The Cell Tropism of Porcine Respiratory Coronavirus for Airway Epithelial Cells Is Determined by the Expression of Porcine Aminopeptidase N

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Porcine Airway Epithelial Cell Cultures
2.2. Cell and Virus
2.3. Virus Infection of Differentiated Epithelial Cells
2.4. Immunohistochemistry Staining (IHC) and Immunofluorescence Analysis (IFA)
2.5. Virus Titration
2.6. Statistical Analyses
3. Results
3.1. Replication of PRCoV on wdPTECs
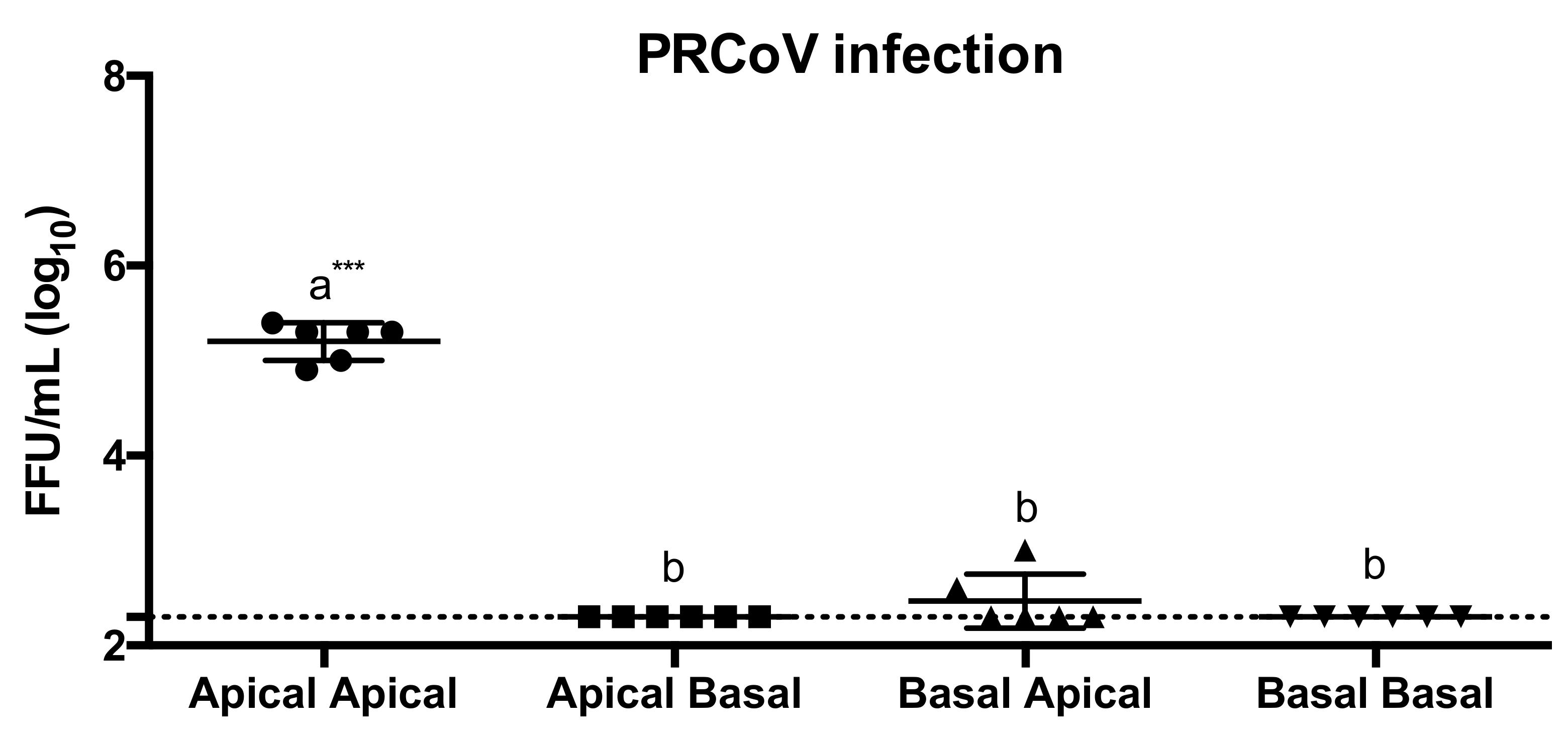
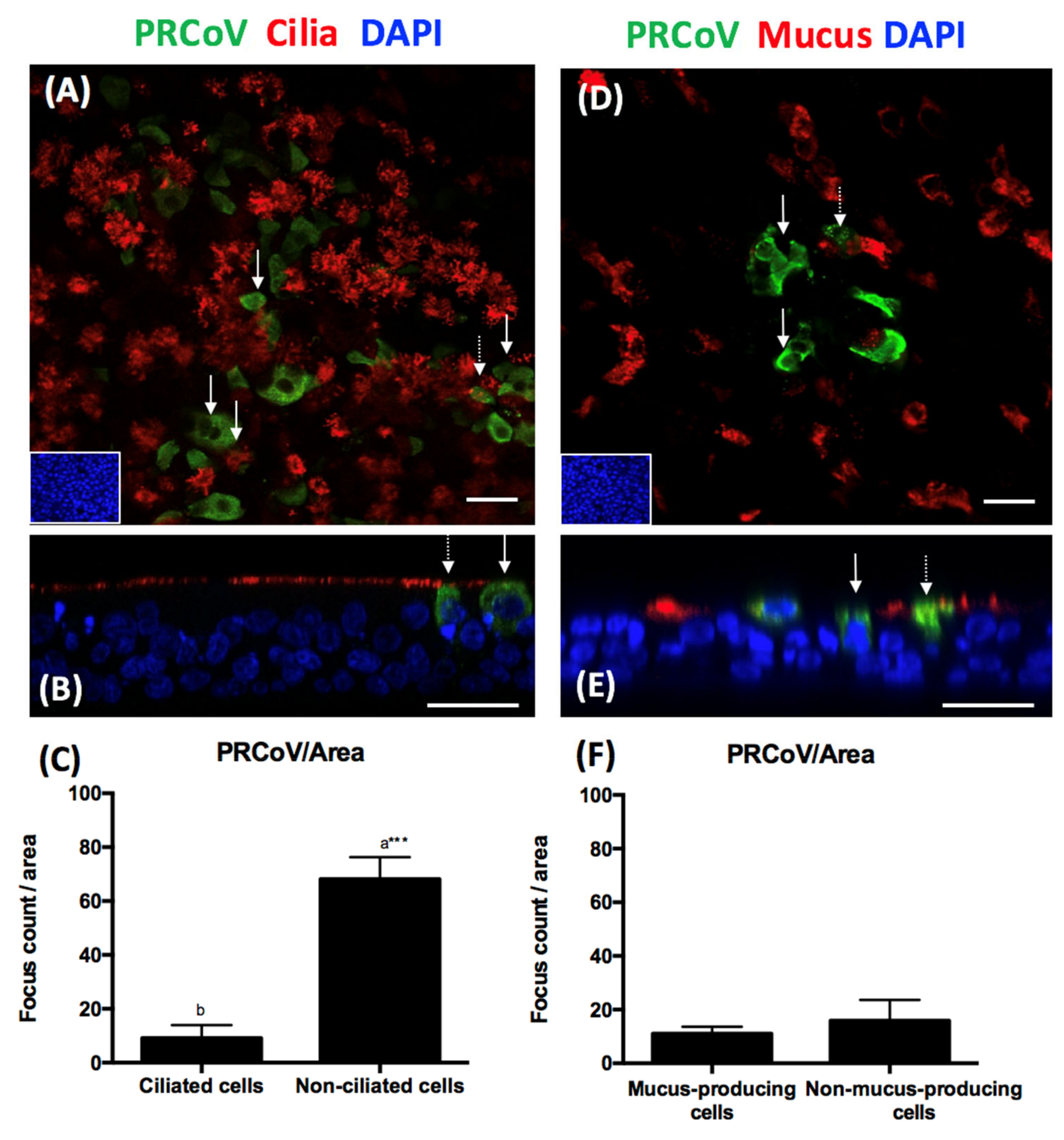
3.2. Cell Tropism of PRCoV
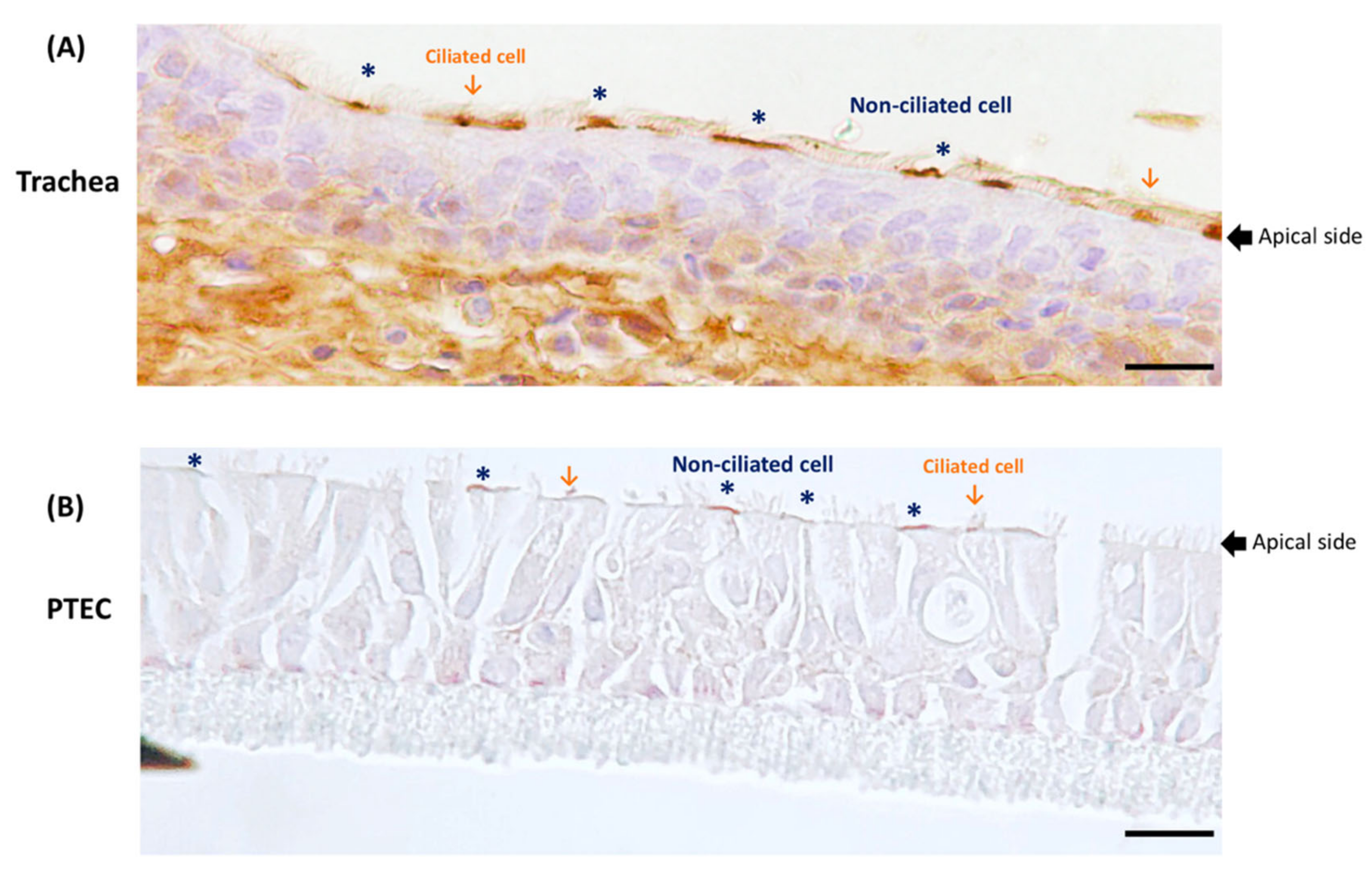
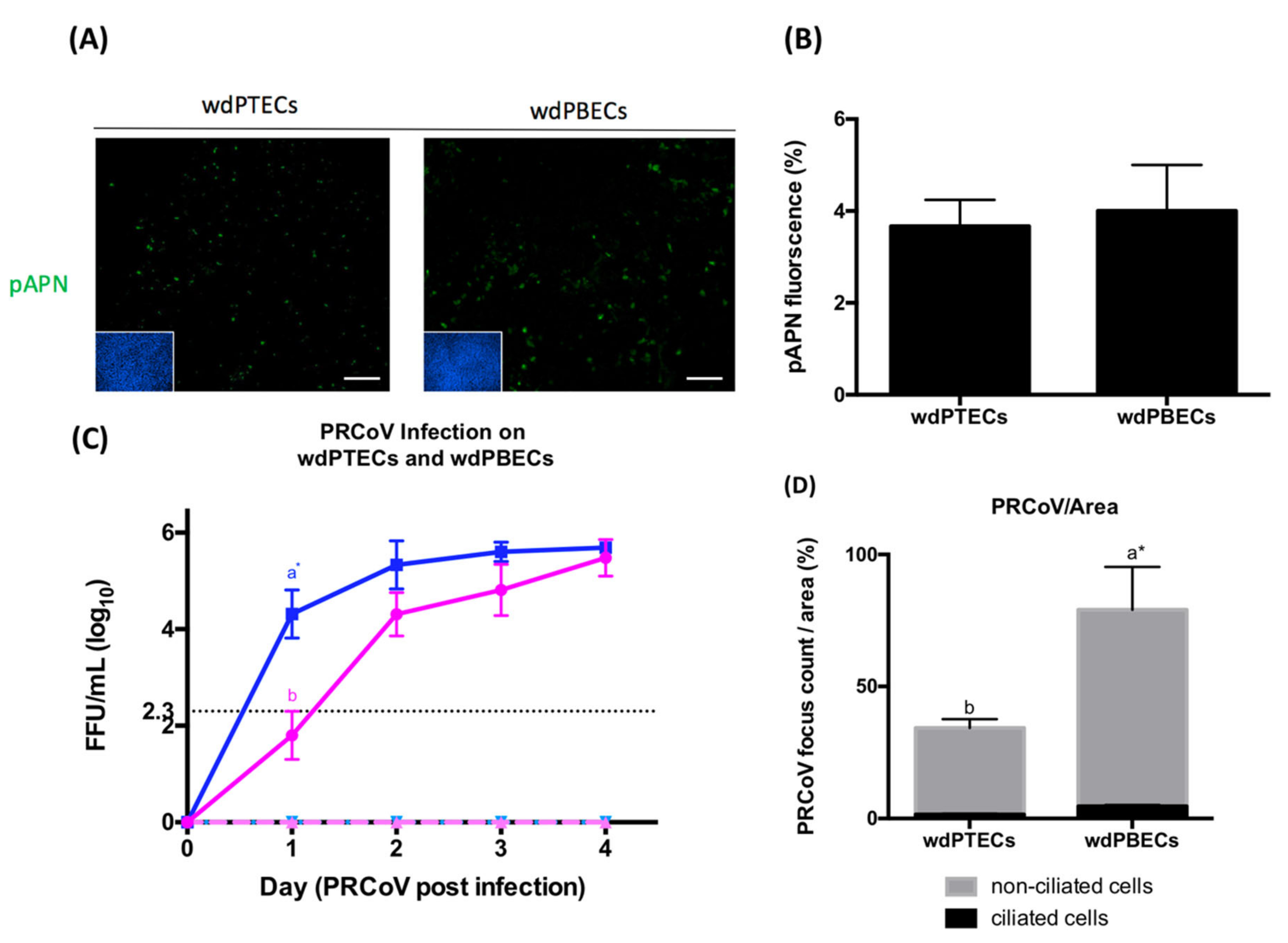
3.3. Expression of pAPN on the Porcine Trachea and Well-Differentiated Porcine Tracheal Epithelial Cells
3.4. Susceptibility to Infection and Expression of APN are Differentiation-Dependent
3.5. Infection in Bronchial Cells Is More Efficient Than It Is in Tracheal Cells
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saif, L.J. Animal coronaviruses: What can they teach us about the severe acute respiratory syndrome? Rev. Sci. Tech. OIE 2004, 23, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; Hayes, J.; Lewis, P.; Parwani, A.V.; Chang, K.O.; Saif, L.J. Molecular characterization and pathogenesis of transmissible gastroenteritis coronavirus (TGEV) and porcine respiratory coronavirus (PRCV) field isolates co-circulating in a swine herd. Arch Virol. 2000, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, A.; Fukuda, M.; Kuga, K.; Takagi, M.; Tsunemitsu, H. Prevalence of Antibodies against Transmissible Gastroenteritis Virus and Porcine Respiratory Coronavirus among Pigs in Six Regions in Japan. J. Vet. Med. Sci. 2010, 72, 943–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groschup, M.H.; Brun, A.; Haas, B. Serological Studies on the Potential Synergism of Porcine Reproductive and Respiratory Syndrome Virus and Influenza, Corona and Paramyxoviruses in the Induction of Respiratory Symptoms in Swine. J. Vet. Med. Ser. B 1993, 40, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Pensaert, M.; Callebaut, P.; Vergote, J. Isolation of a Porcine Respiratory, Non-Enteric Coronavirus Related to Transmissible Gastroenteritis. Vet. Q 1986, 8, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Saif, L.J.; Jung, K. Comparative Pathogenesis of Bovine and Porcine Respiratory Coronaviruses in the Animal Host Species and SARS-CoV-2 in Humans. J. Clin. Microbiol. 2020, 58, e01355-20. [Google Scholar] [CrossRef]
- Jung, K.; Alekseev, K.P.; Zhang, X.; Cheon, D.-S.; Vlasova, A.N.; Saif, L.J. Altered pathogenesis of porcine respiratory coronavirus in pigs due to immunosuppressive effects of dexamethasone: Implications for corticosteroid use in treatment of severe acute respiratory syndrome coronavirus. J. Virol. 2007, 81, 13681–13693. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Gimenez-Lirola, L.G.; Halbur, P.G. Polymicrobial respiratory disease in pigs. Anim. Health Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef]
- Jonsdottir, H.R.; Dijkman, R. Coronaviruses and the human airway: A universal system for virus-host interaction studies. Virol. J. 2016, 13, 24. [Google Scholar] [CrossRef] [Green Version]
- Fulcher, M.L.; Gabriel, S.; Burns, K.A.; Yankaskas, J.R.; Randell, S.H. Well-Differentiated Human Airway Epithelial Cell Cultures. In Human Cell Culture Protocols; Picot, J., Ed.; Humana Press: Totowa, NJ, USA, 2005; Volume 107, pp. 183–206. [Google Scholar]
- Milewska, A.; Kula-Pacurar, A.; Wadas, J.; Suder, A.; Szczepanski, A.; Dabrowska, A.; Owczarek, K.; Marcello, A.; Ochman, M.; Stacel, T.; et al. Replication of SARS-CoV-2 in human respiratory epithelium. J. Virol. 2020, 94, e00957-20. [Google Scholar] [CrossRef]
- Dijkman, R.; Jebbink, M.F.; Koekkoek, S.M.; Deijs, M.; Jónsdóttir, H.R.; Molenkamp, R.; Ieven, M.; Goossens, H.; Thiel, V.; Van Der Hoek, L. Isolation and characterization of current human coronavirus strains in primary human epithelial cell cultures reveal differences in target cell tropism. J. Virol. 2013, 87, 6081–6090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.-H.; Yang, W.; Beineke, A.; Dijkman, R.; Matrosovich, M.; Baumgärtner, W.; Thiel, V.E.; Valentin-Weigand, P.; Meng, F.; Herrler, G. The differentiated airway epithelium infected by influenza viruses maintains the barrier function despite a dramatic loss of ciliated cells. Sci. Rep. 2016, 6, 39668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeager, C.L.; Ashmun, R.A.; Williams, R.K.; Cardellichio, C.B.; Shapiro, L.H.; Look, A.T.; Holmes, K.V. Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 1992, 357, 420–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laude, H.; Van Reeth, K.; Pensaert, M. Porcine respiratory coronavirus- molecular features and virus-host interactions.pdf. Vet. Res. 1993, 24, 125–150. [Google Scholar]
- Delmas, B.; Gelfi, J.; L’Haridon, R.; Vogel, L.K.; Sjöström, H.; Norén, O.; Laude, H. Aminopeptidase N is a major receptor for the enteropathogenic coronavirus TGEV. Nature 1992, 357, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Van der Velden, V.H.; Wierenga-Wolf, A.F.; Adriaansen-Soeting, P.W.; OBerbeerk, S.G.; Möller, G.M.; Hoogsteden, H.C.; Versnel, M.A. Expression of aminopeptidase N and dipeptidyl peptidase IV in the healthy and asthmatic bronchus. Clin. Exp. Allergy 1998, 1, 110–120. [Google Scholar] [CrossRef]
- Schwegmann-Wessels, C.; Zimmer, G.; Laude, H.; Enjuanes, L.; Herrler, G. Binding of transmissible gastroenteritis coronavirus to cell surface sialoglycoproteins. J. Virol. 2002, 76, 6037–6043. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.-L.; Yang, W.; Peng, J.-Y.; Sawatsky, B.; Von Messling, V.; Herrler, G.; Wu, N.-H. Avian Influenza A Virus Infects Swine Airway Epithelial Cells without Prior Adaptation. Viruses 2020, 12, 589. [Google Scholar] [CrossRef]
- Krimmling, T.; Schwegmann-Weßels, C. Comparison of mono- and co-infection by swine influenza A viruses and porcine respiratory coronavirus in porcine precision-cut lung slices. Res. Vet. Sci. 2017, 115, 470–477. [Google Scholar] [CrossRef]
- Klemens, J.; Ciurkiewicz, M.; Chludzinski, E.; Iseringhausen, M.; Klotz, D.; Pfankuche, V.M.; Ulrich, R.; Herder, V.; Puff, C.; Baumgärtner, W.; et al. Neurotoxic potential of reactive astrocytes in canine distemper demyelinating leukoencephalitis. Sci. Rep. 2019, 9, 11689. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Li, G.; Sui, X.; Yin, J.; Wang, H.; Ren, X. Expression and functional analysis of porcine aminopeptidase N produced in prokaryotic expression system. J. Biotechnol. 2009, 141, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.P.; Look, D.C.; Shi, L.; Hickey, M.; Pewe, L.; Netland, J.; Farzan, M.; Wohlford-Lenane, C.; Perlman, S.; McCray, P.B. ACE2 receptor expression and severe acute respiratory syndrome coronavirus infection depend on differentiation of human airway epithelia. J. Virol. 2005, 79, 14614–14621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.W.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.A.; Zaki, A.; Fouchier, R.A.M.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossen, J.W.A.; Voorhout, W.; Horzinek, M.; Van Der Ende, A.; Strous, G.; Rottier, P. MHV-A59 Enters Polarized Murine Epithelial Cells through the Apical Surface but Is Released Basolaterally. Virology 1995, 210, 54–66. [Google Scholar] [CrossRef] [Green Version]
- Rossen, J.W.; Bekker, C.P.; Voorhout, W.F.; Strous, G.J.; Van Der Ende, A.; Rottier, P.J. Entry and Release of Transmissible Gastroenteritis Coronavirus Are Restricted to Apical Surfaces of Polarized Epithelial Cells. J. Virol. 1994, 68, 7966–7973. [Google Scholar] [CrossRef] [Green Version]
- Rossen, J.W.A.; Kouame, J.; Goedheer, A.J.W.; Vennema, H.; Rottier, P.J.M. Feline and canine coronaviruses are released from the basolateral side of polarized epithelial LLC-PK1 cells expressing the recombinant feline aminopeptidase-N cDNA. Arch. Virol. 2001, 146, 791–799. [Google Scholar] [CrossRef]
- Lukassen, S.; Chua, R.L.; Trefzer, T.; Kahn, N.C.; Schneider, M.A.; Muley, T.; Winter, H.; Meister, M.; Veith, C.; Boots, A.W.; et al. SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef]
- Cong, Y.; Li, X.; Bai, Y.; Lv, X.; Herrler, G.; Enjuanes, L.; Zhou, X.; Qu, B.; Meng, F.; Cong, C.; et al. Porcine aminopeptidase N mediated polarized infection by porcine epidemic diarrhea virus in target cells. Virology 2015, 478, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tata, P.R.; Rajagopal, J. Plasticity in the lung: Making and breaking cell identity. Development 2017, 144, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Cozens, D.; Sutherland, E.; Marchesi, F.; Taylor, G.; Berry, C.C.; Davies, R.L. Temporal differentiation of bovine airway epithelial 2 cells grown at an air-liquid interface. Sci. Rep. 2018, 8, 14893. [Google Scholar] [CrossRef] [Green Version]
- Ravindra, N.G.; Alfajaro, M.M.; Gasque, V.; Wei, J.; Filler, R.B.; Huston, N.C.; Wan, H.; Szigeti-Buck, K.; Wang, B.; Montgomery, R.R.; et al. Single-cell longitudinal analysis of SARS-CoV-2 infection in human bronchial epithelial cells. bioRxiv 2020. [Google Scholar] [CrossRef]
- Delmas, B.; Gelfi, J.; Sjöström, H.; Noren, O.; Laude, H. Futher characterization of aminopepetidase-N as a receptor for coronaviruses. Adv Exp Med Biol. 1993, 342, 293–298. [Google Scholar] [PubMed] [Green Version]
- Wang, J.; Li, Y.; Wang, S.; Liu, F. Dynamics of transmissible gastroenteritis virus internalization unraveled by single-virus tracking in live cells. FASEB J. 2020, 34, 4653–4669. [Google Scholar] [CrossRef] [PubMed]
- Bertram, S.; Dijkman, R.; Habjan, M.; Heurich, A.; Gierer, S.; Glowacka, I.; Welsch, K.; Winkler, M.; Schneider, H.; Hofmann-Winkler, H.; et al. TMPRSS2 activates the human coronavirus 229E for cathepsin-independent host cell entry and is expressed in viral target cells in the respiratory epithelium. J. Virol. 2013, 87, 6150–6160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhang, S.; Shen, Y.; Yang, Q. Epidermal growth factor receptor is a co-factor for transmissible gastroenteritis virus entry. Virology 2018, 521, 33–43. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, J.-Y.; Punyadarsaniya, D.; Shin, D.-L.; Pavasutthipaisit, S.; Beineke, A.; Li, G.; Wu, N.-H.; Herrler, G. The Cell Tropism of Porcine Respiratory Coronavirus for Airway Epithelial Cells Is Determined by the Expression of Porcine Aminopeptidase N. Viruses 2020, 12, 1211. https://doi.org/10.3390/v12111211
Peng J-Y, Punyadarsaniya D, Shin D-L, Pavasutthipaisit S, Beineke A, Li G, Wu N-H, Herrler G. The Cell Tropism of Porcine Respiratory Coronavirus for Airway Epithelial Cells Is Determined by the Expression of Porcine Aminopeptidase N. Viruses. 2020; 12(11):1211. https://doi.org/10.3390/v12111211
Chicago/Turabian StylePeng, Ju-Yi, Darsaniya Punyadarsaniya, Dai-Lun Shin, Suvarin Pavasutthipaisit, Andreas Beineke, Guangxing Li, Nai-Huei Wu, and Georg Herrler. 2020. "The Cell Tropism of Porcine Respiratory Coronavirus for Airway Epithelial Cells Is Determined by the Expression of Porcine Aminopeptidase N" Viruses 12, no. 11: 1211. https://doi.org/10.3390/v12111211
APA StylePeng, J. -Y., Punyadarsaniya, D., Shin, D. -L., Pavasutthipaisit, S., Beineke, A., Li, G., Wu, N. -H., & Herrler, G. (2020). The Cell Tropism of Porcine Respiratory Coronavirus for Airway Epithelial Cells Is Determined by the Expression of Porcine Aminopeptidase N. Viruses, 12(11), 1211. https://doi.org/10.3390/v12111211