Population Difference in Allele Frequency of HLA-C*05 and Its Correlation with COVID-19 Mortality



Abstract
:1. Introduction
2. Materials and Methods
Statistical Analysis
3. Results
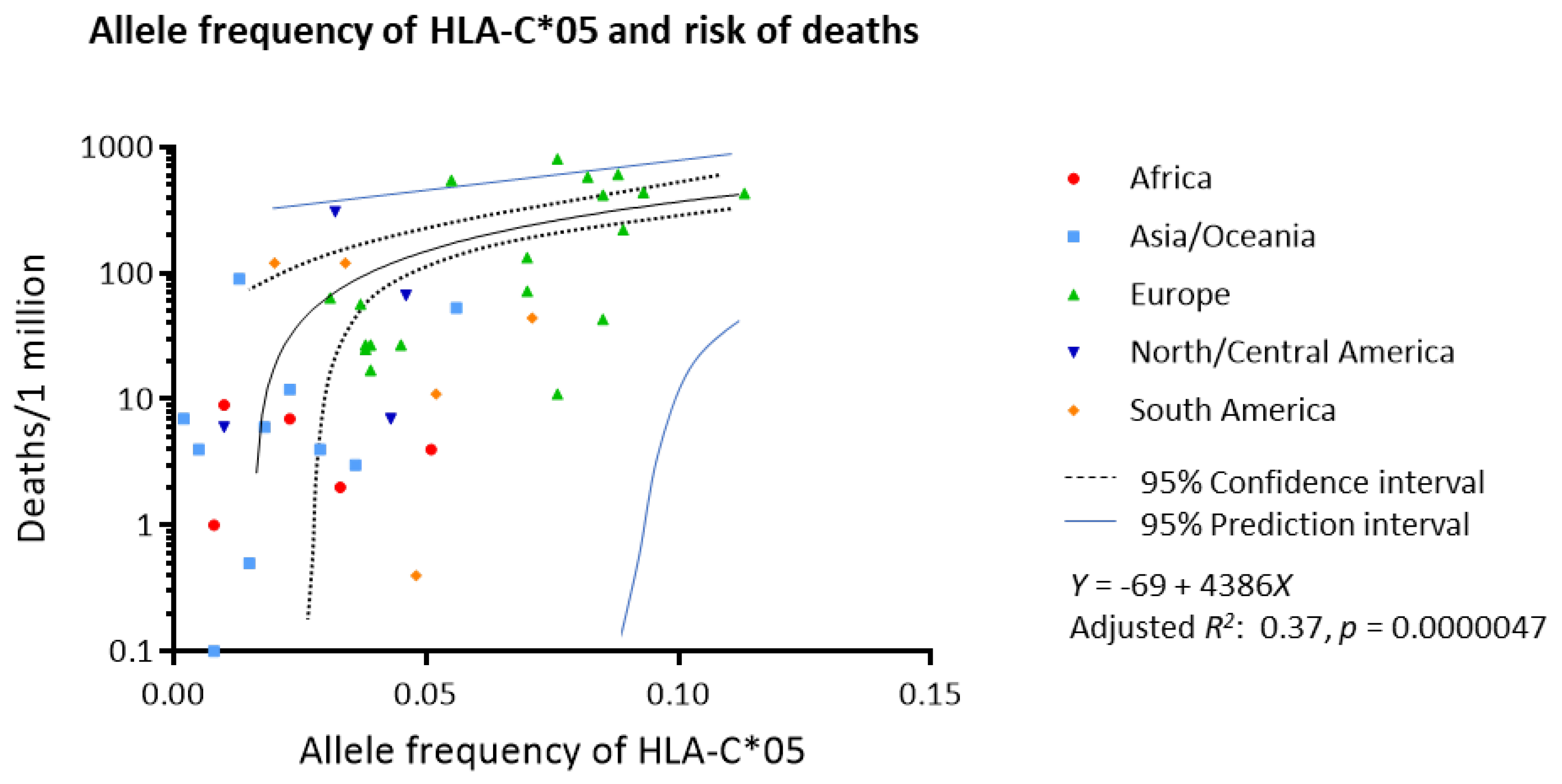
3.1. Association between Worldwide Allele Frequency of HLAs and Mortality to COVID-19
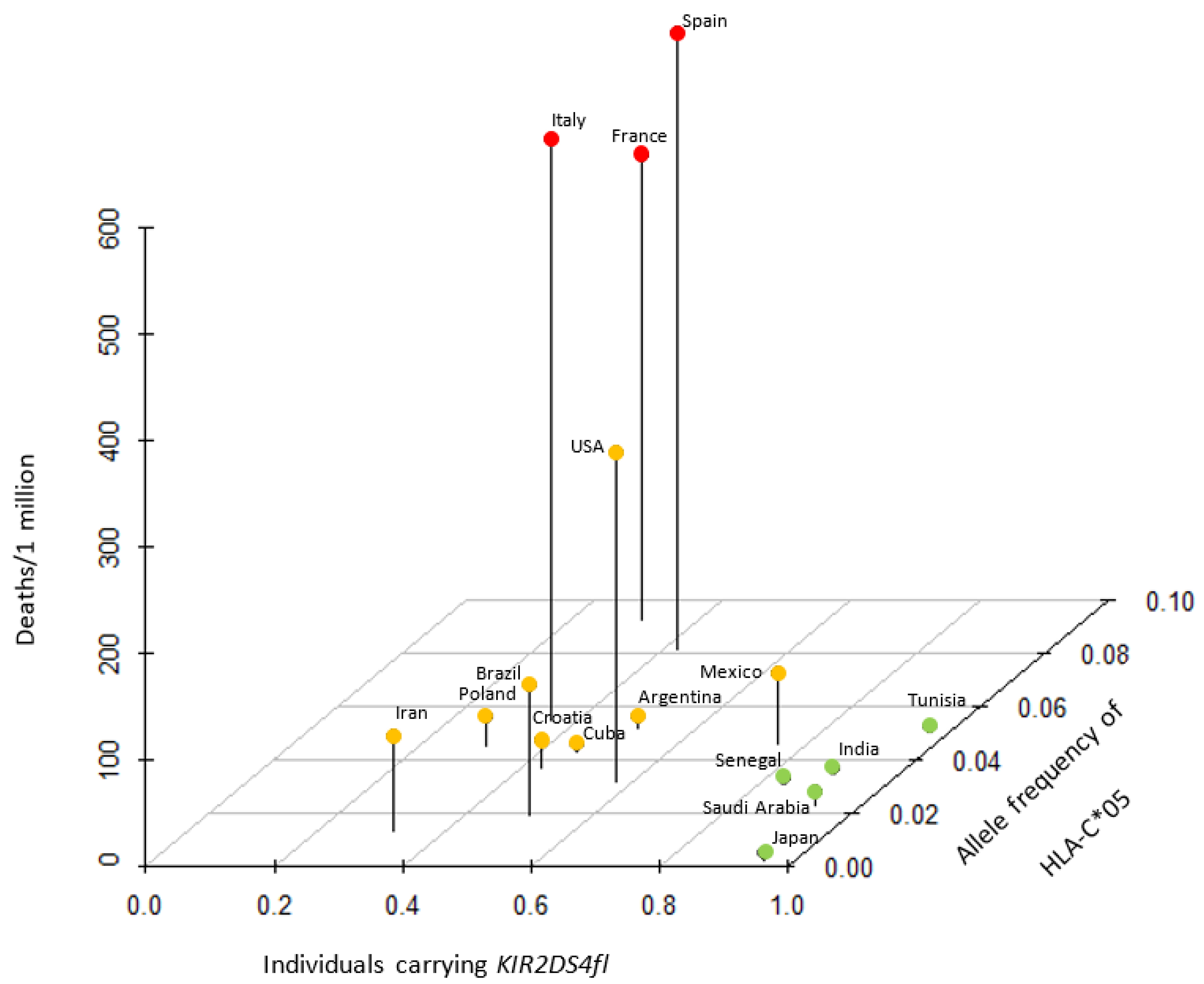
3.2. Association between HLA-C*05 and Its Receptor, KIR2DS4
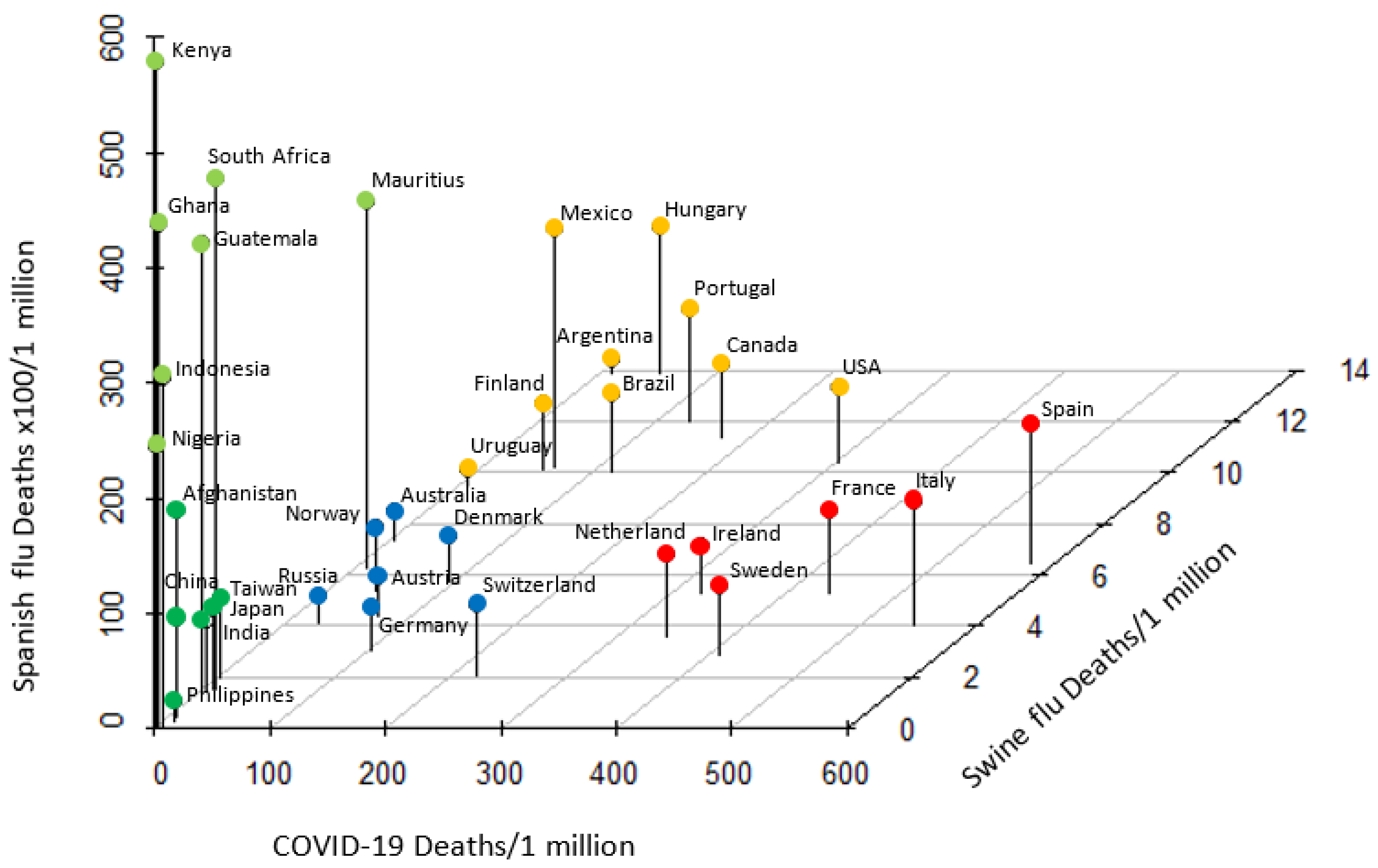
3.3. Distinct Pattern of Response to Historic and COVID-19 Pandemics
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wang, F.; Hou, H.; Luo, Y.; Tang, G.; Wu, S.; Huang, M.; Liu, W.; Zhu, Y.; Lin, Q.; Mao, L.; et al. The laboratory tests and host immunity of COVID-19 patients with different severity of illness. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Verity, R.; Okell, L.C.; Dorigatti, I.; Winskill, P.; Whittaker, C.; Imai, N.; Cuomo-Dannenburg, G.; Thompson, H.; Walker, P.G.T.; Fu, H.; et al. Estimates of the severity of coronavirus disease 2019: A model-based analysis. Lancet Infect. Dis. 2020, 20, 669–677. [Google Scholar] [CrossRef]
- Anyane-Yeboa, A.; Sato, T.; Sakuraba, A. Racial disparities in COVID-19 deaths reveal harsh truths about structural inequality in America. J. Intern. Med. 2020. [Google Scholar] [CrossRef]
- Zhao, M. Cytokine storm and immunomodulatory therapy in COVID-19: Role of chloroquine and anti-IL-6 monoclonal antibodies. Int. J. Antimicrob. Agents 2020, 55, 105982. [Google Scholar] [CrossRef]
- Vabret, N.; Britton, G.J.; Gruber, C.; Hegde, S.; Kim, J.; Kuksin, M.; Levantovsky, R.; Malle, L.; Moreira, A.; Park, M.D.; et al. Immunology of COVID-19: Current State of the Science. Immunity 2020, 52, 910–941. [Google Scholar] [CrossRef]
- Mangalmurti, N.; Hunter, C.A. Cytokine Storms: Understanding COVID-19. Immunity 2020, 53, 19–25. [Google Scholar] [CrossRef]
- Toniati, P.; Piva, S.; Cattalini, M.; Garrafa, E.; Regola, F.; Castelli, F.; Franceschini, F.; Airò, P.; Bazzani, C.; Beindorf, E.-A.; et al. Tocilizumab for the treatment of severe COVID-19 pneumonia with hyperinflammatory syndrome and acute respiratory failure: A single center study of 100 patients in Brescia, Italy. Autoimmun. Rev. 2020, 19, 102568. [Google Scholar] [CrossRef]
- Sim, M.J.W.; Rajagopalan, S.; Altmann, D.M.; Boyton, R.J.; Sun, P.D.; Long, E.O. Human NK cell receptor KIR2DS4 detects a conserved bacterial epitope presented by HLA-C. Proc. Natl. Acad. Sci. USA 2019, 116, 12964–12973. [Google Scholar] [CrossRef] [Green Version]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The cytokine storm in COVID-19: An overview of the involvement of the chemokine/chemokine-receptor system. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef]
- Sun, Y.; Xi, Y. Association between HLA Gene Polymorphism and the Genetic Susceptibility of SARS Infection. In HLA and Associated Important Diseases; InTech: London, UK, 2014. [Google Scholar]
- Sanchez-Mazas, A. A review of HLA allele and SNP associations with highly prevalent infectious diseases in human populations. Swiss Med. Wkly. 2020, 150, w20214. [Google Scholar]
- Prugnolle, F.; Manica, A.; Charpentier, M.; Guégan, J.F.; Guernier, V.; Balloux, F. Pathogen-Driven Selection and Worldwide HLA Class I Diversity. Curr. Biol. 2005, 15, 1022–1027. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Galarza, F.F.; McCabe, A.; Santos, E.; Jones, J.; Takeshita, L.; Ortega-Rivera, N.D.; Cid-Pavon, G.M.D.; Ramsbottom, K.; Ghattaoraya, G.; Alfirevic, A.; et al. Allele frequency net database (AFND) 2020 update: Gold-standard data classification, open access genotype data and new query tools. Nucleic Acids Res. 2020, 48, D783–D788. [Google Scholar] [CrossRef]
- Worldometer COVID-19 Coronavirus Pandemic. Available online: https://www.worldometers.info/coronavirus/ (accessed on 2 October 2020).
- Goodson-Gregg, F.J.; Krepel, S.A.; Anderson, S.K. Tuning of human NK cells by endogenous HLA-C expression. Immunogenetics 2020, 72, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Bontadini, A.; Testi, M.; Cuccia, M.; Martinetti, M.; Carcassi, C.; Chiesa, A.; Cosentini, E.; Dametto, E.; Frison, S.; Iannone, A.; et al. Distribution of killer cell immunoglobulin-like receptors genes in the Italian Caucasian population. J. Transl. Med. 2006, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Tajik, N.; Shahsavar, F.; Mousavi, T.; Radjabzadeh, M.F. Distribution ofKIRgenes in the Iranian population. Tissue Antigens 2009, 74, 22–31. [Google Scholar] [CrossRef]
- Yawata, M.; Yawata, N.; McQueen, K.; Cheng, N.; Guethlein, L.; Rajalingam, R.; Shilling, H.; Parham, P. Predominance of group A KIR haplotypes in Japanese associated with diverse NK cell repertoires of KIR expression. Immunogenetics 2002, 54, 543–550. [Google Scholar] [CrossRef]
- Jiang, K.; Zhu, F.M.; Lv, Q.F.; Yan, L.X. Distribution of killer cell immunoglobulin-like receptor genes in the Chinese Han population. Tissue Antigens 2005, 65, 556–563. [Google Scholar] [CrossRef]
- Whang, D.H.; Park, H.; Yoon, J.A.; Park, M.H. Haplotype analysis of killer cell immunoglobulin-like receptor genes in 77 Korean families. Hum. Immunol. 2005, 66, 146–154. [Google Scholar] [CrossRef]
- Johnson, N.P.; Mueller, J. Updating the accounts: Global mortality of the 1918–1920 “Spanish” influenza pandemic. Bull. Hist. Med. 2002, 76, 105–115. [Google Scholar] [CrossRef]
- 2009 Swine Flu Pandemic by Country. Available online: https://en.wikipedia.org/wiki/2009_swine_flu_pandemic_by_country (accessed on 18 May 2020).
- Sabouri Ghannad, M.; Hajilooi, M.; Solgi, G. HLA-KIR Interactions and Immunity to Viral Infections. Res. Mol. Med. 2014, 2, 1–20. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Graef, T.; Moesta, A.K.; Norman, P.J.; Abi-Rached, L.; Vago, L.; Older Aguilar, A.M.; Gleimer, M.; Hammond, J.A.; Guethlein, L.A.; Bushnell, D.A.; et al. KIR2DS4 is a product of gene conversion with KIR3DL2 that introduced specificity for HLA-A*11 while diminishing avidity for HLA-C. J. Exp. Med. 2009, 206, 2557–2572. [Google Scholar] [CrossRef] [Green Version]
- Barreiro, L.B.; Quintana-Murci, L. From evolutionary genetics to human immunology: How selection shapes host defence genes. Nat. Rev. Genet. 2010, 11, 17–30. [Google Scholar] [CrossRef]
- Peiris, J.S.M.; Yuen, K.Y.; Osterhaus, A.D.M.E.; Stöhr, K. The Severe Acute Respiratory Syndrome. N. Engl. J. Med. 2003, 349, 2431–2441. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.F.; Velickovic, Z.; Ashton, L.J.; Dyer, W.B.; Geczy, A.F.; Dunckley, H.; Lynch, G.W.; Sullivan, J.S. Influence of HLA gene polymorphisms on susceptibility and outcome post infection with the SARS-CoV virus. Virol. Sin. 2014, 29, 128–130. [Google Scholar] [CrossRef] [Green Version]
- Keicho, N.; Itoyama, S.; Kashiwase, K.; Phi, N.C.; Long, H.T.; Ha, L.D.; Ban, V.V.; Hoa, B.K.; Hang, N.T.L.; Hijikata, M.; et al. Association of human leukocyte antigen class II alleles with severe acute respiratory syndrome in the Vietnamese population. Hum. Immunol. 2009, 70, 527–531. [Google Scholar] [CrossRef]
- Tussey, L.G.; Rowland-Jones, S.; Zheng, T.S.; Androlewicz, M.J.; Cresswell, P.; Frelinger, J.A.; McMichael, A.J. Different MHC class I alleles compete for presentation of overlapping viral epitopes. Immunity 1995, 3, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhang, W.; Zhang, J.; He, J.; Zhu, F. Distribution of HLA allele frequencies in 82 Chinese individuals with coronavirus disease-2019 (COVID-19). HLA 2020. [Google Scholar] [CrossRef]
- Parham, P. Immunogenetics of killer-cell immunoglobulin-like receptors. Tissue Antigens 2003, 62, 194–200. [Google Scholar] [CrossRef]
- Yawata, M.; Yawata, N.; Draghi, M.; Little, A.M.; Partheniou, F.; Parham, P. Roles for HLA and KIR polymorphisms in natural killer cell repertoire selection and modulation of effector function. J. Exp. Med. 2006, 203, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Rajaram, S.; Canaday, L.M.; Ochayon, D.E.; Rangel, K.M.; Ali, A.; Gyurova, I.E.; Krishnamurthy, D.; Fletcher, J.S.; Reighard, S.D.; Cox, A.; et al. The Promise and Peril of Natural Killer Cell Therapies in Pulmonary Infection. Immunity 2020, 52, 887–889. [Google Scholar] [CrossRef]
- Whittaker, E.; Bamford, A.; Kenny, J.; Kaforou, M.; Jones, C.E.; Shah, P.; Ramnarayan, P.; Fraisse, A.; Miller, O.; Davies, P.; et al. Clinical Characteristics of 58 Children with a Pediatric Inflammatory Multisystem Syndrome Temporally Associated With SARS-CoV-2. JAMA 2020, 324, 259–269. [Google Scholar] [CrossRef]
- La, D.; Czarnecki, C.; El-Gabalawy, H.; Kumar, A.; Meyers, A.F.A.; Bastien, N.; Simonsen, J.N.; Plummer, F.A.; Luo, M. Enrichment of Variations in KIR3DL1/S1 and KIR2DL2/L3 among H1N1/09 ICU Patients: An Exploratory Study. PLoS ONE 2011, 6, e29200. [Google Scholar] [CrossRef]
- Kobasa, D.; Jones, S.M.; Shinya, K.; Kash, J.C.; Copps, J.; Ebihara, H.; Hatta, Y.; Hyun Kim, J.; Halfmann, P.; Hatta, M.; et al. Aberrant innate immune response in lethal infection of macaques with the 1918 influenza virus. Nature 2007, 445, 319–323. [Google Scholar] [CrossRef]
- Zwerling, A.; Behr, M.A.; Verma, A.; Brewer, T.F.; Menzies, D.; Pai, M. The BCG World Atlas: A Database of Global BCG Vaccination Policies and Practices. PLoS Med. 2011, 8, e1001012. [Google Scholar] [CrossRef] [Green Version]
- Hiby, S.E.; Walker, J.J.; O’Shaughnessy, K.M.; Redman, C.W.G.; Carrington, M.; Trowsdale, J.; Moffett, A. Combinations of Maternal KIR and Fetal HLA-C Genes Influence the Risk of Preeclampsia and Reproductive Success. J. Exp. Med. 2004, 200, 957–965. [Google Scholar] [CrossRef]
- Ayoub, F.; Sato, T.; Sakuraba, A. Football and COVID-19 risk: Correlation is not causation. Clin. Microbiol. Infect. 2020. [Google Scholar] [CrossRef]
- Ellinghaus, D.; Degenhardt, F.; Bujanda, L.; Buti, M.; Albillos, A.; Invernizzi, P.; Fernández, J.; Prati, D.; Baselli, G.; Asselta, R.; et al. Genomewide Association Study of Severe Covid-19 with Respiratory Failure. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Wang, F.; Huang, S.; Gao, H.; Zhou, Y.; Lai, C.; Li, Z.; Xian, W.; Qian, X.; Li, Z.; Huang, Y.; et al. Initial Whole Genome Sequencing and Analysis of the Host Genetic Contribution to COVID-19 Severity and Susceptibility. medRxiv 2020. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| HLA | Adjusted R2 | p Value | Adjusted p Value | HLA | Adjusted R2 | p Value | Adjusted p Value | HLA | Adjusted R2 | p Value | Adjusted p Value |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A*01 | 0.16 | 0.00059 | 0.040 * | B*07 | 0.24 | 0.000012 | 0.00081 * | C*01 | −0.0016 | 0.34 | 1 |
| A*02 | 0.017 | 0.15 | 1 | B*08 | 0.16 | 0.00070 | 0.047 * | C*02 | −0.013 | 0.51 | 1 |
| A*03 | 0.10 | 0.0046 | 0.31 | B*13 | 0.042 | 0.057 | 1 | C*03 | −0.024 | 0.86 | 1 |
| A*11 | 0.020 | 0.13 | 1 | B*14 | 0.15 | 0.0015 | 0.099 | C*04 | −0.012 | 0.49 | 1 |
| A*23 | 0.032 | 0.090 | 1 | B*15 | 0.026 | 0.10 | 1 | C*05 | 0.37 | 0.0000047 | 0.00032 * |
| A*24 | 0.032 | 0.078 | 1 | B*18 | −0.011 | 0.59 | 1 | C*06 | −0.019 | 0.63 | 1 |
| A*25 | 0.022 | 0.15 | 1 | B*27 | −0.0061 | 0.44 | 1 | C*07 | 0.18 | 0.0023 | 0.15 |
| A*26 | −0.013 | 0.69 | 1 | B*35 | −0.0094 | 0.55 | 1 | C*08 | −0.00044 | 0.33 | 1 |
| A*29 | 0.066 | 0.021 | 1 | B*37 | −0.014 | 0.67 | 1 | C*12 | −0.022 | 0.84 | 1 |
| A*30 | 0.0081 | 0.22 | 1 | B*38 | −0.0057 | 0.42 | 1 | C*15 | 0.064 | 0.051 | 1 |
| A*31 | 0.012 | 0.58 | 1 | B*39 | −0.0079 | 0.48 | 1 | C*16 | −0.029 | 0.90 | 1 |
| A*32 | 0.014 | 0.18 | 1 | B*40 | 0.030 | 0.096 | 1 | C*17 | 0.021 | 0.19 | 1 |
| A*33 | 0.11 | 0.0038 | 0.26 | B*41 | −0.0027 | 0.36 | 1 | C*18 | 0.11 | 0.09 | 1 |
| A*34 | 0.030 | 0.12 | 1 | B*42 | 0.057 | 0.054 | 1 | ||||
| A*36 | 0.011 | 0.23 | 1 | B*44 | 0.22 | 0.000032 | 0.0022 * | ||||
| A*43 | −0.022 | 0.53 | 1 | B*45 | 0.027 | 0.12 | 1 | ||||
| A*66 | 0.041 | 0.078 | 1 | B*46 | 0.042 | 0.12 | 1 | ||||
| A*68 | −0.0094 | 0.53 | 1 | B*47 | 0.012 | 0.22 | 1 | ||||
| A*69 | −0.0073 | 0.42 | 1 | B*48 | 0.0049 | 0.27 | 1 | ||||
| A*74 | 0.051 | 0.048 | 1 | B*49 | −0.018 | 0.86 | 1 | ||||
| A*80 | 0.020 | 0.20 | 1 | B*50 | −0.0062 | 0.43 | 1 | ||||
| B*51 | 0.012 | 0.18 | 1 | ||||||||
| B*52 | 0.072 | 0.020 | 1 | ||||||||
| B*53 | 0.030 | 0.10 | 1 | ||||||||
| B*54 | 0.010 | 0.26 | 1 | ||||||||
| B*55 | −0.016 | 0.82 | 1 | ||||||||
| B*56 | 0.020 | 0.16 | 1 | ||||||||
| B*57 | 0.033 | 0.076 | 1 | ||||||||
| B*58 | 0.091 | 0.0089 | 0.60 | ||||||||
| B*59 | 0.060 | 0.19 | 1 | ||||||||
| B*67 | 0.053 | 0.14 | 1 | ||||||||
| B*73 | −0.029 | 0.83 | 1 | ||||||||
| B*81 | 0.021 | 0.24 | 1 | ||||||||
| B*82 | 0.030 | 0.28 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakuraba, A.; Haider, H.; Sato, T. Population Difference in Allele Frequency of HLA-C*05 and Its Correlation with COVID-19 Mortality. Viruses 2020, 12, 1333. https://doi.org/10.3390/v12111333
Sakuraba A, Haider H, Sato T. Population Difference in Allele Frequency of HLA-C*05 and Its Correlation with COVID-19 Mortality. Viruses. 2020; 12(11):1333. https://doi.org/10.3390/v12111333
Chicago/Turabian StyleSakuraba, Atsushi, Haider Haider, and Toshiro Sato. 2020. "Population Difference in Allele Frequency of HLA-C*05 and Its Correlation with COVID-19 Mortality" Viruses 12, no. 11: 1333. https://doi.org/10.3390/v12111333
APA StyleSakuraba, A., Haider, H., & Sato, T. (2020). Population Difference in Allele Frequency of HLA-C*05 and Its Correlation with COVID-19 Mortality. Viruses, 12(11), 1333. https://doi.org/10.3390/v12111333