BoLA-DRB3 Polymorphism is Associated with Differential Susceptibility to Bovine Leukemia Virus-Induced Lymphoma and Proviral Load

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Diagnosis
2.2. BLV Proviral Load Determination
2.3. BoLA-DRB3 Genotyping
2.4. Detection of Anti-BLV gp51 Antibody by Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Association Study and Statistical Analysis
3. Results
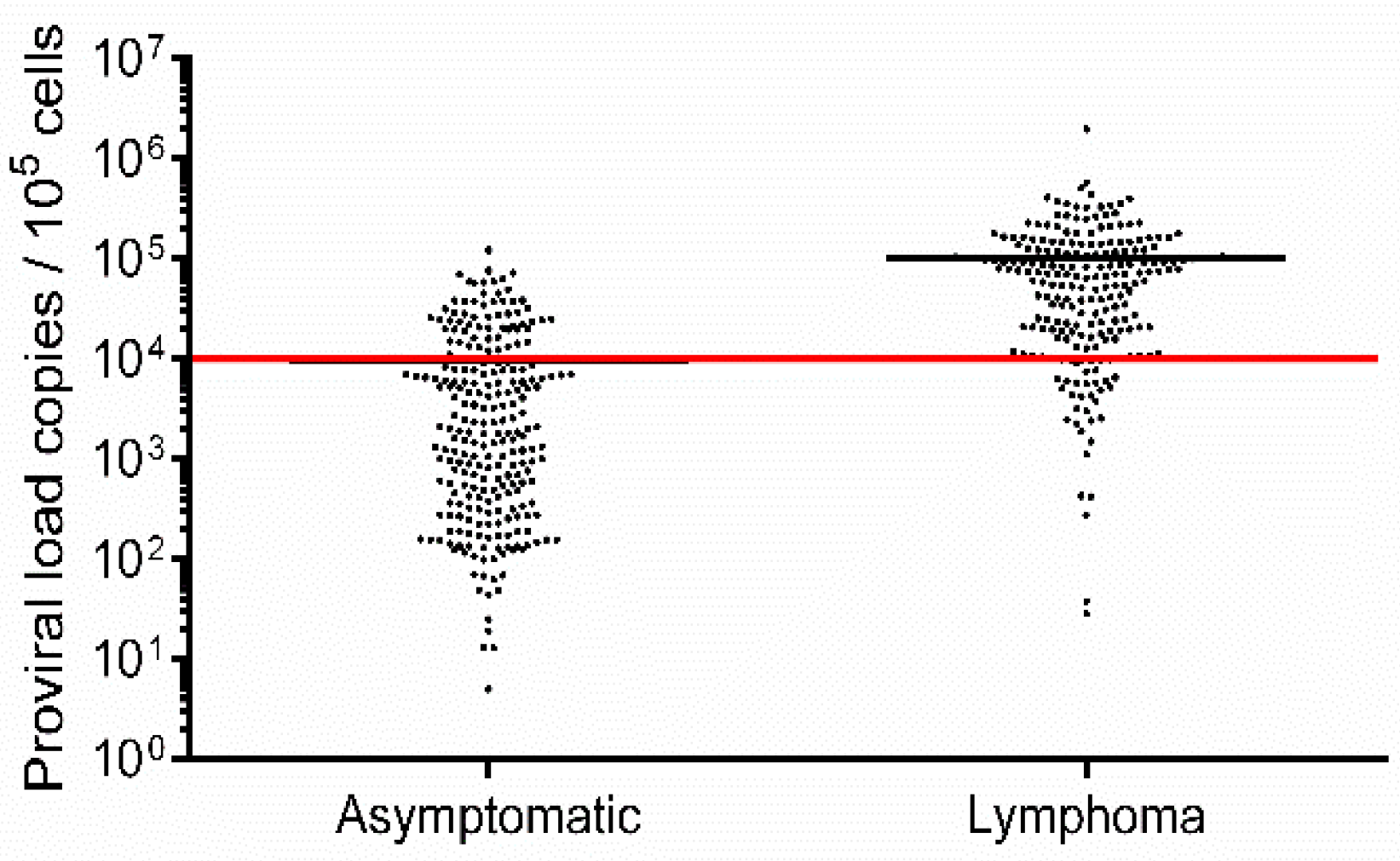
3.1. PVL Is Not Fully Correlated with Lymphoma Development
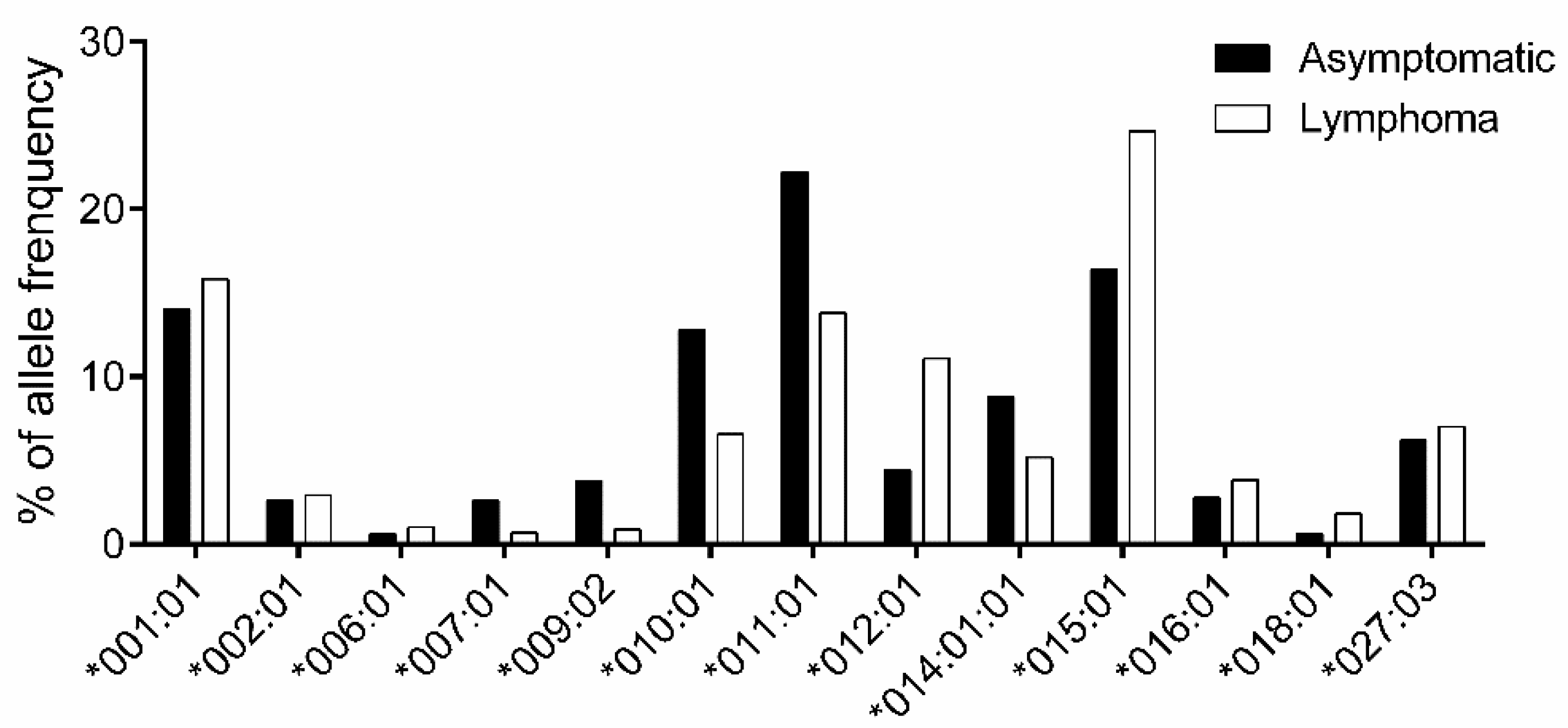
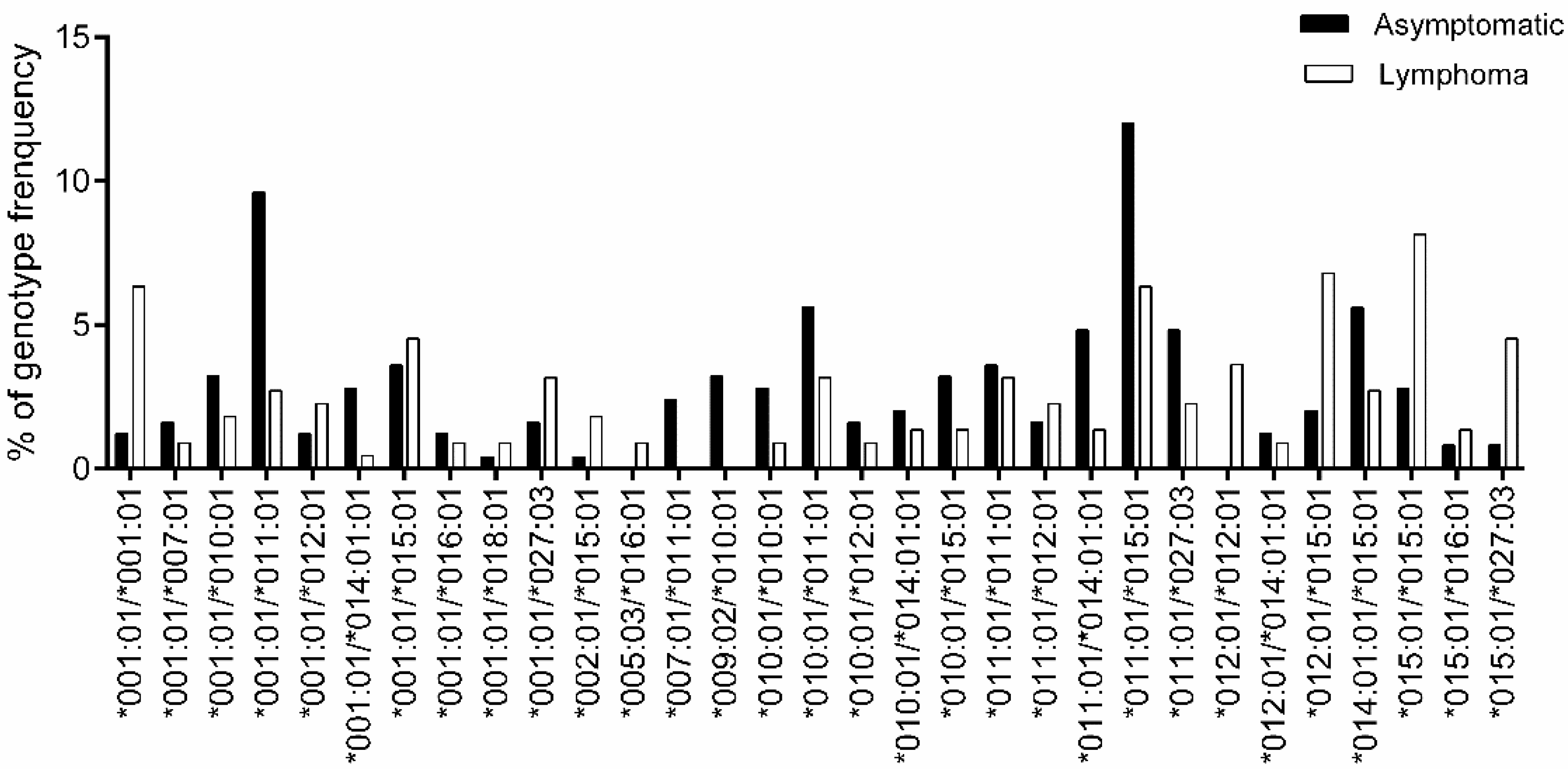
3.2. Association Study of BoLA-DRB3 with Lymphoma
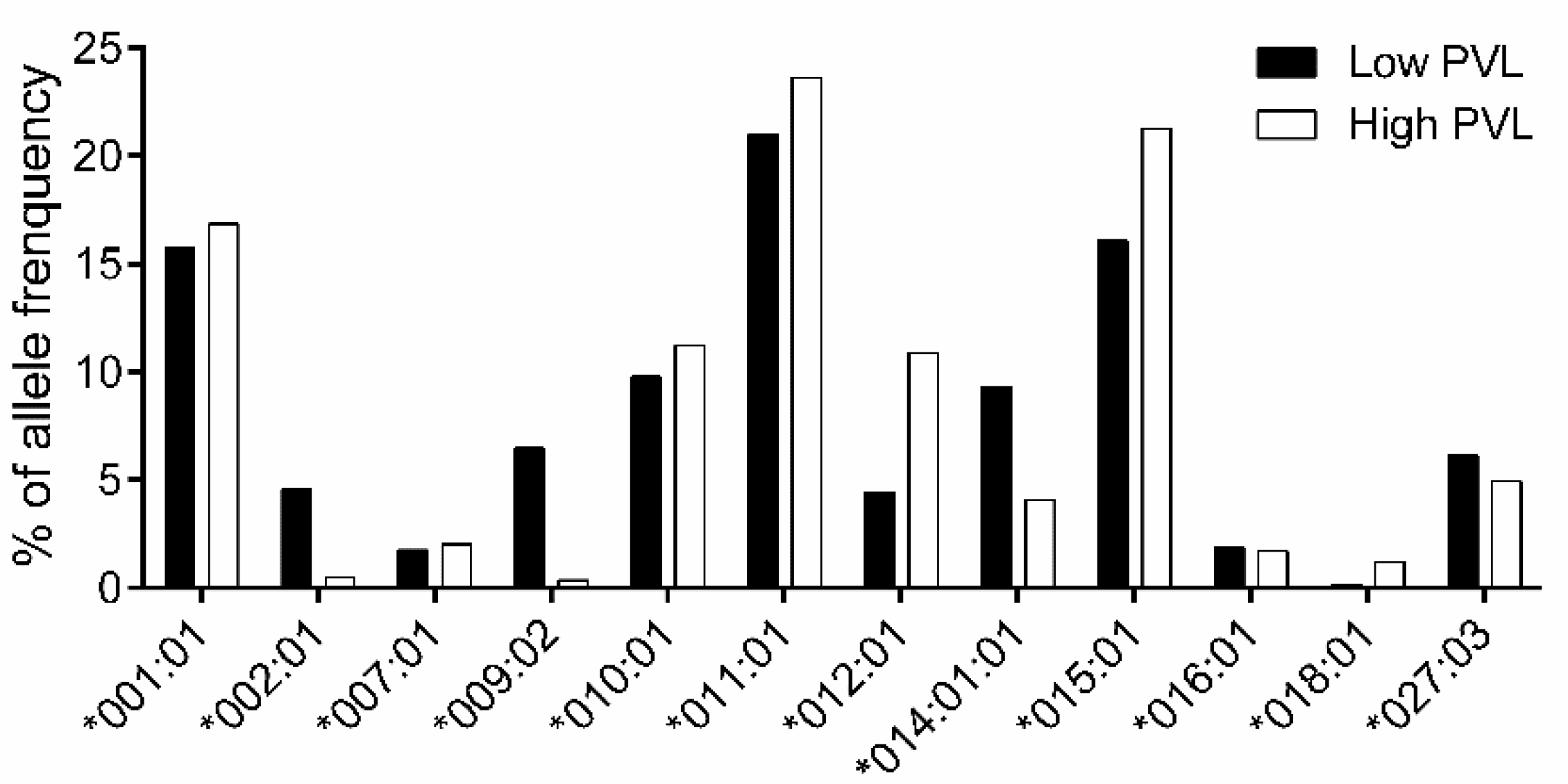
3.3. Association Study of BoLA-DRB3 with PVL
3.4. Differential Susceptibility of BoLA-DRB3 Polymorphisms to Lymphoma and PVL
3.5. BoLA-DRB3 Polymorphisms Are Associated with anti-BLV Antibody Production Levels
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hisada, M.; Chatterjee, N.; Kalaylioglu, Z.; Battjes, R.J.; Goedert, J.J. Hepatitis C virus load and survival among injection drug users in the united states. Hepatology 2005, 42, 1446–1452. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-J.; Yang, H.-I.; Iloeje, U.H.; Reveal-HBV Study Group. Hepatitis B virus DNA levels and outcomes in chronic hepatitis B. Hepatology 2009, 49, S72–S84. [Google Scholar] [CrossRef] [PubMed]
- Shoko, C.; Chikobvu, D. A superiority of viral load over cd4 cell count when predicting mortality in HIV patients on therapy. BMC Infect. Dis. 2019, 19, 169. [Google Scholar] [CrossRef]
- Iwanaga, M.; Watanabe, T.; Utsunomiya, A.; Okayama, A.; Uchimaru, K.; Koh, K.-R.; Ogata, M.; Kikuchi, H.; Sagara, Y.; Uozumi, K.; et al. Human T-cell leukemia virus type I (HTLV-1) proviral load and disease progression in asymptomatic HTLV-1 carriers: A nationwide prospective study in Japan. Blood 2010, 116, 1211–1219. [Google Scholar] [CrossRef] [Green Version]
- Furtado, M.D.S.B.S.; Andrade, R.G.; Romanelli, L.C.F.; Ribeiro, M.A.; Ribas, J.G.; Torres, E.B.; Barbosa-Stancioli, E.F.; Proietti, A.B.D.F.C.; Martins, M.L. Monitoring the HTLV-1 proviral load in the peripheral blood of asymptomatic carriers and patients with HTLV-associated myelopathy/tropical spastic paraparesis from a Brazilian cohort: Roc curve analysis to establish the threshold for risk disease. J. Med Virol. 2012, 84, 664–671. [Google Scholar] [CrossRef]
- Hong, F.; Jacobs, J.L.; Aga, E.; Cillo, A.R.; Fyne, E.; Koontz, D.L.; Zheng, L.; Mellors, J.W. Associations between HIV-1 DNA copy number, proviral transcriptional activity, and plasma viremia in individuals off or on suppressive antiretroviral therapy. Virology 2018, 521, 51–57. [Google Scholar] [CrossRef]
- Aida, Y.; Murakami, H.; Takahashi, M.; Takeshima, S.-N. Mechanisms of pathogenesis induced by bovine leukemia virus as a model for human T-cell leukemia virus. Front. Microbiol. 2013, 4, 328. [Google Scholar] [CrossRef] [Green Version]
- Gillet, N.; Florins, A.; Boxus, M.; Burteau, C.; Nigro, A.; Vandermeers, F.; Balon, H.; Bouzar, A.-B.; Defoiche, J.; Burny, A.; et al. Mechanisms of leukemogenesis induced by bovine leukemia virus: Prospects for novel anti-retroviral therapies in human. Retrovirology 2007, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Panei, C.J.; Takeshima, S.-N.; Omori, T.; Nunoya, T.; Davis, W.C.; Ishizaki, H.; Matoba, K.; Aida, Y. Estimation of bovine leukemia virus (BLV) proviral load harbored by lymphocyte subpopulations in BLV-infected cattle at the subclinical stage of enzootic bovine leucosis using BLV-cocomo-qPCR. BMC Vet. Res. 2013, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Jimba, M.; Takeshima, S.-N.; Murakami, H.; Kohara, J.; Kobayashi, N.; Matsuhashi, T.; Ohmori, T.; Nunoya, T.; Aida, Y. BLV-cocomo-qPCR: A useful tool for evaluating bovine leukemia virus infection status. BMC Vet. Res. 2012, 8, 167. [Google Scholar] [CrossRef] [Green Version]
- Somura, Y.; Sugiyama, E.; Fujikawa, H.; Murakami, K. Comparison of the copy numbers of bovine leukemia virus in the lymph nodes of cattle with enzootic bovine leukosis and cattle with latent infection. Arch. Virol. 2014, 159, 2693–2697. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Inagaki, Y.; Ohnuki, N.; Sato, R.; Murakami, S.; Imakawa, K. Increasing bovine leukemia virus (BLV) proviral load is a risk factor for progression of enzootic bovine leucosis: A prospective study in Japan. Prev. Vet. Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ohno, A.; Takeshima, S.N.; Matsumoto, Y.; Aida, Y. Risk factors associated with increased bovine leukemia virus proviral load in infected cattle in Japan from 2012 to 2014. Virus Res 2015, 210, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, S.-N.; Aida, Y. Structure, function and disease susceptibility of the bovine major histocompatibility complex. Anim. Sci. J. 2006, 77, 138–150. [Google Scholar] [CrossRef]
- Garrick, D.J.; Ruvinsky, A. The Genetics of Cattle; International, C.A.B.: Wallingford, Oxfordshire, UK; Boston, MA, USA, 2015. [Google Scholar]
- Panei, C.J.; Suzuki, K.; Echeverría, M.G.; Serena, M.S.; Metz, G.E.; González, E.T. Association of BOLA-DRB3.2 alleles with resistance and susceptibility to persistent lymphocytosis in BLV infected cattle in Argentina. Int. J. Dairy Sci. 2009, 4, 123–128. [Google Scholar]
- Nikbakht Brujeni, G.; Ghorbanpour, R.; Esmailnejad, A. Association of BOLA-DRB3.2 alleles with BLV infection profiles (persistent lymphocytosis/lymphosarcoma) and lymphocyte subsets in Iranian Holstein cattle. Biochem. Genet. 2016, 54, 194–207. [Google Scholar] [CrossRef]
- Juliarena, M.; Poli, M.; Sala, L.; Ceriani, C.; Gutierrez, S.; Dolcini, G.L.; Rodriguez, E.; Mariño, B.; Rodríguez-Dubra, C.; Esteban, E.N. Association of BLV infection profiles with alleles of the BOLA-DRB3.2 gene. Anim. Genet. 2008, 39, 432–438. [Google Scholar] [CrossRef]
- Takeshima, S.-N.; Ohno, A.; Aida, Y. Bovine leukemia virus proviral load is more strongly associated with bovine major histocompatibility complex class II drb3 polymorphism than with dqa1 polymorphism in holstein cow in Japan. Retrovirology 2019, 16, 14. [Google Scholar] [CrossRef]
- Miyasaka, T.; Takeshima, S.N.; Jimba, M.; Matsumoto, Y.; Kobayashi, N.; Matsuhashi, T.; Sentsui, H.; Aida, Y. Identification of bovine leukocyte antigen class II haplotypes associated with variations in bovine leukemia virus proviral load in Japanese black cattle. Tissue Antigens 2013, 81, 72–82. [Google Scholar] [CrossRef]
- Nieto Farias, M.; Caffaro, M.; Pamela, L.; Passucci, J.; Poli, M.; Ceriani, C.; Dolcini, G.L. A novel association of BOLA DRB3 alleles in blv infected cattle with different proviral loads. Braz. J. Vet. Res. Anim. Sci. 2017, 54, 215. [Google Scholar] [CrossRef] [Green Version]
- Konnai, S.; Takeshima, S.-N.; Tajima, S.; Yin, S.A.; Okada, K.; Onuma, M.; Aida, Y. The influence of ovine MHC class II DRB1 alleles on immune response in bovine leukemia virus infection. Microbiol. Immunol. 2003, 47, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Rosewick, N.; Durkin, K.; Artesi, M.; Marçais, A.; Hahaut, V.; Griebel, P.; Arsic, N.; Avettand-Fenoel, V.; Burny, A.; Charlier, C.; et al. Cis-perturbation of cancer drivers by the HTLV-1/BLV proviruses is an early determinant of leukemogenesis. Nat. Commun. 2017, 8, 15264. [Google Scholar] [CrossRef]
- Thibodeau, J.; Bourgeois-Daigneault, M.-C.; Lapointe, R. Targeting the MHC class II antigen presentation pathway in cancer immunotherapy. Oncoimmunology 2012, 1, 908–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldford, S.A.; Robb, J.D.; Watson, P.H.; Drover, S. HLA-DRB alleles are differentially expressed by tumor cells in breast carcinoma. Int. J. Cancer 2004, 112, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Marty, R.; Kaabinejadian, S.; Rossell, D.; Slifker, M.J.; van de Haar, J.; Engin, H.B.; de Prisco, N.; Ideker, T.; Hildebrand, W.H.; Font-Burgada, J.; et al. MHC-I genotype restricts the oncogenic mutational landscape. Cell 2017, 171, 1272–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florins, A.; Gillet, N.; Boxus, M.; Kerkhofs, P.; Kettmann, R.; Willems, L. Even attenuated bovine leukemia virus proviruses can be pathogenic in sheep. J. Virol. 2007, 81, 10195–10200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimba, M.; Takeshima, S.-N.; Matoba, K.; Endoh, D.; Aida, Y. Blv-cocomo-qpcr: Quantitation of bovine leukemia virus proviral load using the cocomo algorithm. Retrovirology 2010, 7, 91. [Google Scholar] [CrossRef] [Green Version]
- Takeshima, S.N.; Kitamura-Muramatsu, Y.; Yuan, Y.; Polat, M.; Saito, S.; Aida, Y. BLV-cocomo-qPCR-2: Improvements to the BLV-cocomo-qPCR assay for bovine leukemia virus by reducing primer degeneracy and constructing an optimal standard curve. Arch. Virol. 2015, 160, 1325–1332. [Google Scholar] [CrossRef]
- Yuan, Y.; Kitamura-Muramatsu, Y.; Saito, S.; Ishizaki, H.; Nakano, M.; Haga, S.; Matoba, K.; Ohno, A.; Murakami, H.; Takeshima, S.N.; et al. Detection of the BLV provirus from nasal secretion and saliva samples using BLV-cocomo-qPCR-2: Comparison with blood samples from the same cattle. Virus Res. 2015, 210, 248–254. [Google Scholar] [CrossRef]
- Takeshima, S.N.; Matsumoto, Y.; Miyasaka, T.; Arainga-Ramirez, M.; Saito, H.; Onuma, M.; Aida, Y. A new method for typing bovine major histocompatibility complex class II DRB3 alleles by combining two established PCR sequence-based techniques. Tissue Antigens 2011, 78, 208–213. [Google Scholar] [CrossRef]
- Cordell, H.J. Detecting gene-gene interactions that underlie human diseases. Nat. Rev. Genet. 2009, 10, 392–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanuki, S.; Takeshima, S.-N.; Borjigin, L.; Sato, H.; Bai, L.; Murakami, H.; Sato, R.; Ishizaki, H.; Matsumoto, Y.; Aida, Y. Visualizing bovine leukemia virus (BLV)-infected cells and measuring BLV proviral loads in the milk of BLV seropositive dams. Vet. Res. 2019, 50, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forletti, A.; Lutzelschwab, C.M.; Cepeda, R.; Esteban, E.N.; Gutierrez, S.E. Early events following bovine leukaemia virus infection in calves with different alleles of the major histocompatibility complex DRB3 gene. Vet. Res. 2020, 51, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debacq, C.; Asquith, B.; Reichert, M.; Burny, A.; Kettmann, R.; Willems, L. Reduced cell turnover in bovine leukemia virus-infected, persistently lymphocytotic cattle. J. Virol. 2003, 77, 13073–13083. [Google Scholar] [CrossRef] [Green Version]
- Kettmann, R.; Deschamps, J.; Cleuter, Y.; Couez, D.; Burny, A.; Marbaix, G. Leukemogenesis by bovine leukemia virus: Proviral DNA integration and lack of rna expression of viral long terminal repeat and 3’ proximate cellular sequences. Proc. Natl. Acad. Sci. USA 1982, 79, 2465–2469. [Google Scholar] [CrossRef] [Green Version]
- Gillet, N.A.; Gutiérrez, G.; Rodriguez, S.M.; de Brogniez, A.; Renotte, N.; Alvarez, I.; Trono, K.; Willems, L. Massive depletion of bovine leukemia virus proviral clones located in genomic transcriptionally active sites during primary infection. PLoS Pathog. 2013, 9, e1003687. [Google Scholar] [CrossRef]
- Willems, L.; Kerkhofs, P.; Dequiedt, F.; Portetelle, D.; Mammerickx, M.; Burny, A.; Kettmann, R. Attenuation of bovine leukemia virus by deletion of R3 and G4 open reading frames. Proc. Natl. Acad. Sci. USA 1994, 91, 11532–11536. [Google Scholar] [CrossRef] [Green Version]
- Dequiedt, F.; Kettmann, R.; Burny, A.; Willems, L. Mutations in the p53 tumor-suppressor gene are frequently associated with bovine leukemia virus-induced leukemogenesis in cattle but not in sheep. Virology 1995, 209, 676–683. [Google Scholar] [CrossRef]
- Tajima, S.; Zhuang, W.Z.; Kato, M.V.; Okada, K.; Ikawa, Y.; Aida, Y. Function and conformation of wild-type p53 protein are influenced by mutations in bovine leukemia virus-induced b-cell lymphosarcoma. Virology 1998, 243, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Konnai, S.; Usui, T.; Ikeda, M.; Kohara, J.; Hirata, T.-I.; Okada, K.; Ohashi, K.; Onuma, M. Tumor necrosis factor-alpha genetic polymorphism may contribute to progression of bovine leukemia virus-infection. Microbes Infect. 2006, 8, 2163–2171. [Google Scholar] [CrossRef] [Green Version]
- Bugawan, T.L.; Mack, S.J.; Stoneking, M.; Saha, M.; Beck, H.P.; Erlich, H.A. Hla class I allele distributions in six pacific/Asian populations: Evidence of selection at the HLA-a locus. Tissue Antigens 1999, 53, 311–319. [Google Scholar] [CrossRef]
- Velickovic, Z.M.; Delahunt, B.; Carter, J.M. HLA-DRB1 and HLA-DQB1 polymorphisms in Pacific islands populations. Tissue Antigens 2002, 59, 397–406. [Google Scholar] [CrossRef]
- Miyasaka, T.; Takeshima, S.N.; Matsumoto, Y.; Kobayashi, N.; Matsuhashi, T.; Miyazaki, Y.; Tanabe, Y.; Ishibashi, K.; Sentsui, H.; Aida, Y. The diversity of bovine MHC class II drb3 and dqa1 alleles in different herds of Japanese black and Holstein cattle in Japan. Gene 2011, 472, 42–49. [Google Scholar] [CrossRef]
- Takeshima, S.; Chen, S.; Miki, M.; Kado, M.; Aida, Y. Distribution and origin of bovine major histocompatibility complex class II DQA1 genes in Japan. Tissue Antigens 2008, 72, 195–205. [Google Scholar] [CrossRef]
- Takeshima, S.; Nakai, Y.; Ohta, M.; Aida, Y. Short communication: Characterization of DRB3 alleles in the MHC of Japanese shorthorn cattle by polymerase chain reaction-sequence-based typing. J. Dairy Sci. 2002, 85, 1630–1632. [Google Scholar] [CrossRef]
- Takeshima, S.; Saitou, N.; Morita, M.; Inoko, H.; Aida, Y. The diversity of bovine MHC class II drb3 genes in Japanese black, Japanese shorthorn, Jersey and Holstein cattle in Japan. Gene 2003, 316, 111–118. [Google Scholar] [CrossRef]
- Pineda, M.V.; Bouzas, M.B.; Remesar, M.; Fridman, A.; Remondegui, C.; Mammana, L.; Altamirano, N.; Paradiso, P.; Costantini, P.; Tadey, L.; et al. Relevance of HTLV-1 proviral load in asymptomatic and symptomatic patients living in endemic and non-endemic areas of argentina. PLoS ONE 2019, 14, e0225596. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Status | Asymptomatic (n = 250) | Lymphoma (n = 221) |
|---|---|---|
| Low proviral load 1 | 188 | 37 |
| High proviral load 2 | 62 | 184 |
| BoLA-DRB3 Allele | Asymptomatic (250 Cattle) | Lymphoma (221 Cattle) | OR | p-Value | p-Value Rank (I) | BH Value (I/Allele Number)*0.05 | Susceptibility |
|---|---|---|---|---|---|---|---|
| *001:01 | 70 | 70 | 1.156 | 0.463 | - | ||
| *002:01 | 13 | 13 | 1.135 | 1.000 | - | ||
| *006:01 | 3 | 3 | 1.132 | 1.000 | - | ||
| *007:01 | 13 | 3 | 0.256 | 0.024 | 6 | 0.009 | - |
| *009:02 | 19 | 4 | 0.231 | 0.005 | 5 | 0.008 | R |
| *010:01 | 64 | 29 | 0.478 | 0.001 | 2 | 0.003 | R |
| *011:01 | 111 | 61 | 0.561 | 0.001 | 2 | 0.003 | R |
| *012:01 | 22 | 49 | 2.709 | 0.000 | 1 | 0.002 | S |
| *014:01:01 | 44 | 23 | 0.569 | 0.042 | - | ||
| *015:01 | 82 | 109 | 1.669 | 0.002 | 4 | 0.006 | S |
| *016:01 | 14 | 17 | 1.389 | 0.578 | - | ||
| *018:01 | 3 | 8 | 3.054 | 0.127 | - | ||
| *027:03 | 31 | 31 | 1.141 | 0.693 | - |
| BoLA-DRB3 Allele | Univariate | Multivariate | ||||||
|---|---|---|---|---|---|---|---|---|
| p-Value | OR | L95 | U95 | p-Value | OR | L95 1 | U95 2 | |
| *009:02 | 0.002 | 0.10 | 0.02 | 0.43 | 4.27 × 10−4 | 0.07 | 0.01 | 0.30 |
| *010:01 | 0.008 | 0.52 | 0.32 | 0.84 | 7.38 × 10−4 | 0.43 | 0.026 | 0.70 |
| *011:01 | 9.91 × 10−4 | 0.53 | 0.36 | 0.77 | 5.77 × 10−6 | 0.40 | 0.27 | 0.59 |
| *014:01:01 | 0.026 | 0.53 | 0.30 | 0.93 | 7.82 × 10−4 | 0.36 | 0.20 | 0.66 |
| BoLA-DRB3 Genotype | Asymptomatic (212 Cattle) | Lymphoma (221 Cattle) | OR | p-Value | p-Value Rank (I) | BH Value (I/Genotype Number)*0.05 | Susceptibility |
|---|---|---|---|---|---|---|---|
| *001:01/*001:01 | 3 | 14 | 5.568 | 0.005 | - | ||
| *001:01/*007:01 | 4 | 2 | 0.562 | 0.689 | - | ||
| *001:01/*010:01 | 8 | 4 | 0.558 | 0.393 | - | ||
| *001:01/*011:01 | 24 | 6 | 0.263 | 0.002 | 1 | 0.0005 | - |
| *001:01/*012:01 | 3 | 5 | 1.906 | 0.483 | - | ||
| *001:01/*014:01:01 | 7 | 1 | 0.158 | 0.393 | - | ||
| *001:01/*015:01 | 9 | 10 | 1.269 | 0.645 | - | ||
| *001:01/* 016:01 | 3 | 2 | 0.752 | 1.000 | - | ||
| *001:01/*018:01 | 1 | 2 | 2.274 | 0.603 | - | ||
| *001:01/*027:03 | 4 | 7 | 2.012 | 0.362 | - | ||
| *002:01/*015:01 | 1 | 4 | 4.590 | 0.191 | - | ||
| *005:03/*016:01 | 0 | 2 | - | 0.220 | - | ||
| *007:01/*011:01 | 6 | 0 | 0 | 0.057 | - | ||
| *009:02/*010:01 | 8 | 0 | - | 0.008 | - | ||
| *010:01/*010:01 | 7 | 2 | 0.317 | 0.183 | - | ||
| *010:01/*011:01 | 14 | 7 | 0.551 | 0.264 | - | ||
| *010:01/*012:01 | 4 | 2 | 0.562 | 0.689 | - | ||
| *010:01/*014:01:01 | 5 | 3 | 0.674 | 0.728 | - | ||
| *010:01/*015:01 | 8 | 3 | 0.416 | 0.231 | - | ||
| *011:01/*011:01 | 9 | 7 | 0.876 | 1.000 | - | ||
| *011:01/*012:01 | 4 | 5 | 1.424 | 0.740 | - | ||
| *011:01/*014:01:01 | 12 | 3 | 0.273 | 0.037 | - | ||
| *011:01/*015:01 | 30 | 14 | 0.496 | 0.039 | - | ||
| *011:01/*027:03 | 12 | 5 | 0.459 | 0.215 | - | ||
| *012:01/*012:01 | 0 | 8 | - | 0.002 | 1 | 0.0005 | - |
| *012:01/*014:01:01 | 3 | 2 | 0.752 | 1.000 | - | ||
| *012:01/*015:01 | 5 | 15 | 3.568 | 0.011 | - | ||
| *014:01:01/*015:01 | 4 | 1 | 0.280 | 0.377 | - | ||
| *015:01/*015:01 | 7 | 18 | 3.078 | 0.013 | - | ||
| *015:01/*016:01 | 2 | 3 | 1.706 | 0.669 | - | ||
| *015:01/*027:03 | 2 | 10 | 5.877 | 0.016 | - |
| BoLA-DRB3 Allele | Low PVL (317 Cattle) | High PVL (294 Cattle) | OR | p-Value | p-Value Rank (I) | BH Value (I/Genotype Number)*0.05 | Susceptibility |
|---|---|---|---|---|---|---|---|
| *001:01 | 100 | 99 | 1.8681 | 0.6046 | - | ||
| *002:01 | 29 | 3 | 0.1458 | < 0.0001 | 1 | 0.0019 | R |
| *007:01 | 11 | 12 | 1.5744 | 0.8322 | - | ||
| *009:02 | 41 | 2 | 0.0685 | < 0.0001 | 1 | 0.0019 | R |
| *010:01 | 62 | 66 | 1.7989 | 0.4262 | - | ||
| *011:01 | 133 | 139 | 2.2967 | 0.1933 | - | ||
| *012:01 | 28 | 64 | 3.8383 | < 0.0001 | 1 | 0.0019 | S |
| *014:01:01 | 59 | 24 | 0.6068 | 0.0004 | 4 | 0.0077 | R |
| *015:01 | 102 | 125 | 2.5463 | 0.0224 | 5 | 0.0096 | - |
| *016:01 | 12 | 10 | 1.1962 | 0.8314 | - | ||
| *018:01 | 1 | 7 | 9.968 | 0.0319 | - | ||
| *027:03 | 39 | 29 | 1.1249 | 0.369 | - |
| BoLA-DRB3 Allele | Univariate | Multivariate | ||||||
|---|---|---|---|---|---|---|---|---|
| p-Value | OR | L95 | U95 | p-Value | OR | L95 | U95 | |
| *002:01 | 6.28 × 10−4 | 0.13 | 0.04 | 0.41 | 5.19 × 10−4 | 0.12 | 0.04 | 0.40 |
| *009:02 | 3.29 × 10−5 | 0.05 | 0.01 | 0.20 | 1.42 × 10−5 | 0.04 | 0.01 | 0.17 |
| *012:01 | 4.00 × 10−5 | 2.65 | 1.66 | 4.22 | 3.20 × 10−4 | 2.51 | 1.52 | 4.15 |
| *014:01:01 | 2.40 × 10−4 | 0.39 | 0.23 | 0.64 | 2.10 × 10−5 | 0.31 | 0.18 | 0.53 |
| BoLA-DRB3 Genotype | Low PVL (317 Cattle) | High PVL (294 Cattle) | OR | p-Value | p-Value Rank (I) | BH Value (I/Genotype Number)*0.05 | Susceptibility |
|---|---|---|---|---|---|---|---|
| *001:01/*001:01 | 4 | 13 | 3.6201 | 0.0245 | - | ||
| *001:01/*002:01 | 10 | 0 | 0.0000 | 0.0019 | 3 | 0.0016 | - |
| *001:01/*009:02 | 5 | 0 | 0.0000 | 0.0624 | - | ||
| *001:01/*010:01 | 7 | 8 | 1.2388 | 0.7958 | - | ||
| *001:01/*011:01 | 27 | 20 | 0.7840 | 0.4511 | - | ||
| *001:01/*012:01 | 5 | 9 | 1.9705 | 0.2824 | - | ||
| *001:01/*014:01:01 | 8 | 3 | 0.3982 | 0.2260 | - | ||
| *001:01/*015:01 | 15 | 20 | 1.4696 | 0.2990 | - | ||
| *001:01/*027:03 | 9 | 6 | 0.7130 | 0.6067 | - | ||
| *002:01/*011:01 | 4 | 1 | 0.2671 | 0.3747 | - | ||
| *007:01/*011:01 | 3 | 5 | 1.8108 | 0.4908 | - | ||
| *007:01/*012:01 | 1 | 3 | 3.2577 | 0.3559 | - | ||
| *009:02/*010:01 | 7 | 0 | 0.0000 | 0.0156 | - | ||
| *009:02/*012:01 | 5 | 0 | 0.0000 | 0.0624 | - | ||
| *009:02/*015:01 | 13 | 0 | 0.0000 | 0.0002 | 1 | 0.0005 | R |
| *010:01/*010:01 | 7 | 5 | 0.7662 | 0.7741 | - | ||
| *010:01/*011:01 | 9 | 24 | 3.0420 | 0.0039 | - | ||
| *010:01/*012:01 | 3 | 4 | 1.4437 | 0.7160 | - | ||
| *010:01/*014:01:01 | 4 | 1 | 0.2671 | 0.3747 | - | ||
| *010:01/*015:01 | 8 | 10 | 1.3600 | 0.6341 | - | ||
| *010:01/*016:01 | 3 | 2 | 0.7169 | 1.0000 | - | ||
| *010:01/*027:03 | 3 | 3 | 1.0790 | 1.0000 | - | ||
| *011:01/*011:01 | 17 | 11 | 0.6859 | 0.4393 | - | ||
| *011:01/*012:01 | 3 | 18 | 6.8261 | 0.0005 | 2 | 0.0011 | S |
| *011:01/*014:01:01 | 14 | 7 | 0.5279 | 0.1883 | - | ||
| *011:01/*015:01 | 19 | 32 | 1.9156 | 0.0396 | - | ||
| *011:01/*027:03 | 12 | 6 | 0.5295 | 0.2370 | - | ||
| *012:01/*012:01 | 0 | 4 | - | 0.0530 | - | ||
| *012:01/*014:01:01 | 6 | 2 | 0.3550 | 0.2886 | - | ||
| *012:01/*015:01 | 2 | 12 | 6.7021 | 0.0053 | - | ||
| *012:01/*027:03 | 2 | 4 | 2.1724 | 0.4351 | - | ||
| *014:01:01/*015:01 | 18 | 6 | 0.3461 | 0.0223 | - | ||
| *014:01:01/*027:03 | 3 | 2 | 0.7169 | 1.0000 | - | ||
| *015:01/*015:01 | 6 | 19 | 3.5812 | 0.0067 | - | ||
| *015:01/*016:01 | 3 | 2 | 0.7169 | 1.0000 | - | ||
| *015:01/*027:03 | 3 | 4 | 1.4437 | 0.7160 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo, C.-W.; Borjigin, L.; Saito, S.; Fukunaga, K.; Saitou, E.; Okazaki, K.; Mizutani, T.; Wada, S.; Takeshima, S.-n.; Aida, Y. BoLA-DRB3 Polymorphism is Associated with Differential Susceptibility to Bovine Leukemia Virus-Induced Lymphoma and Proviral Load. Viruses 2020, 12, 352. https://doi.org/10.3390/v12030352
Lo C-W, Borjigin L, Saito S, Fukunaga K, Saitou E, Okazaki K, Mizutani T, Wada S, Takeshima S-n, Aida Y. BoLA-DRB3 Polymorphism is Associated with Differential Susceptibility to Bovine Leukemia Virus-Induced Lymphoma and Proviral Load. Viruses. 2020; 12(3):352. https://doi.org/10.3390/v12030352
Chicago/Turabian StyleLo, Chieh-Wen, Liushiqi Borjigin, Susumu Saito, Koya Fukunaga, Etsuko Saitou, Katsunori Okazaki, Tetsuya Mizutani, Satoshi Wada, Shin-nosuke Takeshima, and Yoko Aida. 2020. "BoLA-DRB3 Polymorphism is Associated with Differential Susceptibility to Bovine Leukemia Virus-Induced Lymphoma and Proviral Load" Viruses 12, no. 3: 352. https://doi.org/10.3390/v12030352
APA StyleLo, C.-W., Borjigin, L., Saito, S., Fukunaga, K., Saitou, E., Okazaki, K., Mizutani, T., Wada, S., Takeshima, S.-n., & Aida, Y. (2020). BoLA-DRB3 Polymorphism is Associated with Differential Susceptibility to Bovine Leukemia Virus-Induced Lymphoma and Proviral Load. Viruses, 12(3), 352. https://doi.org/10.3390/v12030352