; Lin, X.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Zhao, X.; et al. Genetically stable reporter virus, Subgenomic replicon and packaging system of duck Tembusu virus based on a reverse genetics system. Virology 2019, 533, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Igarashi, M.; Enkhbold, B.; Suzuki, T.; Okamatsu, M.; Ono, C.; Mori, H.; Izumi, T.; Sato, A.; Fauzyah, Y.; et al. In vivo dynamics of reporter Flaviviridae viruses. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.-I.; Song, B.-H.; Woolley, M.E.; Frank, J.C.; Julander, J.G.; Lee, Y.-M. Development, characterization, and application of two reporter-expressing recombinant zika viruses. Viruses 2020, 12, 572. [Google Scholar] [CrossRef] [PubMed]
- Volkova, E.; Tsetsarkin, K.A.; Sippert, E.; Assis, F.; Liu, G.; Rios, M.; Pletnev, A.G. Novel approach for insertion of heterologous sequences into full-length ZIKV genome results in superior level of gene expression and insert stability. Viruses 2020, 12, 61. [Google Scholar] [CrossRef] [Green Version]
- Baker, C.; Xie, X.; Zou, J.; Muruato, A.; Fink, K.; Shi, P.-Y. Using recombination-dependent lethal mutations to stabilize reporter flaviviruses for rapid serodiagnosis and drug discovery. Ebiomedicine 2020, 57, 102838. [Google Scholar] [CrossRef]
- Baker, C.; Liu, Y.; Zou, J.; Muruato, A.; Xie, X.; Shi, P.Y. Identifying optimal capsid duplication length for the stability of reporter flaviviruses. Emerg. Microbes Infect. 2020. [Google Scholar] [CrossRef]
- Shi, P.-Y.; Tilgner, M.; Lo, M.K. Construction and characterization of Subgenomic replicons of New York strain of west Nile virus. Virology 2002, 296, 219–233. [Google Scholar] [CrossRef]
- Blight, K.J.; Kolykhalov, A.A.; Rice, C.M. Efficient initiation of HCV RNA replication in cell culture. Science 2000, 290, 1972–1974. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, V.; Körner, F.; Koch, J.-O.; Herian, U.; Theilmann, L.; Bartenschlager, R. Replication of Subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science 1999, 285, 110–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, C.M.; Grakoui, A.; Galler, R.; Chambers, T.J. Transcription of infectious yellow fever RNA from full-length cDNA templates produced by in vitro ligation. New Biol. 1989, 1, 285–296. [Google Scholar]
- Sumiyoshi, H.; Hoke, C.H.; Trent, D.W. Infectious Japanese encephalitis virus RNA can be synthesized from in vitro-ligated cDNA templates. J. Virol. 1992, 66, 5425–5431. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, M.; Zhang, L.; Mohan, P.M.; Padmanabhan, R. Synthesis and characterization of an infectious dengue virus type-2 RNA genome (New Guinea C strain). Gene 1995, 162, 175–180. [Google Scholar] [CrossRef]
- Mandl, C.W.; Heinz, F.X.; Holzmann, H.; Kunz, C.; Ecker, M. Infectious cDNA clones of tick-borne encephalitis virus European subtype prototypic strain Neudoerfl and high virulence strain Hypr. J. Gen. Virol. 1997, 78, 1049–1057. [Google Scholar] [CrossRef]
- Lai, C.J.; Zhao, B.T.; Hori, H.; Bray, M. Infectious RNA transcribed from stably cloned full-length cDNA of dengue type 4 virus. Proc. Natl. Acad. Sci. USA 1991, 88, 5139–5143. [Google Scholar] [CrossRef] [Green Version]
- Khromykh, A.A.; Westaway, E.G. Completion of Kunjin virus RNA sequence and recovery of an infectious RNA transcribed from stably cloned full-length cDNA. J. Virol. 1994, 68, 4580–4588. [Google Scholar] [CrossRef] [Green Version]
- Hurrelbrink, R.J.; Nestorowicz, A.; McMinn, P.C. Characterization of infectious Murray Valley encephalitis virus derived from a stably cloned genome-length cDNA The GenBank accession number of the sequence reported in this paper is AF161266. J. Gen. Virol. 1999, 80, 3115–3125. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.S.; Pletnev, A.G. Infectious cDNA clones of Langat tick-borne flavivirus that differ from their parent in peripheral neurovirulence. Virology 2000, 269, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.-Y.; Tilgner, M.; Lo, M.K.; Kent, K.A.; Bernard, K.A. Infectious cDNA clone of the epidemic west Nile virus from New York City. J. Virol. 2002, 76, 5847–5856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Kamiyama, D.; Sekine, S.; Barsi-Rhyne, B.; Hu, J.; Chen, B.; Gilbert, L.A.; Ishikawa, H.; Leonetti, M.D.; Marshall, W.F.; Weissman, J.S.; et al. Versatile protein tagging in cells with split fluorescent protein. Nat. Commun. 2016, 7, 11046. [Google Scholar] [CrossRef]
- Dixon, A.S.; Schwinn, M.K.; Hall, M.P.; Zimmerman, K.; Otto, P.; Lubben, T.H.; Butler, B.L.; Binkowski, B.F.; Machleidt, T.; Kirkland, T.A.; et al. NanoLuc complementation reporter optimized for accurate measurement of protein interactions in cells. ACS Chem. Biol. 2015, 11, 400–408. [Google Scholar] [CrossRef]
- Puig-Basagoiti, F.; Deas, T.S.; Ren, P.; Tilgner, M.; Ferguson, D.M.; Shi, P.-Y. High-throughput assays using a luciferase-expressing replicon, virus-like particles, and full-length virus for west Nile virus drug discovery. Antimicrob. Agents Chemother. 2005, 49, 4980–4988. [Google Scholar] [CrossRef] [Green Version]
- Pierson, T.C.; Sánchez, M.D.; Puffer, B.A.; Ahmed, A.A.; Geiss, B.J.; Valentine, L.E.; Altamura, L.A.; Diamond, M.S.; Doms, R.W. A rapid and quantitative assay for measuring antibody-mediated neutralization of West Nile virus infection. Virology 2006, 346, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Shan, C.; Xie, X.; Ren, P.; Loeffelholz, M.J.; Yang, Y.; Furuya, A.; Dupuis, A.P.; Kramer, L.D.; Wong, S.J.; Shi, P.-Y. A rapid zika diagnostic assay to measure neutralizing antibodies in patients. Ebiomedicine 2017, 17, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Frumence, E.; Viranaicken, W.; Gadea, G.; Desprès, P. A GFP reporter MR766-based flow cytometry neutralization test for rapid detection of zika virus-neutralizing antibodies in serum specimens. Vaccines 2019, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; Yamanaka, A.; Yato, K.; Yoshii, K.; Watashi, K.; Aizaki, H.; Konishi, E.; Takasaki, T.; Kato, T.; Muramatsu, M.; et al. High-throughput neutralization assay for multiple flaviviruses based on single-round infectious particles using dengue virus type 1 reporter replicon. Sci. Rep. UK 2018, 8, 16624. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, A.; Moi, M.L.; Takasaki, T.; Kurane, I.; Matsuda, M.; Suzuki, R.; Konishi, E. Utility of Japanese encephalitis virus Subgenomic replicon-based single-round infectious particles as antigens in neutralization tests for Zika virus and three other flaviviruses. J. Virol. Methods 2017, 243, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, A.; Suzuki, R.; Konishi, E. Evaluation of single-round infectious, chimeric dengue type 1 virus as an antigen for dengue functional antibody assays. Vaccine 2014, 32, 4289–4295. [Google Scholar] [CrossRef] [PubMed]
- Bonaldo, M.C.; Garratt, R.C.; Caufour, P.S.; Freire, M.S.; Rodrigues, M.M.; Nussenzweig, R.S.; Galler, R. Surface expression of an immunodominant malaria protein B cell epitope by yellow fever virus 1 1Edited by J. Karn. J. Mol. Biol. 2002, 315, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Bonaldo, M.C.; Garratt, R.C.; Marchevsky, R.S.; Coutinho, E.S.F.; Jabor, A.V.; Almeida, L.F.C.; Yamamura, A.M.Y.; Duarte, A.S.; Oliveira, P.J.; Lizeu, J.O.P.; et al. Attenuation of recombinant yellow fever 17D viruses expressing foreign protein epitopes at the surface. J. Virol. 2005, 79, 8602–8613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barba-Spaeth, G.; Longman, R.S.; Albert, M.L.; Rice, C.M. Live attenuated yellow fever 17D infects human DCs and allows for presentation of endogenous and recombinant T cell epitopes. J. Exp. Med. 2005, 202, 1179–1184. [Google Scholar] [CrossRef]
- Tao, D.; Barba-Spaeth, G.; Rai, U.; Nussenzweig, V.; Rice, C.M.; Nussenzweig, R.S. Yellow fever 17D as a vaccine vector for microbial CTL epitopes. J. Exp. Med. 2005, 201, 201–209. [Google Scholar] [CrossRef] [Green Version]
- McAllister, A.; Arbetman, A.E.; Mandl, S.; Peña-Rossi, C.; Andino, R. Recombinant yellow fever viruses are effective therapeutic vaccines for treatment of murine experimental solid tumors and pulmonary metastases. J. Virol. 2000, 74, 9197–9205. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.A.; Wilson, N.A.; Piaskowski, S.M.; Weisgrau, K.L.; Furlott, J.R.; Bonaldo, M.C.; de Santana, M.G.V.; Rudersdorf, R.A.; Rakasz, E.G.; Keating, K.D.; et al. Vaccination with Gag, Vif, and Nef gene fragments affords partial control of viral replication after mucosal challenge with SIVmac239. J. Virol. 2014, 88, 7493–7516. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Year of Publication | Reporter Gene and Flavivirus | Reference |
|---|---|---|
| 1997 | CAT KUNV replicons | [14] |
| 1999 | GFP KUNV replicon | [15] |
| 2003 | IRES-GFP/Luc JEV | [16] |
| 2005 | IRES-GFP WNV | [17] |
| 2005 | IRES-RLuc WNV | [18] |
| 2007 | IRES-RLuc DENV2 | [19] |
| 2007 | E/NS1-GFP YFV | [20] |
| 2007 | 5′-GFP YFV | [21] |
| 2009 | 5′-RLuc DENV2 | [22,23] |
| 2010 | IRES-RLuc DENV2 | [24] |
| 2011 | 5′-RLuc DENV2 | [25] |
| 2012 | 5′-GFP/FLuc DENV2 | [26] |
| 2014 | E/NS1-gag YFV | [27] |
| 2016 | 5′-RLuc ZIKV | [28] |
| 2016 | 5′-GLuc WNV | [29] |
| 2016 | 5′-GFP ZIKV | [30] |
| 2017 | E/NS1-GFP LGTV | [31] |
| 2017 | 5′-GFP/mCherry/NanoLuc ZIKV | [32] |
| 2017 | 5′-RLuc JEV | [33] |
| 2017 | NS1-HiBiT JEV/DENV4 | [34] |
| 2018 | 5′-GFP/Clover2/bfloGFP DENV2 | [35] |
| 2019 | 5′ NanoLuc DTMUV | [36] |
| 2019 | NS1-HiBiT JEV | [37] |
| 2020 | IRES-NanoLuc/GFP ZIKV | [38] |
| 2020 | E/NS1-NanoLuc/GFP ZIKV | [39] |
| 2020 | 5′-GFP/NanoLuc ZIKV | [39] |
| 2020 | 5′ w/+1C-1 NanoLuc/GFP ZIKV | [39] |
| 2020 | 5′ w/ΔC-NanoLuc ZIKV/YFV | [40] |
| 2020 | 5′ C33-38 NanoLuc ZIKV/YFV/DENV1-4/JEV/WNV | [41] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baker, C.; Shi, P.-Y. Construction of Stable Reporter Flaviviruses and Their Applications. Viruses 2020, 12, 1082. https://doi.org/10.3390/v12101082
Baker C, Shi P-Y. Construction of Stable Reporter Flaviviruses and Their Applications. Viruses. 2020; 12(10):1082. https://doi.org/10.3390/v12101082
Chicago/Turabian StyleBaker, Coleman, and Pei-Yong Shi. 2020. "Construction of Stable Reporter Flaviviruses and Their Applications" Viruses 12, no. 10: 1082. https://doi.org/10.3390/v12101082
APA StyleBaker, C., & Shi, P. -Y. (2020). Construction of Stable Reporter Flaviviruses and Their Applications. Viruses, 12(10), 1082. https://doi.org/10.3390/v12101082