A Nationwide Study about the Dispersal Patterns of the Predominant HIV-1 Subtypes A1 and B in Greece: Inference of the Molecular Transmission Clusters

 ,
, 
 ,
,  , , , , , , , ,
, , , , , , , , 
 ,
, 
 add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Subtyping Analysis
2.3. Phylogenetic Analysis
2.4. Phylogeographic Analysis
2.5. Statistical Analysis
2.6. Sequences Availability
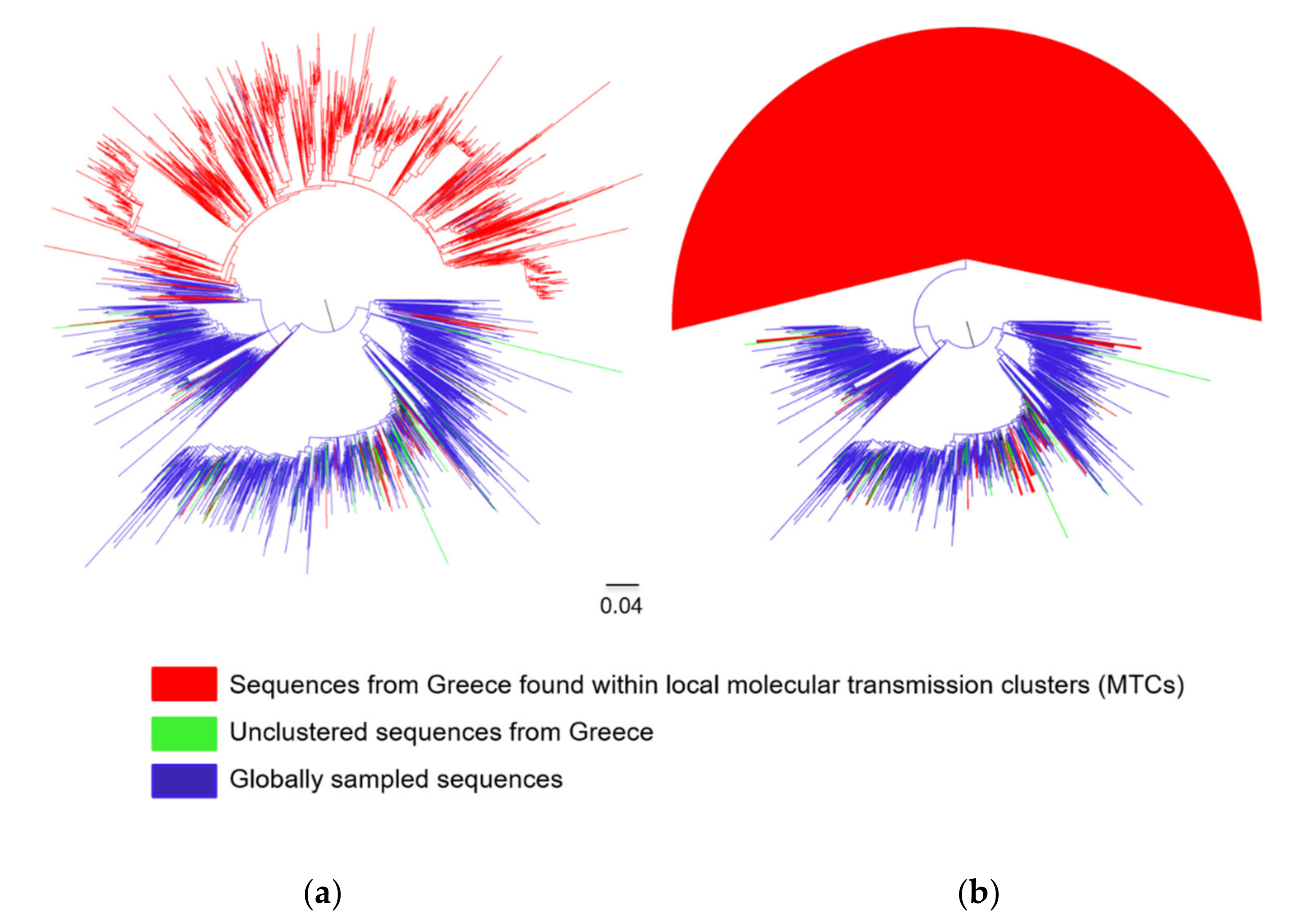
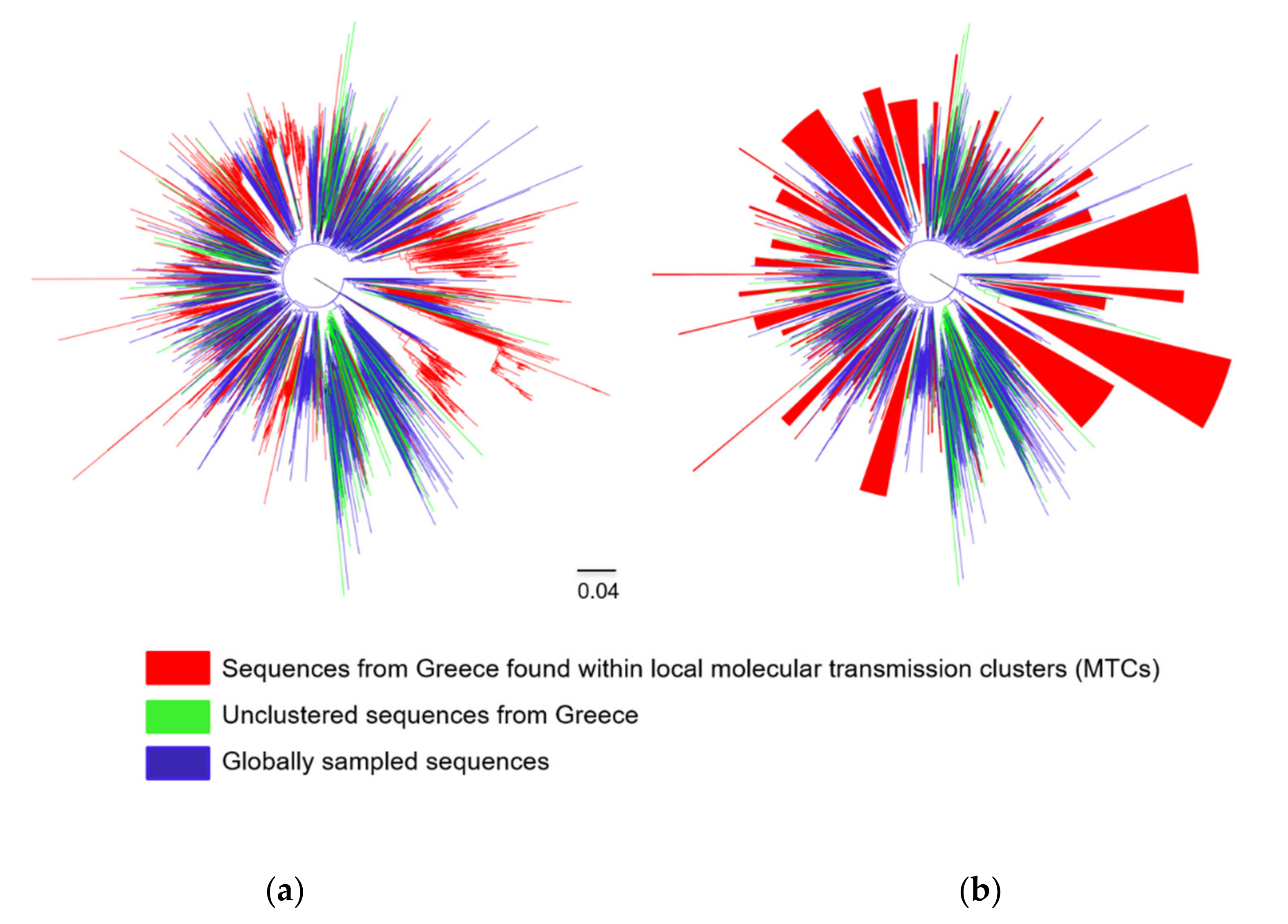
3. Results
3.1. Subtyping Results
3.2. Subtypes A1 and B Dispersal Patterns and Regional Transmission
3.3. Characteristics of Transmissions Within and Outside MTCs
3.4. Factors Associated with Local Transmission of Subtypes A1 and B
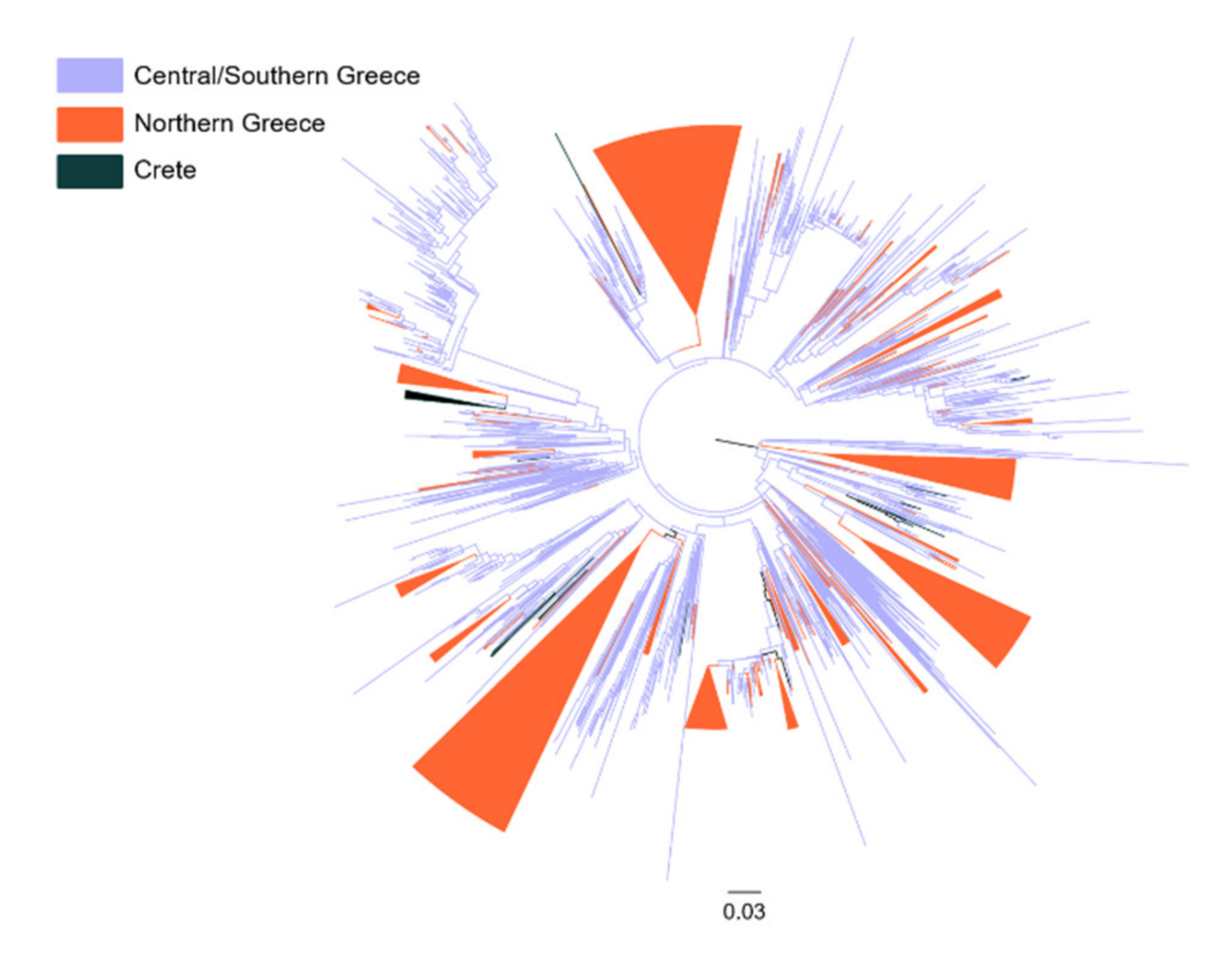
3.5. Patterns of Geographical Dispersal
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foley, B.T.; Leitner, T.; Paraskevis, D.; Peeters, M. Primate immunodeficiency virus classification and nomenclature: Review. Infect. Genet. Evol. 2016, 46, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, J.; Vallari, A.; McArthur, C.; Sthreshley, L.; Cloherty, G.A.; Berg, M.G.; Rodgers, M.A. Complete Genome Sequence of CG-0018a-01 Establishes HIV-1 Subtype L. J. Acquir Immune Defic. Syndr. 2020, 83, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Hemelaar, J.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Fleminger, I.; Kirtley, S.; Williams, B.; Gouws-Williams, E.; Ghys, P.D.; Abimiku, A.l.G.; et al. Global and regional molecular epidemiology of HIV-1, 1990–2015: A systematic review, global survey, and trend analysis. Lancet Infect. Dis. 2019, 19, 143–155. [Google Scholar] [CrossRef]
- Abeler-Dorner, L.; Grabowski, M.K.; Rambaut, A.; Pillay, D.; Fraser, C.; PANGEA Consortium. PANGEA-HIV 2: Phylogenetics And Networks for Generalised Epidemics in Africa. Curr. Opin. HIV AIDS 2019, 14, 173–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Vijver, D.; Boucher, C.A.B. Insights on transmission of HIV from phylogenetic analysis to locally optimize HIV prevention strategies. Curr. Opin. HIV AIDS 2018, 13, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B.G.; Ibanescu, R.I.; Hardy, I.; Roger, M. Genotypic and Phylogenetic Insights on Prevention of the Spread of HIV-1 and Drug Resistance in “Real-World” Settings. Viruses 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Chaillon, A.; Essat, A.; Frange, P.; Smith, D.M.; Delaugerre, C.; Barin, F.; Ghosn, J.; Pialoux, G.; Robineau, O.; Rouzioux, C.; et al. Spatiotemporal dynamics of HIV-1 transmission in France (1999–2014) and impact of targeted prevention strategies. Retrovirology 2017, 14, 15. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, M.K.; Lessler, J. Phylogenetic insights into age-disparate partnerships and HIV. Lancet HIV 2017, 4, e8–e9. [Google Scholar] [CrossRef]
- Bezemer, D.; Cori, A.; Ratmann, O.; van Sighem, A.; Hermanides, H.S.; Dutilh, B.E.; Gras, L.; Rodrigues Faria, N.; van den Hengel, R.; Duits, A.J.; et al. Dispersion of the HIV-1 Epidemic in Men Who Have Sex with Men in the Netherlands: A Combined Mathematical Model and Phylogenetic Analysis. PLoS Med. 2015, 12, e1001898. [Google Scholar] [CrossRef] [Green Version]
- Vasylyeva, T.I.; Zarebski, A.; Smyrnov, P.; Williams, L.D.; Korobchuk, A.; Liulchuk, M.; Zadorozhna, V.; Nikolopoulos, G.; Paraskevis, D.; Schneider, J.; et al. Phylodynamics Helps to Evaluate the Impact of an HIV Prevention Intervention. Viruses 2020, 12, 469. [Google Scholar] [CrossRef] [Green Version]
- Paraskevis, D.; Nikolopoulos, G.K.; Magiorkinis, G.; Hodges-Mameletzis, I.; Hatzakis, A. The application of HIV molecular epidemiology to public health. Infect. Genet. Evol. 2016, 46, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B.G.; Wainberg, M.A. Future of phylogeny in HIV prevention. J. Acquir. Immune Defic. Syndr. 2013, 63, S248–S254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivay, M.V.; Grabowski, M.K.; Zhang, Y.; Palumbo, P.J.; Guo, X.; Piwowar-Manning, E.; Hamilton, E.L.; Viet Ha, T.; Antonyak, S.; Imran, D.; et al. Phylogenetic Analysis of Human Immunodeficiency Virus from People Who Inject Drugs in Indonesia, Ukraine, and Vietnam: HPTN 074. Clin. Infect. Dis. 2019. [Google Scholar] [CrossRef]
- Alpren, C.; Dawson, E.L.; John, B.; Cranston, K.; Panneer, N.; Fukuda, H.D.; Roosevelt, K.; Klevens, R.M.; Bryant, J.; Peters, P.J.; et al. Opioid Use Fueling HIV Transmission in an Urban Setting: An Outbreak of HIV Infection Among People Who Inject Drugs-Massachusetts, 2015–2018. Am. J. Public Health 2020, 110, 37–44. [Google Scholar] [CrossRef]
- Kostaki, E.G.; Frampton, D.; Paraskevis, D.; Pantavou, K.; Ferns, B.; Raffle, J.; Grant, P.; Kozlakidis, Z.; Hadjikou, A.; Pavlitina, E.; et al. Near Full-length Genomic Sequencing and Molecular Analysis of HIV-Infected Individuals in a Network-based Intervention (TRIP) in Athens, Greece: Evidence that Transmissions Occur More Frequently from those with High HIV-RNA. Curr. HIV Res. 2018, 16, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Peters, P.J.; Pontones, P.; Hoover, K.W.; Patel, M.R.; Galang, R.R.; Shields, J.; Blosser, S.J.; Spiller, M.W.; Combs, B.; Switzer, W.M.; et al. HIV Infection Linked to Injection Use of Oxymorphone in Indiana, 2014–2015. N. Engl. J. Med. 2016, 375, 229–239. [Google Scholar] [CrossRef] [Green Version]
- CDC: Detecting and Responding to HIV Transmission Clusters. A Guide for Health Departments. Available online: https://www.cdc.gov/hiv/pdf/funding/announcements/ps18-1802/CDC-HIV-PS18-1802-AttachmentE-Detecting-Investigating-and-Responding-to-HIV-Transmission-Clusters.pdf (accessed on 19 September 2020).
- Paraskevis, D.; Beloukas, A.; Stasinos, K.; Pantazis, N.; de Mendoza, C.; Bannert, N.; Meyer, L.; Zangerle, R.; Gill, J.; Prins, M.; et al. HIV-1 molecular transmission clusters in nine European countries and Canada: Association with demographic and clinical factors. BMC Med. 2019, 17, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paraskevis, D.; Magiorkinis, E.; Magiorkinis, G.; Sypsa, V.; Paparizos, V.; Lazanas, M.; Gargalianos, P.; Antoniadou, A.; Panos, G.; Chrysos, G.; et al. Increasing prevalence of HIV-1 subtype A in Greece: Estimating epidemic history and origin. J. Infect. Dis. 2007, 196, 1167–1176. [Google Scholar] [CrossRef] [Green Version]
- Paraskevis, D.; Paraschiv, S.; Sypsa, V.; Nikolopoulos, G.; Tsiara, C.; Magiorkinis, G.; Psichogiou, M.; Flampouris, A.; Mardarescu, M.; Niculescu, I.; et al. Enhanced HIV-1 surveillance using molecular epidemiology to study and monitor HIV-1 outbreaks among intravenous drug users (IDUs) in Athens and Bucharest. Infect. Genet. Evol. 2015, 35, 109–121. [Google Scholar] [CrossRef]
- Paraskevis, D.; Kostaki, E.; Magiorkinis, G.; Gargalianos, P.; Xylomenos, G.; Magiorkinis, E.; Lazanas, M.; Chini, M.; Nikolopoulos, G.; Skoutelis, A.; et al. Prevalence of drug resistance among HIV-1 treatment-naive patients in Greece during 2003–2015: Transmitted drug resistance is due to onward transmissions. Infect. Genet. Evol. 2017, 54, 183–191. [Google Scholar] [CrossRef]
- NPHO: HIV/AIDS Surveillance in Greece Diagnoses through 31/12/2019. Available online: https://eody.gov.gr/wp-content/uploads/2018/12/epidimiologiko-deltio-hiv-2019.pdf (accessed on 19 September 2020).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-Length Human Immunodeficiency Virus Type 1 Genomes from Subtype C-Infected Seroconverters in India, with Evidence of Intersubtype Recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Kostaki, E.G.; Flampouris, A.; Karamitros, T.; Chueca, N.; Alvarez, M.; Casas, P.; Alejos, B.; Hatzakis, A.; Garcia, F.; Paraskevis, D.; et al. Spatiotemporal Characteristics of the Largest HIV-1 CRF02_AG Outbreak in Spain: Evidence for Onward Transmissions. Front. Microbiol. 2019, 10, 370. [Google Scholar] [CrossRef] [PubMed]
- Von Wyl, V.; Kouyos, R.D.; Yerly, S.; Boni, J.; Shah, C.; Burgisser, P.; Klimkait, T.; Weber, R.; Hirschel, B.; Cavassini, M.; et al. The role of migration and domestic transmission in the spread of HIV-1 non-B subtypes in Switzerland. J. Infect. Dis. 2011, 204, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.; Domelevo Entfellner, J.B.; Wilkinson, E.; Correia, D.; Davila Felipe, M.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.61. Available online: http://www.mesquiteproject.org (accessed on 19 September 2020).
- Wilgenbusch, J.C.; Swofford, D. Inferring evolutionary trees with PAUP*. Curr. Protoc. Bioinform. 2003. [Google Scholar] [CrossRef]
- Angelis, K.; Albert, J.; Mamais, I.; Magiorkinis, G.; Hatzakis, A.; Hamouda, O.; Struck, D.; Vercauteren, J.; Wensing, A.M.; Alexiev, I.; et al. Global Dispersal Pattern of HIV Type 1 Subtype CRF01_AE: A Genetic Trace of Human Mobility Related to Heterosexual Sexual Activities Centralized in Southeast Asia. J. Infect. Dis. 2015, 211, 1735–1744. [Google Scholar] [CrossRef] [PubMed]
- Magiorkinis, G.; Angelis, K.; Mamais, I.; Katzourakis, A.; Hatzakis, A.; Albert, J.; Lawyer, G.; Hamouda, O.; Struck, D.; Vercauteren, J.; et al. The global spread of HIV-1 subtype B epidemic. Infect. Genet. Evol. 2016, 46, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Paraskevis, D.; Kostaki, E.; Nikolopoulos, G.K.; Sypsa, V.; Psichogiou, M.; Del Amo, J.; Hodges-Mameletzis, I.; Paraskeva, D.; Skoutelis, A.; Malliori, M.; et al. Molecular Tracing of the Geographical Origin of Human Immunodeficiency Virus Type 1 Infection and Patterns of Epidemic Spread Among Migrants Who Inject Drugs in Athens. Clin. Infect. Dis. 2017, 65, 2078–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabeni, L.; Santoro, M.M.; Lorenzini, P.; Rusconi, S.; Gianotti, N.; Costantini, A.; Sarmati, L.; Antinori, A.; Ceccherini-Silberstein, F.; d’Arminio Monforte, A.; et al. Evaluation of HIV Transmission Clusters among Natives and Foreigners Living in Italy. Viruses 2020, 12, 791. [Google Scholar] [CrossRef] [PubMed]
- Verhofstede, C.; Mortier, V.; Dauwe, K.; Callens, S.; Deblonde, J.; Dessilly, G.; Delforge, M.L.; Fransen, K.; Sasse, A.; Stoffels, K.; et al. Exploring HIV-1 Transmission Dynamics by Combining Phylogenetic Analysis and Infection Timing. Viruses 2019, 11, 1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostrikis, L.G.; Hezka, J.; Stylianou, D.C.; Kostaki, E.; Andreou, M.; Kousiappa, I.; Paraskevis, D.; Demetriades, I. HIV-1 transmission networks across Cyprus (2010–2012). PLoS ONE 2018, 13, e0195660. [Google Scholar] [CrossRef] [Green Version]
- Lewis, F.; Hughes, G.J.; Rambaut, A.; Pozniak, A.; Brown, A.J.L. Episodic Sexual Transmission of HIV Revealed by Molecular Phylodynamics. PLoS Med. 2008, 5, e50. [Google Scholar] [CrossRef] [Green Version]
- Beloukas, A.; Psarris, A.; Giannelou, P.; Kostaki, E.; Hatzakis, A.; Paraskevis, D. Molecular epidemiology of HIV-1 infection in Europe: An overview. Infect. Genet. Evol. 2016, 46, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Ciccozzi, M.; Gori, C.; Boros, S.; Ruiz-Alvarez, M.J.; Harxhi, A.; Dervishi, M.; Qyra, S.; Schinaia, N.; D’Arrigo, R.; Ceccherini-Silberstein, F.; et al. Molecular Diversity of HIV in Albania. J. Infect. Dis. 2005, 192, 475–479. [Google Scholar] [CrossRef] [Green Version]
- Aibekova, L.; Foley, B.; Hortelano, G.; Raees, M.; Abdraimov, S.; Toichuev, R.; Ali, S. Molecular epidemiology of HIV-1 subtype A in former Soviet Union countries. PLoS ONE 2018, 13, e0191891. [Google Scholar] [CrossRef]
- Schlosser, M.; Kartashev, V.V.; Mikkola, V.H.; Shemshura, A.; Saukhat, S.; Kolpakov, D.; Suladze, A.; Tverdokhlebova, T.; Hutt, K.; Heger, E.; et al. HIV-1 Sub-Subtype A6: Settings for Normalised Identification and Molecular Epidemiology in the Southern Federal District, Russia. Viruses 2020, 12, 475. [Google Scholar] [CrossRef] [Green Version]
- Bobkov, A.; Cheingsong-Popov, R.; Selimova, L.; Ladnaya, N.; Kazennova, E.; Kravchenko, A.; Fedotov, E.; Saukhat, S.; Zverev, S.; Pokrovsky, V.; et al. An HIV Type 1 Epidemic among Injecting Drug Users in the Former Soviet Union Caused by a Homogeneous Subtype A Strain. AIDS Res. Hum. Retrovir. 1997, 13, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.M.; de Parga, E.V.; Vinogradova, A.; Sierra, M.; Yakovlev, A.; Rakhmanova, A.; Delgado, E.; Casado, G.; Munoz, M.; Carmona, R.; et al. New insights into the origin of the HIV type 1 subtype A epidemic in former Soviet Union’s countries derived from sequence analyses of preepidemically transmitted viruses. AIDS Res. Hum. Retrovir. 2007, 23, 1599–1604. [Google Scholar] [CrossRef]
- Diez-Fuertes, F.; Cabello, M.; Thomson, M.M. Bayesian phylogeographic analyses clarify the origin of the HIV-1 subtype A variant circulating in former Soviet Union’s countries. Infect. Genet. Evol. 2015, 33, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Araujo, P.M.M.; Carvalho, A.; Pingarilho, M.; Abecasis, A.B.; Osorio, N.S. Characterization of a large cluster of HIV-1 A1 infections detected in Portugal and connected to several Western European countries. Sci. Rep. 2019, 9, 7223. [Google Scholar] [CrossRef] [Green Version]
- Abecasis, A.B.; Wensing, A.M.J.; Paraskevis, D.; Vercauteren, J.; Theys, K.; Van de Vijver, D.A.M.C.; Albert, J.; Asjö, B.; Balotta, C.; Beshkov, D.; et al. HIV-1 subtype distribution and its demographic determinants in newly diagnosed patients in Europe suggest highly compartmentalized epidemics. Retrovirology 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Aldous, J.L.; Pond, S.K.; Poon, A.; Jain, S.; Qin, H.; Kahn, J.S.; Kitahata, M.; Rodriguez, B.; Dennis, A.M.; Boswell, S.L.; et al. Characterizing HIV transmission networks across the United States. Clin. Infect. Dis. 2012, 55, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Drescher, S.M.; von Wyl, V.; Yang, W.L.; Boni, J.; Yerly, S.; Shah, C.; Aubert, V.; Klimkait, T.; Taffe, P.; Furrer, H.; et al. Treatment-naive individuals are the major source of transmitted HIV-1 drug resistance in men who have sex with men in the Swiss HIV Cohort Study. Clin. Infect. Dis. 2014, 58, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostaki, E.G.; Soulie, C.; Visseaux, B.; Storto, A.; Charpentier, C.; Wirden, M.; Landman, R.; Katlama, C.; Calvez, V.; Descamps, D.; et al. Molecular Analysis Suggests Post-Migration HIV-1 Acquisition among Migrants in Paris. In Proceedings of the Virtual Conference on Retroviruses and Opportunistic Infections, Boston, MA, USA, 8–11 March 2020; Abstract Number 921. Available online: https://www.croiconference.org/abstract/molecular-analysis-suggests-post-migration-hiv-1-acquisition-among-migrants-in-paris/ (accessed on 19 September 2020).
- Skoura, L.; Metallidis, S.; Buckton, A.J.; Mbisa, J.L.; Pilalas, D.; Papadimitriou, E.; Papoutsi, A.; Haidich, A.B.; Chrysanthidis, T.; Tsachouridou, O.; et al. Molecular and epidemiological characterization of HIV-1 infection networks involving transmitted drug resistance mutations in Northern Greece. J. Antimicrob. Chemother. 2011, 66, 2831–2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabeni, L.; Alteri, C.; Di Carlo, D.; Orchi, N.; Carioti, L.; Bertoli, A.; Gori, C.; Forbici, F.; Continenza, F.; Maffongelli, G.; et al. Dynamics and phylogenetic relationships of HIV-1 transmitted drug resistance according to subtype in Italy over the years 2000-14. J. Antimicrob. Chemother. 2017, 72, 2837–2845. [Google Scholar] [CrossRef]
- Rhee, S.Y.; Blanco, J.L.; Jordan, M.R.; Taylor, J.; Lemey, P.; Varghese, V.; Hamers, R.L.; Bertagnolio, S.; Rinke de Wit, T.F.; Aghokeng, A.F.; et al. Geographic and temporal trends in the molecular epidemiology and genetic mechanisms of transmitted HIV-1 drug resistance: An individual-patient- and sequence-level meta-analysis. PLoS Med. 2015, 12, e1001810. [Google Scholar] [CrossRef] [Green Version]
- Kleyn, T.J.; Liedtke, M.D.; Harrison, D.L.; Lockhart, S.M.; Salvaggio, M.R.; Ripley, T.L.; Rathbun, R.C. Incidence of transmitted antiretroviral drug resistance in treatment-naive HIV-1-infected persons in a large South Central United States clinic. Ann. Pharm. 2014, 48, 470–475. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Subtype A1 | Subtype B |
|---|---|---|
| Age, years, median (IQR) | 37 (30–46) 1 | 36 (30–44) 2 |
| Gender, n (%) | ||
| Male | 1.435 (81.9) | 2217 (86.1) |
| Female | 232 (13.3) | 282 (10.9) |
| Unknown | 84 (4.8) | 76 (3.0) |
| Transmission risk group, n (%) | ||
| MSM | 1035 (59.1) | 1718 (66.7) |
| PWID | 150 (8.6) | 182 (7.1) |
| Heterosexuals | 387 (22.1) | 438 (17.0) |
| Other | 20 (1.1) | 88 (3.4) |
| Unknown | 159 (9.1) | 149 (5.8) |
| Origin, n (%) | ||
| Greek | 1321 (75.4) | 2117 (82.2) |
| Non-Greek | 197 (11.3) | 186 (7.2) |
| Unknown | 233 (13.3) | 272 (10.6) |
| Sampling period | ||
| 1999–2005 | 365 (20.8) | 1020 (39.6) |
| 2006–2010 | 503 (28.7) | 618 (24.0) |
| 2011–2015 | 880 (50.3) | 934 (36.3) |
| Unknown | 3 (0.2) | 3 (0.1) |
| Total, n, (%) | 1751 (100.0) | 2575 (100.0) |
| Subtype A1 | Subtype B | |||
|---|---|---|---|---|
| Characteristic | Within MTCs | Outside MTCs | Within MTCs | Outside MTCs |
| Age, years, median (IQR) | 37 (29–46) 1 | 37 (31–48) 2 | 35 (29–42) 3 | 40 (34–48) 4 |
| Gender, n (%) | ||||
| Male | 1373 (83.6) | 62 (56.9) | 1729 (87.2) | 488 (82.3) |
| Female | 193 (11.8) | 39 (35.8) | 196 (9.9) | 86 (14.5) |
| Unknown | 76 (4.6) | 8 (7.3) | 57 (2.9) | 19 (3.2) |
| Transmission risk group, n (%) | ||||
| MSM | 1022 (62.2) | 13 (11.9) | 1373 (69.3) | 345 (58.2) |
| PWID | 124 (7.6) | 26 (23.9) | 148 (7.5) | 34 (5.7) |
| Heterosexuals | 339 (20.6) | 48 (44.0) | 321 (16.2) | 117 (19.7) |
| Other | 18 (1.1) | 2 (1.8) | 23 (1.1) | 65 (11.0) |
| Unknown | 139 (8.5) | 20 (18.4) | 117 (5.9) | 32 (5.4) |
| Origin, n (%) | ||||
| Greek | 1283 (78.1) | 38 (34.9) | 1635 (82.5) | 482 (81.3) |
| Non-Greek | 150 (9.2) | 47 (43.1) | 139 (7.0) | 47 (7.9) |
| Unknown | 209 (12.7) | 24 (22.0) | 208 (10.5) | 64 (10.8) |
| Sampling period | ||||
| [1999–2005] | 350 (21.3) | 15 (13.7) | 647 (32.6) | 373 (62.9) |
| [2006–2010] | 459 (28.0) | 44 (40.4) | 510 (25.7) | 108 (18.2) |
| [2011–2015] | 830 (50.5) | 50 (45.9) | 822 (41.5) | 112 (18.9) |
| Unknown | 3 (0.2) | 0 (0.0) | 3 (0.2) | 0 (0.0) |
| Total, n, (%) | 1642 (100.0) | 109 (100.0) | 1982 (100.0) | 593 (100.0) |
| Subtype A1 1 | Subtype B 1 | |||||
|---|---|---|---|---|---|---|
| Explanatory Variable | Odds Ratio | 95% CI | p-Value | Odds Ratio | 95% CI | p-Value |
| Age | 0.99 | 0.97–1.01 | 0.341 | 0.96 | 0.95–0.97 | <0.001 |
| Gender | ||||||
| Male | 1.52 | 0.87–2.67 | 0.140 | 1.33 | 0.91–1.97 | 0.141 |
| Female 2 | ||||||
| Unknown | 2.80 | 0.64–12.3 | 0.172 | 0.84 | 0.22–3.16 | 0.796 |
| Transmission risk group | ||||||
| MSM | 5.34 | 2.52–11.31 | <0.001 | 0.98 | 0.69–1.38 | 0.893 |
| PWID | 0.42 | 0.22–0.80 | 0.008 | 0.80 | 0.49–1.30 | 0.364 |
| Heterosexuals 2 | ||||||
| Other | 0.86 | 0.17–4.47 | 0.861 | 0.10 | 0.05–0.18 | <0.001 |
| Unknown | 0.81 | 0.37–1.80 | 0.610 | 0.96 | 0.48–1.95 | 0.919 |
| Origin | ||||||
| Greek | 6.05 | 3.62–10.10 | <0.001 | 1.57 | 1.08–2.29 | 0.019 |
| Non-Greek 2 | ||||||
| Unknown | 1.27 | 0.62–2.60 | 0.516 | 1.26 | 0.74–2.12 | 0.394 |
| Sampling period | ||||||
| [1999–2005] 2 | ||||||
| [2006–2010] | 0.51 | 0.25–1.01 | 0.054 | 2.33 | 1.80–3.02 | <0.001 |
| [2011–2015] | 0.75 | 0.38–1.48 | 0.411 | 3.83 | 2.95–4.96 | <0.001 |
| Unknown | 1 | - | - | 1 | - | - |
| Sampling area | ||||||
| Central/Southern Greece 2 | ||||||
| Northern Greece | 0.91 | 0.54–1.55 | 0.736 | 1.32 | 0.96–1.81 | 0.092 |
| Crete | 1 | - | - | 1 | - | - |
| Subtype A1 | |||
|---|---|---|---|
| Importing to | |||
| Exporting from | Central/Southern Greece | Northern Greece | Crete |
| Central/Southern Greece | 90.9 | 11.6 | |
| Northern Greece | 35.41 | 0.2 | |
| Crete | 0.11 | 0 | |
| Subtype B | |||
| Central/Southern Greece | 67.6 | 15.0 | |
| Northern Greece | 17.41 | 1.41 | |
| Crete | 1.21 | 0.51 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostaki, E.G.; Gova, M.; Adamis, G.; Xylomenos, G.; Chini, M.; Mangafas, N.; Lazanas, M.; Metallidis, S.; Tsachouridou, O.; Papastamopoulos, V.; et al. A Nationwide Study about the Dispersal Patterns of the Predominant HIV-1 Subtypes A1 and B in Greece: Inference of the Molecular Transmission Clusters. Viruses 2020, 12, 1183. https://doi.org/10.3390/v12101183
Kostaki EG, Gova M, Adamis G, Xylomenos G, Chini M, Mangafas N, Lazanas M, Metallidis S, Tsachouridou O, Papastamopoulos V, et al. A Nationwide Study about the Dispersal Patterns of the Predominant HIV-1 Subtypes A1 and B in Greece: Inference of the Molecular Transmission Clusters. Viruses. 2020; 12(10):1183. https://doi.org/10.3390/v12101183
Chicago/Turabian StyleKostaki, Evangelia Georgia, Maria Gova, Georgios Adamis, Georgios Xylomenos, Maria Chini, Nikos Mangafas, Marios Lazanas, Simeon Metallidis, Olga Tsachouridou, Vasileios Papastamopoulos, and et al. 2020. "A Nationwide Study about the Dispersal Patterns of the Predominant HIV-1 Subtypes A1 and B in Greece: Inference of the Molecular Transmission Clusters" Viruses 12, no. 10: 1183. https://doi.org/10.3390/v12101183
APA StyleKostaki, E. G., Gova, M., Adamis, G., Xylomenos, G., Chini, M., Mangafas, N., Lazanas, M., Metallidis, S., Tsachouridou, O., Papastamopoulos, V., Chatzidimitriou, D., Kakalou, E., Antoniadou, A., Papadopoulos, A., Psichogiou, M., Basoulis, D., Pilalas, D., Papageorgiou, I., Paraskeva, D., ... Paraskevis, D. (2020). A Nationwide Study about the Dispersal Patterns of the Predominant HIV-1 Subtypes A1 and B in Greece: Inference of the Molecular Transmission Clusters. Viruses, 12(10), 1183. https://doi.org/10.3390/v12101183