User-Friendly Reverse Genetics System for Modification of the Right End of Fowl Adenovirus 4 Genome

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Plasmids and Oligonucleotides
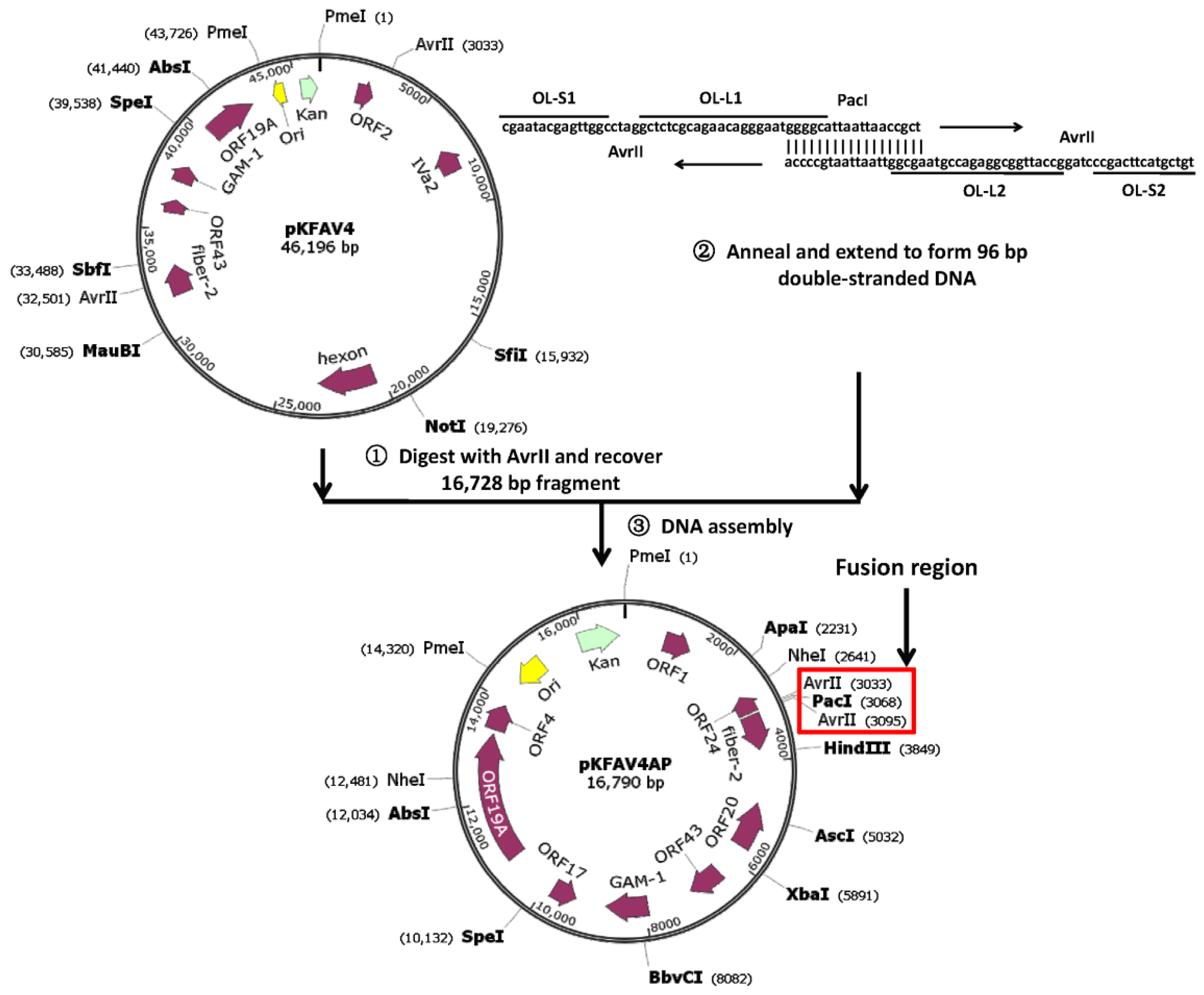
2.2. Construct Intermediate Plasmid pKFAV4AP
2.3. Delete ORF1-ORF2 in pKFAV4
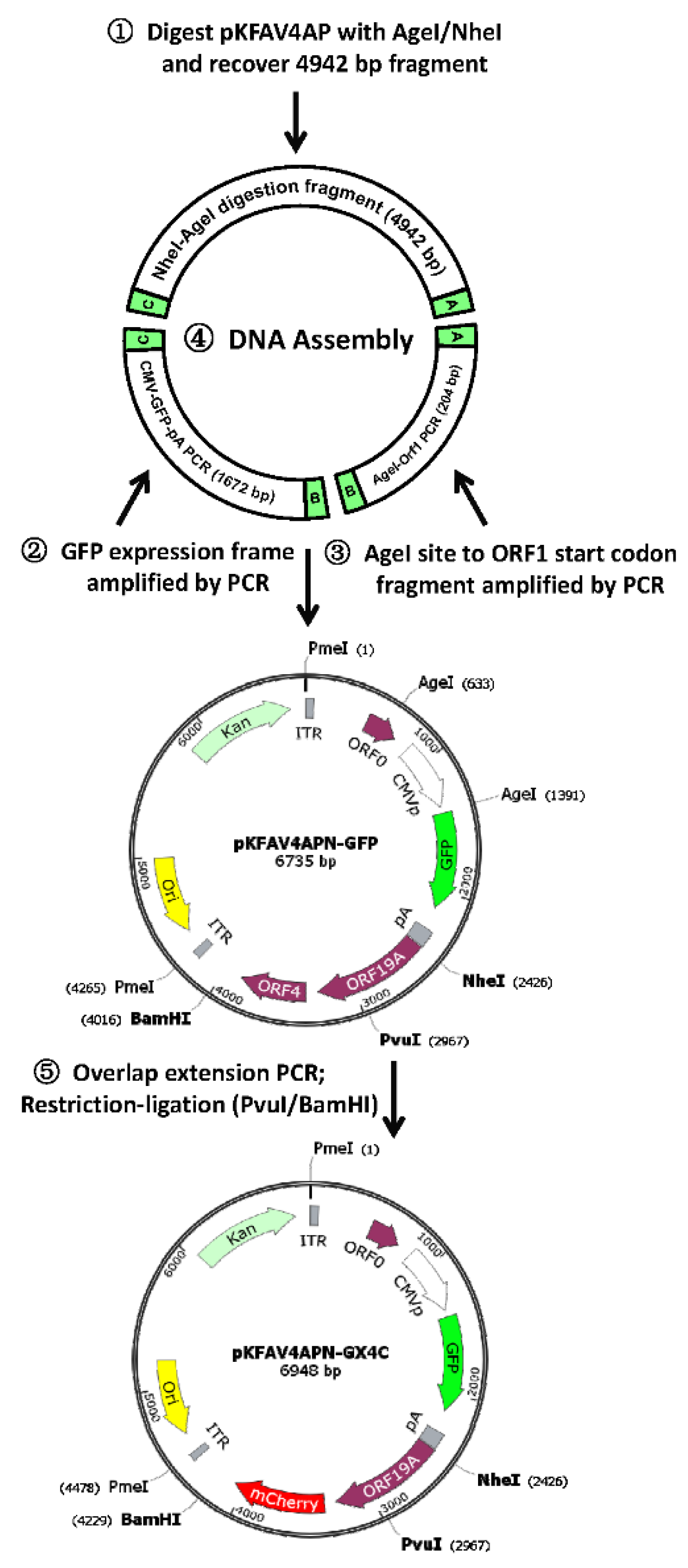
2.4. Replace Coding Sequence of FAdV-4 ORF4 with That of mCherry in pKFAV4-GFP
2.5. Remove the AvrII Site in Fiber-2 Gene in pKFAV4 and Reduce the Size of the Intermediate Plasmid
2.6. Delete ORF19A in pKFAV4-GFP
2.7. Rescue, Amplification, Purification and Titration of Recombinant Virus
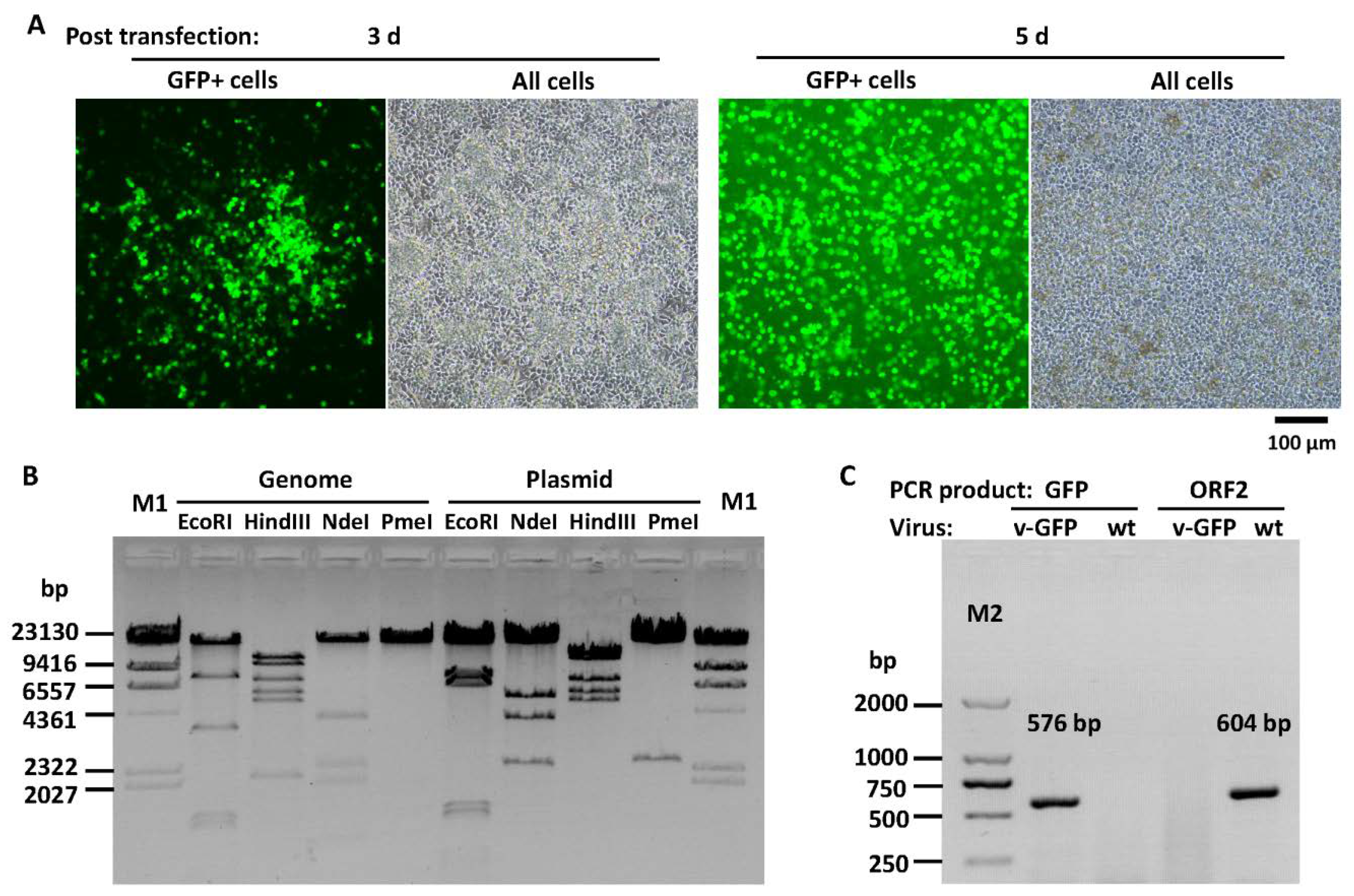
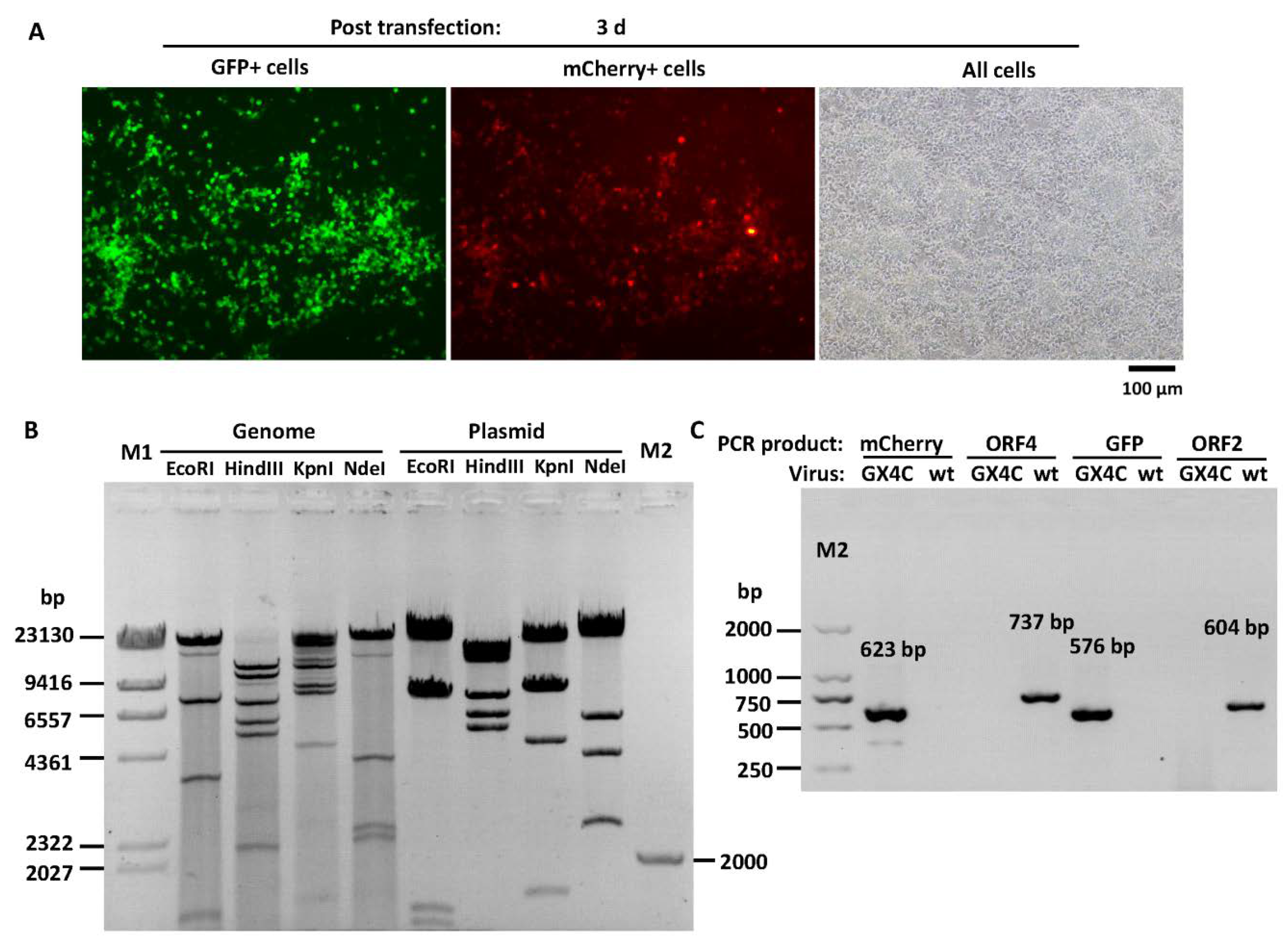
2.8. Identification of Recombinant Virus
2.9. Plaque Forming Experiment
2.10. Viral Inoculation of Embryonated Chicken Eggs
3. Results
3.1. Reverse Genetics System for Modification of the Right End of FAdV-4 Genome
3.2. Construction of Recombinant FAdV-4 with Deletions of ORF1, ORF1b and ORF2
3.3. Construction of Recombinant FAdV-4 with Deletions of ORF1, ORF1b, ORF2 and ORF4
3.4. Modification of the Reverse Genetics Systems
3.5. Construction of Recombinant FAdV-4 with Deletion of ORF1, ORF1b, ORF2 and ORF19A
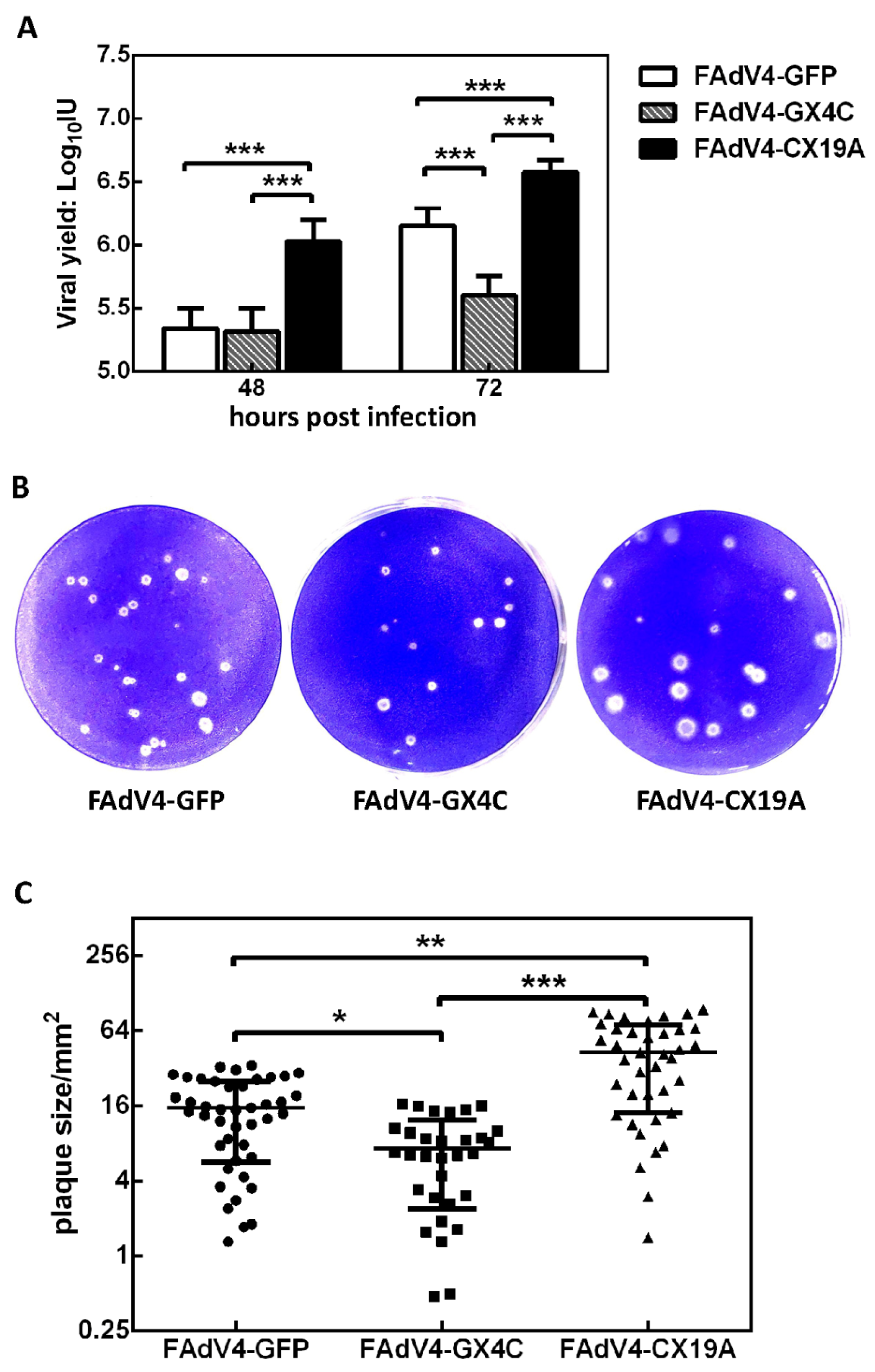
3.6. Growth Property of Recombinant Viruses in LMH Cells
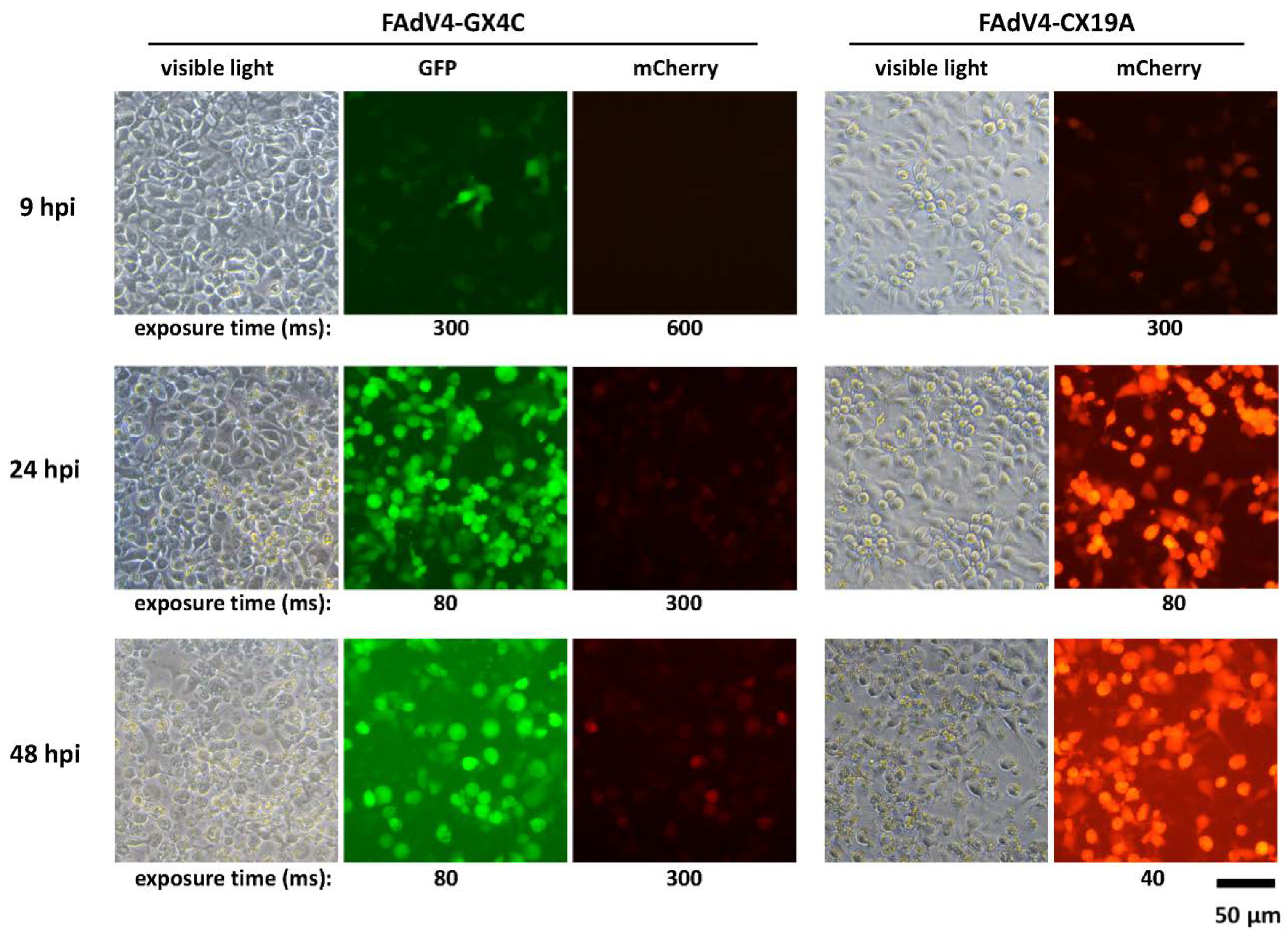
3.7. Expression of mCherry Controlled by the FAdV-4 ORF4 Promoter in LMH Cells
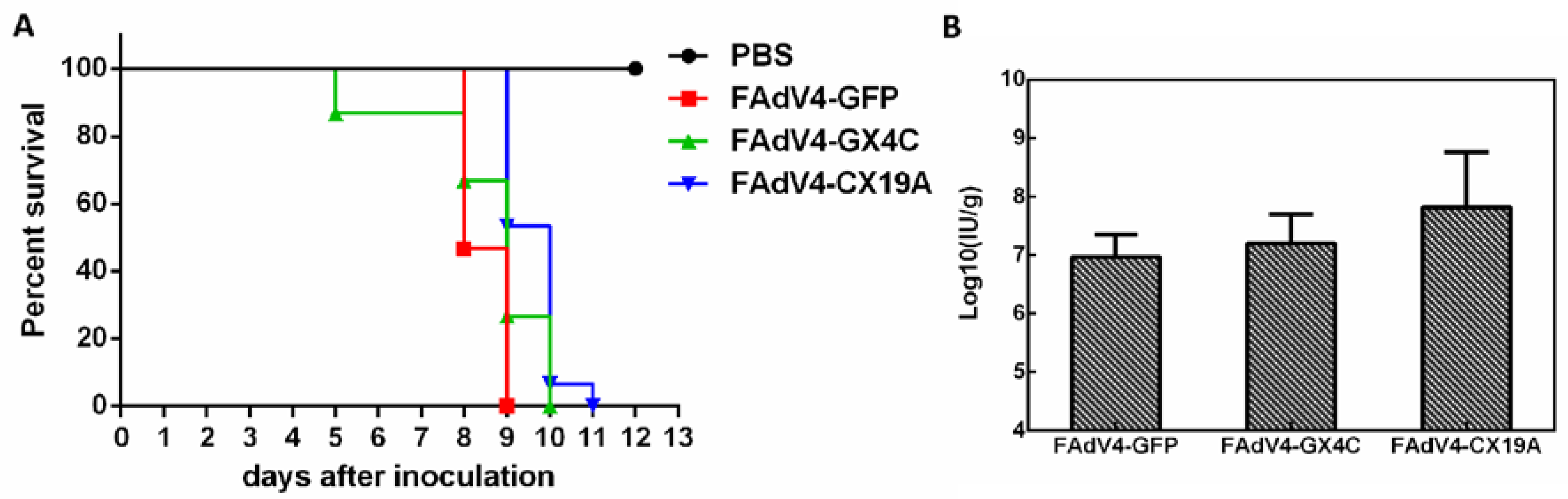
3.8. Embryonic Lethality of Recombinant FAdV-4 Infection in Chicken
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Virus taxonomy: Classification and nomenclature of viruses: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: San Diego, CA, USA, 2011; pp. 129–141. [Google Scholar]
- Corredor, J.C.; Garceac, A.; Krell, P.J.; Nagy, E. Sequence comparison of the right end of fowl adenovirus genomes. Virus Genes 2008, 36, 331–344. [Google Scholar] [CrossRef]
- Schachner, A.; Matos, M.; Grafl, B.; Hess, M. Fowl adenovirus-induced diseases and strategies for their control-a review on the current global situation. Avian Pathol. 2018, 47, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Anjum, A.D.; Sabri, M.A.; Iqbal, Z. Hydropericarditis syndrome in broiler chickens in Pakistan. Vet. Rec 1989, 124, 247–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yin, L.; Zhou, Q.; Peng, P.; Du, Y.; Liu, L.; Zhang, Y.; Xue, C.; Cao, Y. Epidemiological investigation of fowl adenovirus infections in poultry in China during 2015–2018. BMC Vet. Res. 2019, 15, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.H.; Zheng, P.P.; Zhang, T.F.; Wen, G.Y.; Shao, H.B.; Luo, Q.P. Fowl adenovirus serotype 4: Epidemiology, pathogenesis, diagnostic detection, and vaccine strategies. Poult. Sci. 2017, 96, 2630–2640. [Google Scholar] [CrossRef]
- Liu, Y.; Wan, W.; Gao, D.; Li, Y.; Yang, X.; Liu, H.; Yao, H.; Chen, L.; Wang, C.; Zhao, J. Genetic characterization of novel fowl aviadenovirus 4 isolates from outbreaks of hepatitis-hydropericardium syndrome in broiler chickens in China. Emerg. Microbes. Infect. 2016, 5, e117. [Google Scholar] [CrossRef]
- Davison, A.J.; Benko, M.; Harrach, B. Genetic content and evolution of adenoviruses. J. Gen. Virol. 2003, 84, 2895–2908. [Google Scholar] [CrossRef]
- Sheppard, M.; Werner, W.; Tsatas, E.; McCoy, R.; Prowse, S.; Johnson, M. Fowl adenovirus recombinant expressing VP2 of infectious bursal disease virus induces protective immunity against bursal disease. Arch. Virol. 1998, 143, 915–930. [Google Scholar] [CrossRef]
- Corredor, J.C.; Pei, Y.; Nagy, E. Fowl Adenovirus-Based Vaccine Platform. Methods Mol. Biol. 2017, 1581, 29–54. [Google Scholar] [CrossRef]
- Liu, H.; Lu, Z.; Zhang, X.; Guo, X.; Mei, L.; Zou, X.; Zhong, Y.; Wang, M.; Hung, T. Single Plasmid-Based, Upgradable, and Backward-Compatible Adenoviral Vector Systems. Hum. Gene. Ther. 2019. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Mei, L.; Yan, B.; Zou, X.; Hung, T.; Lu, Z. Site-directed modification of adenoviral vector with combined DNA assembly and restriction-ligation cloning. J. Biotechnol. 2019, 307, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.H.; Bi, Z.X.; Guo, X.J.; Zhang, Z.; Zhao, Y.; Wang, M.; Zhu, Y.L.; Jie, H.Y.; Yu, Y.; Hung, T.; et al. DNA assembly technique simplifies the construction of infectious clone of fowl adenovirus. J. Virol. Methods 2018, 257, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.Z.; Zou, X.H.; Dong, L.X.; Qu, J.G.; Song, J.D.; Wang, M.; Guo, L.; Hung, T. Novel recombinant adenovirus type 41 vector and its biological properties. J. Gene Med. 2009, 11, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.L.; Dong, L.X.; Li, M.; Guo, X.J.; Wang, M.; Liu, X.F.; Lu, Z.Z.; Hung, T. Construction of an infectious clone of human adenovirus type 41. Arch. Virol. 2012, 157, 1313–1321. [Google Scholar] [CrossRef]
- Arad, U. Modified Hirt procedure for rapid purification of extrachromosomal DNA from mammalian cells. Biotechniques 1998, 24, 760–762. [Google Scholar] [CrossRef]
- Baer, A.; Kehn-Hall, K. Viral concentration determination through plaque assays: Using traditional and novel overlay systems. J. Vis. Exp. 2014. [Google Scholar] [CrossRef]
- Brauer, R.; Chen, P. Influenza virus propagation in embryonated chicken eggs. J. Vis. Exp. 2015. [Google Scholar] [CrossRef] [Green Version]
- Alemnesh, W.; Hair-Bejo, M.; Aini, I.; Omar, A.R. Pathogenicity of fowl adenovirus in specific pathogen free chicken embryos. J. Comp. Pathol. 2012, 146, 223–229. [Google Scholar] [CrossRef]
- Cowen, B.S. Chicken embryo propagation of type I avian adenoviruses. Avian Dis. 1988, 32, 347–352. [Google Scholar] [CrossRef]
- Abdul-Cader, M.S.; Palomino-Tapia, V.; Amarasinghe, A.; Ahmed-Hassan, H.; De Silva Senapathi, U.; Abdul-Careem, M.F. Hatchery Vaccination Against Poultry Viral Diseases: Potential Mechanisms and Limitations. Viral Immunol. 2018, 31, 23–33. [Google Scholar] [CrossRef]
- Pei, Y.; Corredor, J.C.; Krell, P.J.; Nagy, E. Fowl adenovirus 9 ORF19, a lipase homolog, is nonessential for virus replication and is suitable for foreign gene expression. Virus Res. 2019, 260, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Griffin, B.D.; Nagy, E. Coding potential and transcript analysis of fowl adenovirus 4: Insight into upstream ORFs as common sequence features in adenoviral transcripts. J. Gen. Virol. 2011, 92, 1260–1272. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tang, Q.; Qiu, L.; Yang, Z. Penton-dodecahedron of fowl adenovirus serotype 4 as a vaccine candidate for the control of related diseases. Vaccine 2019, 37, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Ruan, S.; Zhao, J.; Yin, X.; He, Z.; Zhang, G. A subunit vaccine based on fiber-2 protein provides full protection against fowl adenovirus serotype 4 and induces quicker and stronger immune responses than an inactivated oil-emulsion vaccine. Infect. Genet. Evol. 2018, 61, 145–150. [Google Scholar] [CrossRef]
- Chen, L.; Yin, L.; Zhou, Q.; Li, Q.; Luo, Y.; Xu, Z.; Zhang, Y.; Xue, C.; Cao, Y. Immunogenicity and protective efficacy of recombinant fiber-2 protein in protecting SPF chickens against fowl adenovirus 4. Vaccine 2018, 36, 1203–1208. [Google Scholar] [CrossRef]
- Grgic, H.; Poljak, Z.; Sharif, S.; Nagy, E. Pathogenicity and cytokine gene expression pattern of a serotype 4 fowl adenovirus isolate. PLoS ONE 2013, 8, e77601. [Google Scholar] [CrossRef]
- Michou, A.I.; Lehrmann, H.; Saltik, M.; Cotten, M. Mutational analysis of the avian adenovirus CELO, which provides a basis for gene delivery vectors. J. Virol. 1999, 73, 1399–1410. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Wang, J.; Gao, Y.; Cui, H.; Liu, C.; Qi, X.; Zhang, Y.; Wang, Y.; Wang, X. The Natural Large Genomic Deletion Is Unrelated to the Increased Virulence of the Novel Genotype Fowl Adenovirus 4 Recently Emerged in China. Viruses 2018, 10, 494. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, R.; Tian, K.; Wang, Z.; Yang, X.; Gao, D.; Zhang, Y.; Fu, J.; Wang, H.; Zhao, J. Fiber2 and hexon genes are closely associated with the virulence of the emerging and highly pathogenic fowl adenovirus 4. Emerg. Microbes Infect. 2018, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A., 3rd; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Yonemoto, I.T.; Weyman, P.D. Facile Site-Directed Mutagenesis of Large Constructs Using Gibson Isothermal DNA Assembly. Methods Mol. Biol. 2017, 1498, 359–366. [Google Scholar] [CrossRef]
- Lee, C.S.; Bishop, E.S.; Zhang, R.; Yu, X.; Farina, E.M.; Yan, S.; Zhao, C.; Zheng, Z.; Shu, Y.; Wu, X.; et al. Adenovirus-Mediated Gene Delivery: Potential Applications for Gene and Cell-Based Therapies in the New Era of Personalized Medicine. Genes Dis. 2017, 4, 43–63. [Google Scholar] [CrossRef]
- Mizuguchi, H.; Kay, M.A.; Hayakawa, T. Approaches for generating recombinant adenovirus vectors. Adv. Drug Deliv. Rev. 2001, 52, 165–176. [Google Scholar] [CrossRef]
- Chee, M.S.; Bankier, A.T.; Beck, S.; Bohni, R.; Brown, C.M.; Cerny, R.; Horsnell, T.; Hutchison, C.A., 3rd; Kouzarides, T.; Martignetti, J.A.; et al. Analysis of the protein-coding content of the sequence of human cytomegalovirus strain AD169. Curr. Top. Microbiol. Immunol. 1990, 154, 125–169. [Google Scholar] [CrossRef] [PubMed]
- Menard, C.; Wagner, M.; Ruzsics, Z.; Holak, K.; Brune, W.; Campbell, A.E.; Koszinowski, U.H. Role of murine cytomegalovirus US22 gene family members in replication in macrophages. J. Virol. 2003, 77, 5557–5570. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Iyer, L.M.; Aravind, L. A novel immunity system for bacterial nucleic acid degrading toxins and its recruitment in various eukaryotic and DNA viral systems. Nucleic Acids Res. 2011, 39, 4532–4552. [Google Scholar] [CrossRef] [Green Version]
- Zmasek, C.M.; Knipe, D.M.; Pellett, P.E.; Scheuermann, R.H. Classification of human Herpesviridae proteins using Domain-architecture Aware Inference of Orthologs (DAIO). Virology 2019, 529, 29–42. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fragment | Oligo Name | Sequence | Template | Product Length (bp) |
|---|---|---|---|---|
| AvrII-PacI | 1707Avr-PacF | cgaatacgagttggcctaggctctcgcagaacagggaatggggcattaattaaccgct | self-anneal | 96 |
| 1707Avr-PacR | tgtcgtacttcagccctaggccattggcggagaccgtaagcggttaattaatgcccca | |||
| AgeI-ORF1 | 1707KFAV4AgeIF | attcctccactgctttgaaccca | pKFAV4AP | 204 |
| 1707KFAV4AgeIR | cccgtaattgattactattaccttgtagaaaaagagagaaaattg | |||
| GFP cassette | 1707F02GFPF | ttctacaaggtaatagtaatcaattacggggtcattagtt | pAd5GFP | 1672 |
| 1707F02GFPR | tcgatttactgtgaagctacaagtgctagctaagatacattgatgagtttggacaaac | |||
| PvuI-ORF4 | 1711FAV4GCX5 | ggaaccgatcgaagaaagcaacag | pKFAV4-GFP | 424 |
| 1711FAV4GCX6 | gcccttgctcaccatgtcagaatatatagagaaaggaatgggc | |||
| mCherry CDS | 1711FAV4GCX7 | ctctatatattctgacatggtgagcaagggcgaggag | pmCherry-N1 | 742 |
| 1711FAV4GCX8 | ctggaatatagtgtgttacttgtacagctcgtccatgccg | |||
| ORF4-BamHI | 1711FAV4GCX9 | ggacgagctgtacaagtaacacactatattccagtccgaggagg | pKFAV4-GFP | 180 |
| 1711FAV4GCX10 | taagtggatccgcacaccattgc | 1281 | ||
| 117-bp linker | 1812FAV4APf | cgaaccagtaggcgaatacgagttggcct | pKFAV4AP | 117 |
| 1812FAV4APxAr | gggtattagtgtcgtacttcagcccAaggccattggcggagacc | |||
| 116-bp linker | 1812FAV4SAP1 | ccagtaggcgaatacgagttggcctaggctctcgcagaacagg | self-anneal | 116 |
| 1812FAV4SAP2 | tatctctatgctttgTTAATTAAcccattccctgttctgcgagagcctag g | |||
| 1812FAV4SAP3 | aatgggTTAATTAAcaaagcatagagataaaagaaacccgttactagtccagga | |||
| 1812FAV4SAP4 | ggatttaggaaaagtgtttcctggactagtaacgggtttctt | |||
| SpeI-BstZ17I | 1902FAdV4Xorf19a1 | caaagcatagagataaaagaaacccgttac ta | pKFAV4AP | 804 |
| 1902FAdV4Xorf19a2 | tacaacatttcagtagtttcctggtatacattgatgtgaccttcatggcgaac | |||
| CMVp | 1904FAV4MCHE1 | tcaagtgtatcatatgccaagtacg | pKFAV4-GFP | 395 |
| 1904FAV4MCHE2 | ccttgctcaccatggtaggcctctagcggatctgacggttcact | |||
| mCherry CDS | 1904FAV4MCHE3 | ctagaggcctaccatggtgagcaagggcgagga | pmCherry-N1 | 733 |
| 1904FAV4MCHE4 | ggccgtcgactacttgtacagctcgtccatgc | 1102 | ||
| mCherry | 1906mCHEF1 | ggccatcatcaaggagttcatg | 623 | |
| 1906mCHER1 | gttccacgatggtgtagtcctcg | |||
| GFP | 1906GFPf | cggccacaagttcagcgtgt c | 576 | |
| 1906GFPr | cgcttctcgttggggtcttt g | |||
| ORF2 | 1905ORF2f | ggcttcggaccgttactggg | 604 | |
| 1905ORF2r | gggggtacggttaatctccc | |||
| ORF19A/ORF4 | 1905ORF19Af | ctcccgttcaaagtagtgaa ca | 737 | |
| 1906ORF4r | ggcagatacagcacttcgcagta |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, B.; Zou, X.; Liu, X.; Zhao, J.; Zhang, W.; Guo, X.; Wang, M.; Lv, Y.; Lu, Z. User-Friendly Reverse Genetics System for Modification of the Right End of Fowl Adenovirus 4 Genome. Viruses 2020, 12, 301. https://doi.org/10.3390/v12030301
Yan B, Zou X, Liu X, Zhao J, Zhang W, Guo X, Wang M, Lv Y, Lu Z. User-Friendly Reverse Genetics System for Modification of the Right End of Fowl Adenovirus 4 Genome. Viruses. 2020; 12(3):301. https://doi.org/10.3390/v12030301
Chicago/Turabian StyleYan, Bingyu, Xiaohui Zou, Xinglong Liu, Jiaming Zhao, Wenfeng Zhang, Xiaojuan Guo, Min Wang, Yingtao Lv, and Zhuozhuang Lu. 2020. "User-Friendly Reverse Genetics System for Modification of the Right End of Fowl Adenovirus 4 Genome" Viruses 12, no. 3: 301. https://doi.org/10.3390/v12030301
APA StyleYan, B., Zou, X., Liu, X., Zhao, J., Zhang, W., Guo, X., Wang, M., Lv, Y., & Lu, Z. (2020). User-Friendly Reverse Genetics System for Modification of the Right End of Fowl Adenovirus 4 Genome. Viruses, 12(3), 301. https://doi.org/10.3390/v12030301