LysSAP26, a New Recombinant Phage Endolysin with a Broad Spectrum Antibacterial Activity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
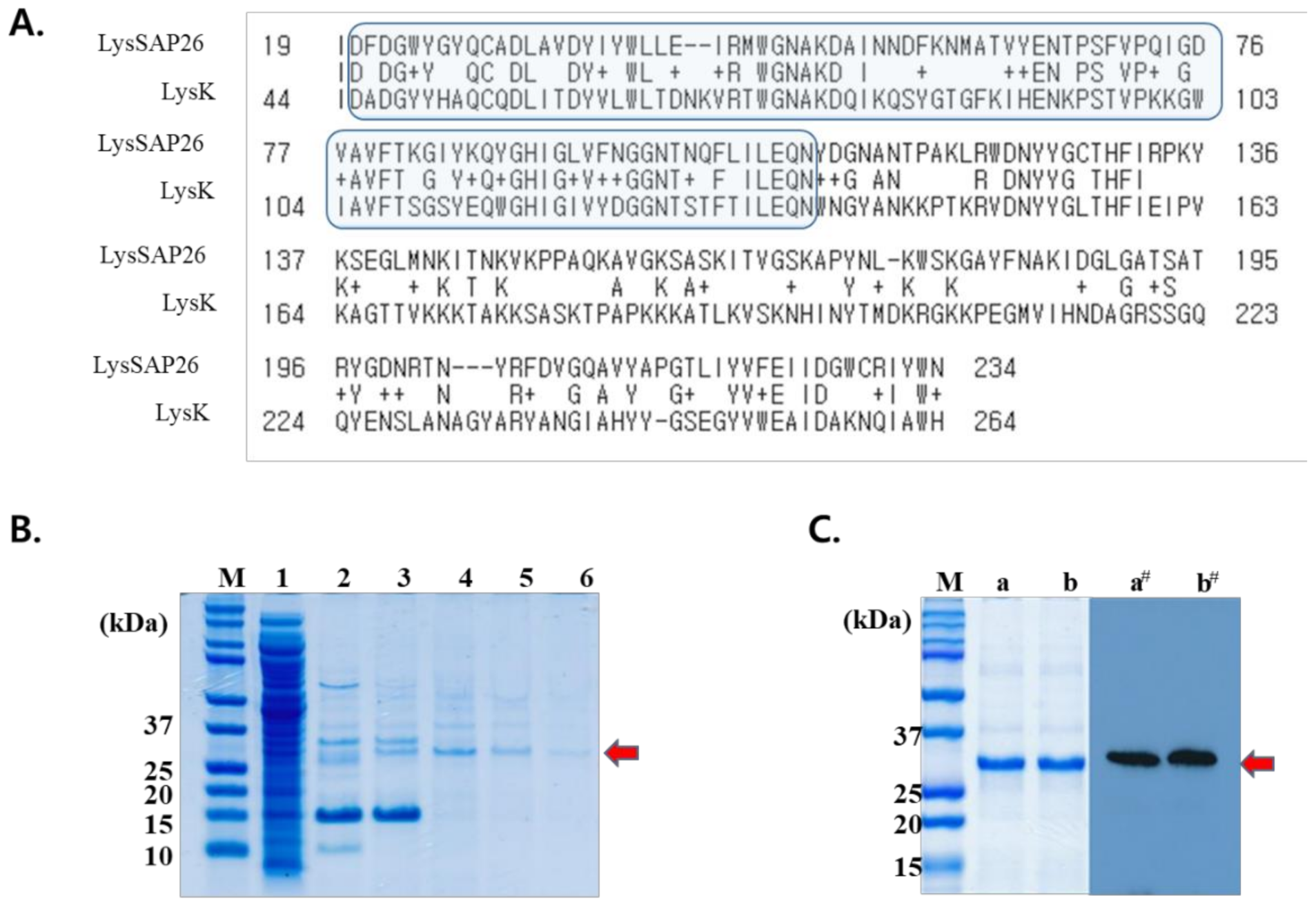
2.2. LysSAP26 Sequence In Silico Analysis
2.3. Construction of the LysSAP26 Expression Vector
2.4. Purification of LysSAP26
2.5. Antibacterial Activity Test of LysSAP26 against Various Clinical Isolates
2.6. Transmission Electron Microscopic (TEM) Analysis of A. baumannii Treated with LysSAP26
2.7. LysSAP26 Protection Assay with A. baumannii Systemic Infection Mouse Model
- Group 1. A negative control group injected with 100 μL of the lysis buffer alone;
- Group 2. First safety test group injected with 35 μg of LysSAP26 alone;
- Group 3. Second safety test group injected with 70 μg of LysSAP26 alone;
- Group 4. An infection control group infected with A. baumannii ATCC 17978;
- Group 5. First lysin-treated group infected with A. baumannii ATCC 17978 and treated with 35 μg of LysSAP26 30 min after infection;
- Group 6. Second lysin-treated group infected with A. baumannii ATCC 17978 and treated with 70 μg of LysSAP26 30 min after infection.
3. Results
3.1. LysSAP26 DNA and Protein Sequence Analysis
3.2. LysSAP26 Purification
3.3. Antibacterial Activity of LysSAP26
3.4. A. baumannii Cell Features Affected by LysSAP26
3.5. In Vivo Protection Assays of LysSAP26
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Medina, E.; Pieper, D.H. Tackling Threats and Future Problems of Multidrug-Resistant Bacteria. Curr. Top. Microbiol. Immunol. 2016, 398, 3–33. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- De la Fuente-Nunez, C.; Torres, M.D.; Mojica, F.J.M.; Lu, T.K. Next-generation precision antimicrobials: Towards personalized treatment of infectious diseases. Curr. Opin. Microbiol. 2017, 37, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, D.W.; Jin, J.S.; Kim, J. Antimicrobial activity of LysSS, a novel phage endolysin, against Acinetobacter baumannii and Pseudomonas aeruginosa. J. Glob. Antimicrob. Resist. 2020, 22, 32–39. [Google Scholar] [CrossRef]
- Dams, D.; Briers, Y. Enzybiotics: Enzyme-Based Antibacterials as Therapeutics. Adv. Exp. Med. Biol. 2019, 1148, 233–253. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Y.; Sun, E.; Hua, L.; Peng, Z.; Wu, B. Characterization of a Lytic Bacteriophage vB_EfaS_PHB08 Harboring Endolysin Lys08 Against Enterococcus faecalis Biofilms. Microorganisms 2020, 8, 1332. [Google Scholar] [CrossRef] [PubMed]
- Grishin, A.V.; Karyagina, A.S.; Vasina, D.V.; Vasina, I.V.; Gushchin, V.A.; Lunin, V.G. Resistance to peptidoglycan-degrading enzymes. Crit. Rev. Microbiol. 2020, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Vermassen, A.; Leroy, S.; Talon, R.; Provot, C.; Popowska, M.; Desvaux, M. Cell Wall Hydrolases in Bacteria: Insight on the Diversity of Cell Wall Amidases, Glycosidases and Peptidases Toward Peptidoglycan. Front. Microbiol. 2019, 10, 331. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.C.B.; Chen, X.; Ho, M.K.Y.; Xia, J.; Leung, S.S.Y. Bacteriophage-derived endolysins to target gram-negative bacteria. Int. J. Pharm. 2020, 589, 119833. [Google Scholar] [CrossRef]
- Rahman, M.; Kim, S.; Kim, S.M.; Seol, S.Y.; Kim, J. Characterization of induced Staphylococcus aureus bacteriophage SAP-26 and its anti-biofilm activity with rifampicin. Biofouling 2011, 27, 1087–1093. [Google Scholar] [CrossRef]
- Bartual, S.G.; Seifert, H.; Hippler, C.; Luzon, M.A.D.; Wisplinghoff, H.; Rodríguez-Valera, F. Development of a Multilocus Sequence Typing Scheme for Characterization of Clinical Isolates of Acinetobacter baumannii. J. Clin. Microbiol. 2005, 43, 4382–4390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, H.; Kim, S.; Kim, M.H.; Kim, S.Y.; Nam, D.; Park, S.C.; Park, S.-H.; Bae, H.; Lee, H.-J.; Cho, J.H.; et al. Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii isolates from a Korean hospital that carry blaOXA-23. Infect. Genet. Evol. 2018, 58, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jin, J.S.; Lee, D.W.; Kim, J. Antibacterial activities of and biofilm removal by Ablysin, an endogenous lysozyme-like protein originated from Acinetobacter baumannii 1656-2. J. Glob. Antimicrob. Resist. 2020, S2213–S7165. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.C.; Dong, S.; Baker, J.R.; Foster-Frey, J.; Pritchard, D.G.; Donovan, D.M. LysK CHAP endopeptidase domain is required for lysis of live staphylococcal cells. FEMS Microbiol. Lett. 2009, 294, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Kim, M.; Ryu, S. Characterization of a novel endolysin LysSA11 and its utility as a potent biocontrol agent against Staphylococcus aureus on food and utensils. Food Microbiol. 2017, 68, 112–120. [Google Scholar] [CrossRef]
- Fischetti, V.A. Bacteriophage lysins as effective antibacterials. Curr. Opin. Microbiol. 2008, 11, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Ghose, C.; Euler, C.W. Gram-Negative Bacterial Lysins. Antibiotics 2020, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Hou, C. Systematic analysis of an amidase domain CHAP in 12 Staphylococcus aureus genomes and 44 staphylococcal phage genomes. Comput. Biol. Chem. 2010, 34, 251–257. [Google Scholar] [CrossRef]
- Horgan, M.; O’Flynn, G.; Garry, J.; Cooney, J.; Coffey, A.; Fitzgerald, G.F.; Ross, R.P.; McAuliffe, O. Phage Lysin LysK Can Be Truncated to Its CHAP Domain and Retain Lytic Activity against Live Antibiotic-Resistant Staphylococci. Appl. Environ. Microbiol. 2009, 75, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Fenton, M.; Casey, P.G.; Hill, C.; Gahan, C.G.; McAuliffe, O.; Mahony, J.O.; Maher, F.; Coffey, A. The truncated phage lysin CHAP(k) eliminates Staphylococcus aureusin the nares of mice. Bioeng. Bugs 2010, 1, 404–407. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, S.; Coffey, A.; Meaney, W.; Fitzgerald, G.F.; Ross, R.P. The Recombinant Phage Lysin LysK Has a Broad Spectrum of Lytic Activity against Clinically Relevant Staphylococci, Including Methicillin-Resistant Staphylococcus aureus. J. Bacteriol. 2005, 187, 7161–7164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zowawi, H.M.; Sartor, A.L.; Sidjabat, H.E.; Balkhy, H.H.; Walsh, T.R.; Al Johani, S.M.; Aljindan, R.Y.; Alfaresi, M.; Ibrahim, E.; Al-Jardani, A.; et al. Molecular Epidemiology of Carbapenem-Resistant Acinetobacter baumannii Isolates in the Gulf Cooperation Council States: Dominance of OXA-23-Type Producers. J. Clin. Microbiol. 2015, 53, 896–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelkader, K.; Gutiérrez, D.; Grimon, D.; Ruas-Madiedo, P.; Lood, C.; Lavigne, R.; Safaan, A.; Khairalla, A.S.; Gaber, Y.; Dishisha, T.; et al. Lysin LysMK34 of Acinetobacter baumannii Bacteriophage PMK34 Has a Turgor Pressure-Dependent Intrinsic Antibacterial Activity and Reverts Colistin Resistance. Appl. Environ. Microbiol. 2020, 86, e01311-20. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Bacterium | Strain | MIC (µg/mL) | MBC (µg/mL) |
|---|---|---|---|
| Staphylococcus aureus | ATCC 33591 | 40 | >80 |
| Klebsiella pneumoniae | KCTC 2208 | 20 | >80 |
| Acinetobacter baumannii | ATCC 17978 | 10 | 10 |
| Acinetobacter baumannii | ATCC 19606 | 10 | 10 |
| Pseudomonas aeruginosa | PA01 | 20 | 20 |
| Escherichia coli | ATCC 25922 | 20 | >80 |
| Bacteria | No. of Isolates Tested | Source 1 | MIC 2 (µg/mL) | MBC 3 (µg/mL) |
|---|---|---|---|---|
| Enterococcus faecium | 20 | KNUHCCP | 40–80 | >80 |
| Staphylococcus aureus | 20 | KNUHCCP | 20 | >80 |
| Klebsiella pneumoniae | 20 | KNUHCCP | 40 | >80 |
| Acinetobacter baumannii | 17 | KNUHCCP | 10 | 10 |
| Pseudomonas aeruginosa | 20 | KNUHCCP | 20–40 | 20–40 |
| Escherichia coli | 20 | KNUHCCP | 20–40 | >80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Jin, J.-S.; Choi, Y.-J.; Kim, J. LysSAP26, a New Recombinant Phage Endolysin with a Broad Spectrum Antibacterial Activity. Viruses 2020, 12, 1340. https://doi.org/10.3390/v12111340
Kim S, Jin J-S, Choi Y-J, Kim J. LysSAP26, a New Recombinant Phage Endolysin with a Broad Spectrum Antibacterial Activity. Viruses. 2020; 12(11):1340. https://doi.org/10.3390/v12111340
Chicago/Turabian StyleKim, Shukho, Jong-Sook Jin, Yoon-Jung Choi, and Jungmin Kim. 2020. "LysSAP26, a New Recombinant Phage Endolysin with a Broad Spectrum Antibacterial Activity" Viruses 12, no. 11: 1340. https://doi.org/10.3390/v12111340
APA StyleKim, S., Jin, J.-S., Choi, Y.-J., & Kim, J. (2020). LysSAP26, a New Recombinant Phage Endolysin with a Broad Spectrum Antibacterial Activity. Viruses, 12(11), 1340. https://doi.org/10.3390/v12111340