Virus-Like Particle-Mediated Vaccination against Interleukin-13 May Harbour General Anti-Allergic Potential beyond Atopic Dermatitis

Abstract
:1. Introduction: Why Develop an Anti-IL-13 Vaccine?
2. Virus-Like Particles as a Construct for IL-13 Therapeutic Vaccine
3. The Health Economics of IL-13-Targetable Diseases
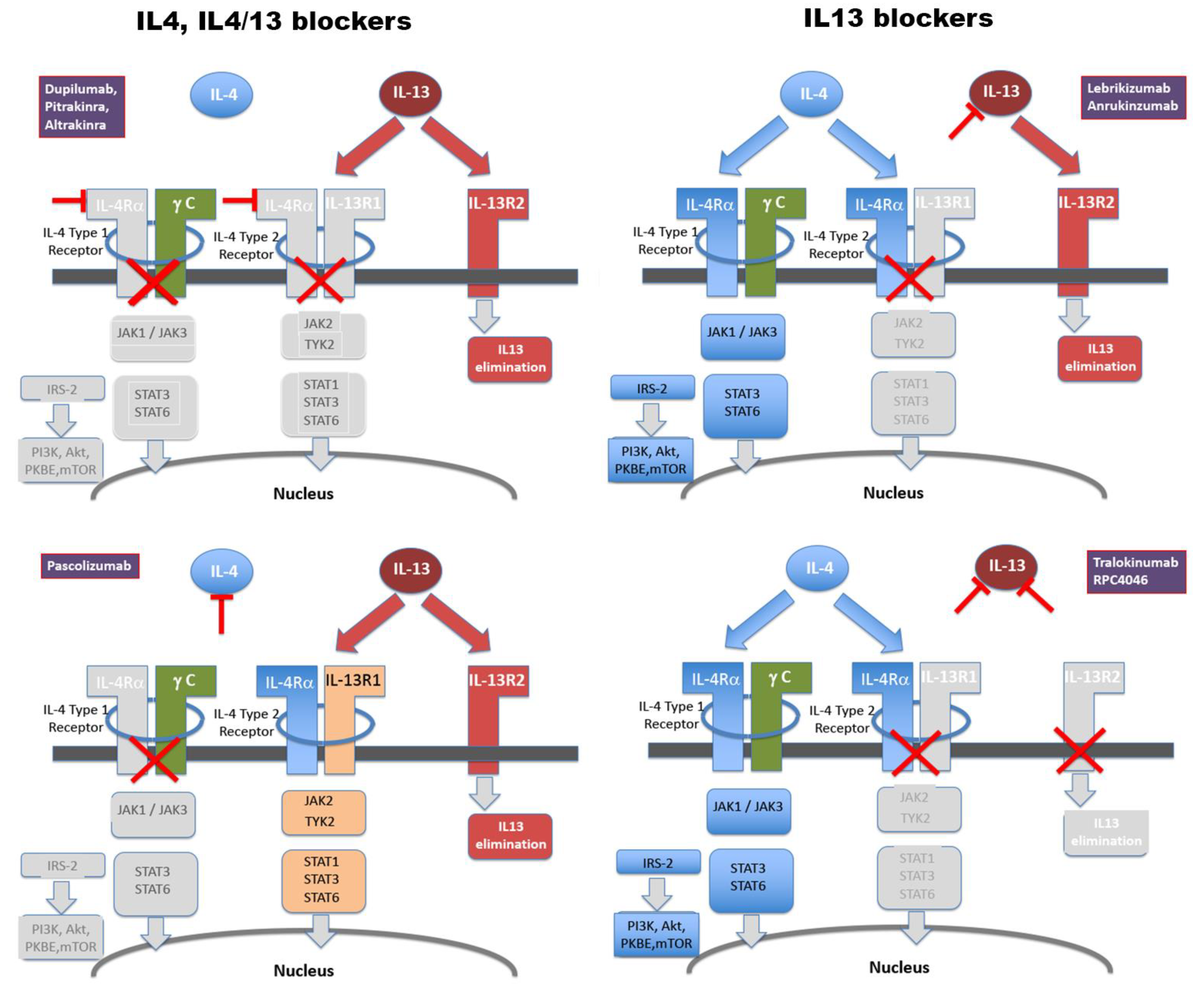
4. Monoclonal Antibodies vs. Polyclonal Vaccine Responses
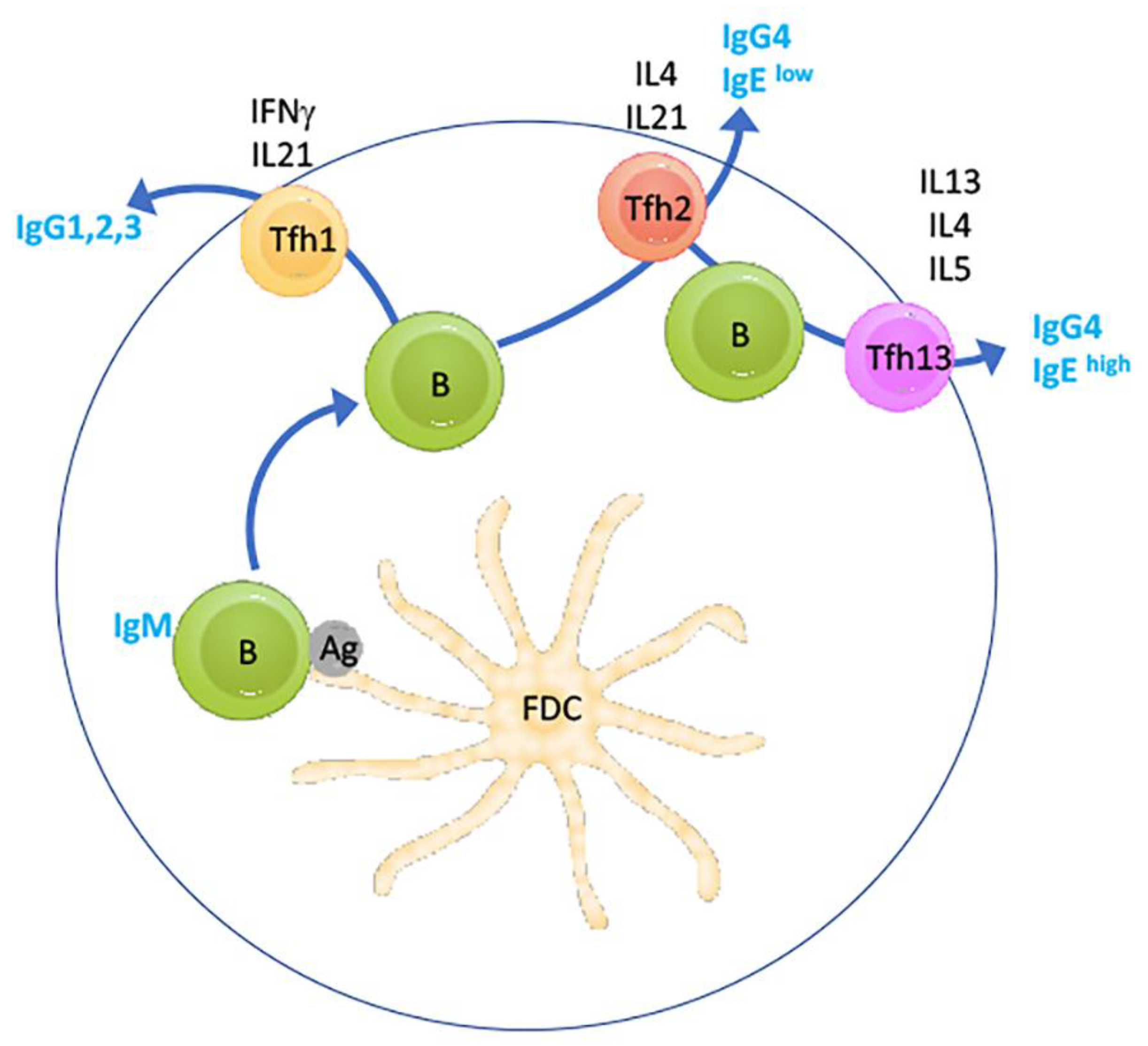
5. The Inhibition of Anaphylactic TfH Cells: A Novel Role for IL-13 Neutralisation
6. Safety of Anti-IL-13 Vaccination
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bao, K.; Reinhardt, R.L. The differential expression of IL-4 and IL-13 and its impact on type-2 immunity. Cytokine 2015, 75, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Junttila, I. Tuning the Cytokine Responses: An Update on Interleukin (IL)-4 and IL-13 Receptor Complexes. Front. Immunol. 2018, 9, 888. [Google Scholar] [CrossRef] [PubMed]
- LaPorte, S.L.; Juo, Z.S.; Vaclavikova, J.; Colf, L.A.; Qi, X.; Heller, N.M.; Keegan, A.D.; Garcia, K.C. Molecular and structural basis of cytokine receptor pleiotropy in the interleukin-4/13 system. Cell 2008, 132, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimica, V.; Galarza, J.M. Adjuvant formulations for virus-like particle (VLP) based vaccines. Clin. Immunol. 2017, 183, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Fuenmayor, J.; Gòdia, F.; Cervera, L. Production of virus-like particles for vaccines. N. Biotechnol. 2017, 39, 174–180. [Google Scholar] [CrossRef]
- Lino, C.A.; Caldeira, J.C.; Peabody, D.S. Display of single-chain variable fragments on bacteriophage MS2 virus-like particles. J. Nanobiotechnol. 2017, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Tumban, E.; Peabody, J.; Peabody, D.S.; Chackerian, B. A Pan-HPV Vaccine Based on Bacteriophage PP7 VLPs Displaying Broadly Cross-Neutralizing Epitopes from the HPV Minor Capsid Protein, L2. PLoS ONE 2011, 6, e23310. [Google Scholar] [CrossRef]
- Janitzek, C.M.; Matondo, S.; Thrane, S.; Nielsen, M.; Kavishe, R.; Mwakalinga, S.B.; Theander, T.; Salanti, A.; Sander, A.F. Bacterial superglue generates a full-length circumsporozoite protein virus-like particle vaccine capable of inducing high and durable antibody responses. Malar. J. 2016, 15, 545. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, D.; Rolf, T.; Suckow, M.; Kranz, A.; Barbian, A.; Chan, J.A.; Leitsch, J.; Weniger, M.; Jenzelewski, V.; Kouskousis, B.; et al. Establishment of a yeast-based VLP platform for antigen presentation. Microb. Cell Factories 2018, 17, 17. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, H.-J. Yeast as an expression system for producing virus-like particles: What factors do we need to consider? Lett. Appl. Microbiol. 2017, 64, 111–123. [Google Scholar] [CrossRef]
- Huang, X.; Wang, X.; Zhang, J.; Xia, N.; Zhao, Q. Escherichia coli-derived virus-like particles in vaccine development. NPJ Vaccines 2017, 2, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.-Y.; Yeh, Y.-C.; Yang, Y.-C.; Chou, C.; Liu, M.-T.; Wu, H.-S.; Chan, J.-T.; Hsiao, P.-W. Mammalian Expression of Virus-Like Particles for Advanced Mimicry of Authentic Influenza Virus. PLoS ONE 2010, 5, e9784. [Google Scholar] [CrossRef] [Green Version]
- Arevalo, M.T.; Wong, T.M.; Ross, T.M. Expression and Purification of Virus-like Particles for Vaccination. J. Vis. Exp. 2016, 112, e54041. [Google Scholar] [CrossRef] [PubMed]
- Lindesmith, L.C.; McDaniel, J.R.; Changela, A.; Verardi, R.; Kerr, S.A.; Costantini, V.; Brewer-Jensen, P.D.; Mallory, M.L.; Voss, W.N.; Boutz, D.R.; et al. Sera Antibody Repertoire Analyses Reveal Mechanisms of Broad and Pandemic Strain Neutralizing Responses after Human Norovirus Vaccination. Immunity 2019, 50, 1530–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sainsbury, F.; Saxena, P.; Aljabali, A.A.; Saunders, K.; Evans, D.; Lomonossoff, G.P. Genetic Engineering and Characterization of Cowpea Mosaic Virus Empty Virus-Like Particles; Humana Press: Totowa, NJ, USA, 2014; pp. 139–153. [Google Scholar]
- Storni, F.; Zeltins, A.; Balke, I.; Heath, M.D.; Kramer, M.F.; Skinner, M.A.; Zha, L.; Roesti, E.; Engeroff, P.; Muri, L.; et al. Vaccine against peanut allergy based on engineered virus-like particles displaying single major peanut allergens. J. Allergy Clin. Immunol. 2019, 145, 1240–1253. [Google Scholar] [CrossRef]
- Larkin, E.J.; Brown, A.D.; Culver, J.N. Fabrication of Tobacco Mosaic Virus-Like Nanorods for Peptide Display. In Methods in Molecular Biology (Clifton, N.J.); Humana Press: Totowa, NJ, USA, 2018; Volume 1776, pp. 51–60. [Google Scholar]
- Abdoli, A.; Soleimanjahi, H.; Fotouhi, F.; Teimoori, A.; Pour Beiranvand, S.; Kianmehr, Z. Human Papillomavirus Type16- L1 VLP Production in Insect Cells. Iran. J. Basic Med. Sci. 2013, 16, 891–895. [Google Scholar]
- Jennings, G.T.; Bachmann, M.F. Immunodrugs: Therapeutic VLP-Based Vaccines for Chronic Diseases. Annu. Rev. Pharmacol. Toxicol. 2008, 49, 303–326. [Google Scholar] [CrossRef]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major findings and recent advances in virus–like particle (VLP)-based vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Bachmann, M.; Rohrer, U.; Kündig, T.; Burki, K.; Hengartner, H.; Zinkernagel, R. The influence of antigen organization on B cell responsiveness. Science 1993, 262, 1448–1451. [Google Scholar] [CrossRef]
- Phan, T.G.; Grigorova, I.; Okada, T.; Cyster, J.G. Subcapsular encounter and complement-dependent transport of immune complexes by lymph node B cells. Nat. Immunol. 2007, 8, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.A.; Bauer, M.; Manolova, V.; Muntwiler, S.; Saudan, P.; Bachmann, M.F. Cutting Edge: Limited Specialization of Dendritic Cell Subsets for MHC Class II-Associated Presentation of Viral Particles. J. Immunol. 2010, 184, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; El-Turabi, A.; Fettelschoss-Gabriel, A.; Vogel, M. The Prospects of an Active Vaccine Against Asthma Targeting IL-5. Front. Microbiol. 2018, 9, 2522. [Google Scholar] [CrossRef] [PubMed]
- Himmelstein, D.U.; Lawless, R.M.; Thorne, D.; Foohey, P.; Woolhandler, S. Medical Bankruptcy: Still Common Despite the Affordable Care Act. Am. J. Public Health 2019, 109, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Foerster, J.; Molęda, A. Feasibility Analysis of Interleukin-13 as a Target for a Therapeutic Vaccine. Vaccines (Basel) 2019, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA Application Review Guselkumab. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2017/761061Orig1s000MultidisciplineR.pdf. (accessed on 1 March 2020).
- Blauvelt, A.; Papp, K.; Griffiths, C.E.M.; Randazzo, B.; Wasfi, Y.; Shen, Y.-K.; Li, S.; Kimball, A. Efficacy and safety of guselkumab, an anti-interleukin-23 monoclonal antibody, compared with adalimumab for the continuous treatment of patients with moderate to severe psoriasis: Results from the phase III, double-blinded, placebo- and active comparator–controlled VOYAGE 1 trial. J. Am. Acad. Dermatol. 2017, 76, 405–417. [Google Scholar]
- Singh, S.; Kroe-Barrett, R.R.; Canada, K.A.; Zhu, X.; Sepulveda, E.; Wu, H.; He, Y.; Raymond, E.L.; Ahlberg, J.; Frego, L.E.; et al. Selective targeting of the IL23 pathway: Generation and characterization of a novel high-affinity humanized anti-IL23A antibody. MAbs 2015, 7, 778–791. [Google Scholar] [CrossRef]
- Gordon, K.B.; Strober, B.; Lebwohl, M.; Augustin, M.; Blauvelt, A.; Poulin, Y.; Papp, K.; Sofen, H.; Puig, L.; Foley, P.; et al. Efficacy and safety of risankizumab in moderate-to-severe plaque psoriasis (UltIMMa-1 and UltIMMa-2): Results from two double-blind, randomised, placebo-controlled and ustekinumab-controlled phase 3 trials. Lancet 2018, 392, 650–661. [Google Scholar] [CrossRef]
- Reich, K.; Papp, K.; Blauvelt, A.; Tyring, S.K.; Sinclair, R.; Thaçi, D.; Nograles, K.; Mehta, A.; Cichanowitz, N.; Li, Q.; et al. Tildrakizumab versus placebo or etanercept for chronic plaque psoriasis (reSURFACE 1 and reSURFACE 2): Results from two randomised controlled, phase 3 trials. Lancet 2017, 390, 276–288. [Google Scholar] [CrossRef]
- Bloch, Y.; Bouchareychas, L.; Merceron, R.; Składanowska, K.; Van den Bossche, L.; Detry, S.; Govindarajan, S.; Elewaut, D.; Haerynck, F.; Dullaers, M.; et al. Structural Activation of Pro-inflammatory Human Cytokine IL-23 by Cognate IL-23 Receptor Enables Recruitment of the Shared Receptor IL-12Rbeta1. Immunity 2018, 48, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Kasaian, M.T.; Raible, D.; Marquette, K.; Cook, T.A.; Zhou, S.; Tan, X.-Y.; Tchistiakova, L. IL-13 Antibodies Influence IL-13 Clearance in Humans by Modulating Scavenger Activity of IL-13Ralpha2. J. Immunol. 2011, 187, 561–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, S.; Chen, R.; Wei, J.; Guan, G.; Zhou, M.; Dong, N.; Cao, Y. Meta-analysis of randomized controlled trials for the efficacy and safety of anti-interleukin-13 therapy with lebrikizumab in patients with uncontrolled asthma. Allergy Asthma Proc. 2018, 39, 332–337. [Google Scholar] [CrossRef]
- Hanania, N.A.; Korenblat, P.; Chapman, K.; Bateman, E.D.; Kopecky, P.; Paggiaro, P.; Yokoyama, A.; Olsson, J.; Gray, S.; Holweg, C.T.J.; et al. Efficacy and safety of lebrikizumab in patients with uncontrolled asthma (LAVOLTA I and LAVOLTA II): Replicate, phase 3, randomised, double-blind, placebo-controlled trials. Lancet Respir. Med. 2016, 4, 781–796. [Google Scholar] [CrossRef]
- Tiwari, A.; Kasaian, M.; Heatherington, A.C.; Jones, H.M.; Hua, F. A mechanistic PK/PD model for two anti-IL13 antibodies explains the difference in total IL-13 accumulation observed in clinical studies. MAbs 2016, 8, 983–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripp, C.S.; Cuff, C.; Campbell, A.L.; Hendrickson, B.A.; Voss, J.; Melim, T.; Wu, C.; Cherniack, A.D.; Kim, K. RPC4046, A Novel Anti-interleukin-13 Antibody, Blocks IL-13 Binding to IL-13 alpha1 and alpha2 Receptors: A Randomized, Double-Blind, Placebo-Controlled, Dose-Escalation First-in-Human Study. Adv. Ther. 2017, 34, 1364–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, I.; Collins, M.H.; Dayan, Y.A.; Evans, L.; Gupta, S.; Schoepfer, A.M.; Straumann, A.; Safroneeva, E.; Grimm, M.; Smith, H.; et al. RPC4046, a Monoclonal Antibody Against IL13, Reduces Histologic and Endoscopic Activity in Patients With Eosinophilic Esophagitis. Gastroenterology 2019, 156, 592–603. [Google Scholar] [CrossRef] [Green Version]
- Popovic, B.; Breed, J.; Rees, D.; Gardener, M.; Vinall, L.; Kemp, B.; Spooner, J.; Keen, J.; Minter, R.; Uddin, F.; et al. Structural Characterisation Reveals Mechanism of IL-13-Neutralising Monoclonal Antibody Tralokinumab as Inhibition of Binding to IL-13Ralpha1 and IL-13Ralpha2. J. Mol. Biol. 2017, 429, 208–219. [Google Scholar] [CrossRef]
- Hart, T.K.; Blackburn, M.N.; Brigham-Burke, M.; DeDe, K.; Al-Mahdi, N.; Zia-Amirhosseini, P.; Cook, R.M. Preclinical efficacy and safety of pascolizumab (SB 240683): A humanized anti-interleukin-4 antibody with therapeutic potential in asthma. Clin. Exp. Immunol. 2002, 130, 93–100. [Google Scholar] [CrossRef]
- Kim, J.-E.; Jung, K.; Kim, J.-A.; Kim, S.-H.; Park, H.-S.; Kim, Y.S. Engineering of anti-human interleukin-4 receptor alpha antibodies with potent antagonistic activity. Sci. Rep. 2019, 9, 7772. [Google Scholar] [CrossRef] [Green Version]
- Getz, E.B.; Fisher, D.M.; Fuller, R. Human Pharmacokinetics/Pharmacodynamics of an Interleukin-4 and Interleukin-13 Dual Antagonist in Asthma. J. Clin. Pharmacol. 2009, 49, 1025–1036. [Google Scholar] [CrossRef]
- Maliszewski, C.R.; Sato, T.A.; Bos, T.V.; Waugh, S.; Dower, S.K.; Slack, J.; Beckmann, M.P.; Grabstein, K.H. Cytokine receptors and B cell functions. I. Recombinant soluble receptors specifically inhibit IL-1- and IL-4-induced B cell activities in vitro. J. Immunol. 1990, 144, 3028–3033. [Google Scholar] [PubMed]
- Borish, L.C.; Nelson, H.S.; Lanz, M.J.; Claussen, L.; Whitmore, J.B.; Agosti, J.M.; Garrison, L. Interleukin-4 receptor in moderate atopic asthma. A phase I/II randomized, placebo-controlled trial. Am. J. Respir. Crit. Care Med. 1999, 160, 1816–1823. [Google Scholar] [CrossRef] [PubMed]
- Corren, J.; Busse, W.; Meltzer, E.O.; Mansfield, L.; Bensch, G.; Fahrenholz, J.; Wenzel, S.E.; Chon, Y.; Dunn, M.; Weng, H.H.; et al. A randomized, controlled, phase 2 study of AMG 317, an IL-4Ralpha antagonist, in patients with asthma. Am. J. Respir. Crit. Care Med. 2010, 181, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Kraich, M.; Klein, M.; Patiño, E.; Harrer, H.; Nickel, J.; Sebald, W.; Mueller, T.D. A modular interface of IL-4 allows for scalable affinity without affecting specificity for the IL-4 receptor. BMC Biol. 2006, 4, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledger, K.S.; Agee, S.J.; Kasaian, M.T.; Forlow, S.B.; Durn, B.L.; Minyard, J.; Lu, Q.A.; Todd, J.; Vesterqvist, O.; Burczynski, M.E. Analytical validation of a highly sensitive microparticle-based immunoassay for the quantitation of IL-13 in human serum using the Erenna® immunoassay system. J. Immunol. Methods 2009, 350, 161–170. [Google Scholar] [CrossRef]
- Hua, F.; Ribbing, J.; Reinisch, W.; Cataldi, F.; Martin, S. A pharmacokinetic comparison of anrukinzumab, an anti- IL-13 monoclonal antibody, among healthy volunteers, asthma and ulcerative colitis patients. Br. J. Clin. Pharmacol. 2015, 80, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Horns, F.; Vollmers, C.; Dekker, C.L.; Quake, S.R. Signatures of selection in the human antibody repertoire: Selective sweeps, competing subclones, and neutral drift. Proc. Natl. Acad. Sci. USA 2019, 116, 1261–1266. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Boutz, D.R.; Chromikova, V.; Joyce, M.G.; Vollmers, C.; Leung, K.; Horton, A.P.; DeKosky, B.J.; Lee, C.-H.; Lavinder, J.J.; et al. Molecular-level analysis of the serum antibody repertoire in young adults before and after seasonal influenza vaccination. Nat. Med. 2016, 22, 1456–1464. [Google Scholar] [CrossRef]
- Kaur, K.; Zheng, N.-Y.; Smith, K.; Huang, M.; Li, L.; Pauli, N.T.; Dunand, C.J.H.; Lee, J.-H.; Morrissey, M.; Wu, Y.; et al. High Affinity Antibodies against Influenza Characterize the Plasmablast Response in SLE Patients After Vaccination. PLoS ONE 2015, 10, e0125618. [Google Scholar] [CrossRef]
- Wang, Y.; Sundling, C.; Wilson, R.; O’Dell, S.; Chen, Y.; Dai, K.; PhaD, G.E.; Zhu, J.; Xiao, Y.; Mascola, J.R.; et al. High-Resolution Longitudinal Study of HIV-1 Env Vaccine-Elicited B Cell Responses to the Virus Primary Receptor Binding Site Reveals Affinity Maturation and Clonal Persistence. J. Immunol. 2016, 196, 3729–3743. [Google Scholar] [CrossRef] [Green Version]
- Cavelti-Weder, C.; Timper, K.; Seelig, E.; Keller, C.; Osranek, M.; Lässing, U.; Spohn, G.; Maurer, P.; Müller, P.; Jennings, G.T.; et al. Development of an Interleukin-1beta Vaccine in Patients with Type 2 Diabetes. Mol. Ther. 2016, 24, 1003–1012. [Google Scholar] [CrossRef] [Green Version]
- Tissot, A.C.; Maurer, P.; Nussberger, J.; Sabat, R.; Pfister, T.; Ignatenko, S.; Volk, H.-D.; Stocker, H.; Müller, P.; Jennings, G.T.; et al. Effect of immunisation against angiotensin II with CYT006-AngQb on ambulatory blood pressure: A double-blind, randomised, placebo-controlled phase IIa study. Lancet 2008, 371, 821–827. [Google Scholar] [CrossRef]
- Fettelschoss-Gabriel, A.; Fettelschoss, V.; Thoms, F.; Giese, C.; Daniel, M.; Olomski, F.; Kamarachev, J.; Birkmann, K.; Bühler, M.; Kummer, M.; et al. Treating insect-bite hypersensitivity in horses with active vaccination against IL-5. J. Allergy Clin. Immunol. 2018, 142, 1194–1205. [Google Scholar] [CrossRef] [Green Version]
- Zeltins, A.; West, J.; Zabel, F.; El-Turabi, A.; Balke, I.; Haas, S.; Maudrich, M.; Storni, F.; Engeroff, P.; Jennings, G.T.; et al. Incorporation of tetanus-epitope into virus-like particles achieves vaccine responses even in older recipients in models of psoriasis, Alzheimer’s and cat allergy. NPJ Vaccines 2017, 2, 30. [Google Scholar] [CrossRef] [Green Version]
- Berry, C.M. Antibody immunoprophylaxis and immunotherapy for influenza virus infection: Utilization of monoclonal or polyclonal antibodies? Hum. Vaccin. Immunother. 2017, 14, 796–799. [Google Scholar] [CrossRef]
- Wang, X.; Mathieu, M.; Brezski, R. IgG Fc engineering to modulate antibody effector functions. Protein Cell 2017, 9, 63–73. [Google Scholar] [CrossRef]
- Sahoo, A.; Wali, S.; Nurieva, R. T helper 2 and T follicular helper cells: Regulation and function of interleukin-4. Cytokine Growth Factor Rev. 2016, 30, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Gowthaman, U.; Chen, J.S.; Zhang, B.; Flynn, W.F.; Lu, Y.; Song, W.; Joseph, J.; Gertie, J.A.; Xu, L.; Collet, M.A.; et al. Identification of a T follicular helper cell subset that drives anaphylactic IgE. Science 2019, 365, 6456. [Google Scholar] [CrossRef]
- Gowthaman, U.; Chen, J.S.; Eisenbarth, S.C. Regulation of IgE by T follicular helper cells. J. Leukoc. Biol. 2020, 107, 409–418. [Google Scholar] [CrossRef]
- Heesters, B.A.; Van Der Poel, C.E.; Das, A.; Carroll, M.C. Antigen Presentation to B Cells. Trends Immunol. 2016, 37, 844–854. [Google Scholar] [CrossRef]
- Laudenbach, M.; Baruffaldi, F.; Robinson, C.; Carter, P.; Seelig, D.; Baehr, C.; Pravetoni, M. Blocking interleukin-4 enhances efficacy of vaccines for treatment of opioid abuse and prevention of opioid overdose. Sci. Rep. 2018, 8, 5508. [Google Scholar] [CrossRef] [PubMed]
- Blauvelt, A.; Simpson, E.L.; Tyring, S.K.; Purcell, L.A.; Shumel, B.; Petro, C.D.; Akinlade, B.; Gadkari, A.; Eckert, L.; Graham, N.M.; et al. Dupilumab does not affect correlates of vaccine-induced immunity: A randomized, placebo-controlled trial in adults with moderate-to-severe atopic dermatitis. J. Am. Acad. Dermatol. 2019, 80, 158–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, C.E.; Georgakopoulos, J.R.; Ladda, M.; Ighani, A.; Mufti, A.; Drucker, A.M.; Piguet, V.; Yeung, J. Evaluation of long-term efficacy, safety, and reasons for discontinuation of dupilumab for moderate-to-severe atopic dermatitis in clinical practice: A retrospective cohort study. J. Am. Acad. Dermatol. 2020, 82. In Press. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Liu, D.; Liu, C.-T. The Efficacy and Safety of Antiinterleukin 13, a Monoclonal Antibody, in Adult Patients With Asthma: A Systematic Review and Meta-Analysis. Medicine (Baltimore) 2016, 95, e2556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cheng, J.; Li, Y.; He, R.; Pan, P.; Su, X.; Hu, C. The Safety and Efficacy of Anti–IL-13 Treatment with Tralokinumab (CAT-354) in Moderate to Severe Asthma: A Systematic Review and Meta-Analysis. J. Allergy Clin. Immunol. Pr. 2019, 7, 2661–2671. [Google Scholar] [CrossRef]
- Carlsson, M.; Braddock, M.; Li, Y.; Wang, J.; Xu, W.; White, N.; Megally, A.; Hunter, G.; Colice, G. Evaluation of Antibody Properties and Clinically Relevant Immunogenicity, Anaphylaxis, and Hypersensitivity Reactions in Two Phase III Trials of Tralokinumab in Severe, Uncontrolled Asthma. Drug Saf. 2019, 42, 769–784. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Target MAb Affinity | Clinical Efficacy 1 | Fold-Increase Vs. Receptor Binding 2 | Ref | |
|---|---|---|---|---|
| Guselkumab | 2 pM | 91% | 3000-fold | [28,29] |
| Risankizumab | 2 pM | 91% | 3000-fold | [30,31] |
| Tildrakizumab | 300 pM | 61% | 20-fold | [32] |
| Mab/Blocker | Net Target 1 | Affinity (pM) | Tested in 2 | Effective in | Fold Affinity Vs. Receptor 3 | Ref |
|---|---|---|---|---|---|---|
| Group 1: IL-13-only blockers | ||||||
| Lebrikizumab | IL-13Ra1 | <10 | Asthma, AD | AD, Asthma 4 | 3.5 | [35,36] |
| Anrukinzumab | IL-13Ra1 | 385 | UC, Asthma | --- | 0.1 | [37] |
| RPC4046 | IL-13Ra1/Ra2 | 50 | EoE | EoE | 0.7 | [38,39] |
| Tralokinumab | IL-13Ra1/Ra2 | 58 | Asthma, AD | AD | 0.8 | [40] |
| Group 2: IL-4-selective and IL4/IL-13 combined blockers | ||||||
| Pascolizumab | IL-4Ra | 60 | Asthma | --- | 1.4 | [41] |
| Dupilumab | IL-4Ra, IL-13Ra1 | 9 | Asthma, AD | Asthma, AD | 10 | [42] |
| Pitrankinra | IL-4Ra, IL-13Ra1 | 100 5 | Asthma | --- | 1 | [43] |
| Altrakincept | IL-4Ra, IL-13Ra1 | 1000 | Asthma | --- | 0.1 | [44,45] |
| AMG317 | IL-4Ra, IL-13Ra1 | 180 | Asthma | --- | 0.8 | [46] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foerster, J.; Molęda, A. Virus-Like Particle-Mediated Vaccination against Interleukin-13 May Harbour General Anti-Allergic Potential beyond Atopic Dermatitis. Viruses 2020, 12, 438. https://doi.org/10.3390/v12040438
Foerster J, Molęda A. Virus-Like Particle-Mediated Vaccination against Interleukin-13 May Harbour General Anti-Allergic Potential beyond Atopic Dermatitis. Viruses. 2020; 12(4):438. https://doi.org/10.3390/v12040438
Chicago/Turabian StyleFoerster, John, and Aleksandra Molęda. 2020. "Virus-Like Particle-Mediated Vaccination against Interleukin-13 May Harbour General Anti-Allergic Potential beyond Atopic Dermatitis" Viruses 12, no. 4: 438. https://doi.org/10.3390/v12040438
APA StyleFoerster, J., & Molęda, A. (2020). Virus-Like Particle-Mediated Vaccination against Interleukin-13 May Harbour General Anti-Allergic Potential beyond Atopic Dermatitis. Viruses, 12(4), 438. https://doi.org/10.3390/v12040438