Battling Neurodegenerative Diseases with Adeno-Associated Virus-Based Approaches


 ,
,
Abstract
:1. Background
2. Viral Vector-Based Gene Therapy: Efficiency and Selectivity of Delivery
2.1. Choosing an Ideal Vector
2.2. Advantages of AAV-Based Vectors
3. AAV in NDD Experimental Therapy
3.1. In Vitro vs. In Vivo Models
3.2. Selection of Optimal Gene Therapy Strategy
3.3. AAV-Based Gene Therapy for Parkinson’s Disease
3.4. AAV Vectors and Gene Targeting
4. Modulation of Interaction Between the CNS and the Environment
5. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AAV | Adeno-associated virus |
| Ad | Adenovirus |
| BBB | Blood–brain barrier |
| CNS | Central neural system |
| DNA | Deoxyribonucleic acid |
| GDNF | Glial cell-line derived neurotrophic factor |
| FGF | Fibroblast growth factor |
| HSPG | Heparan sulfate proteoglycan |
| HSV | Herpes simplex virus |
| NDD | Neuro-degenerative disease |
| PDGFR | Platelet-derived growth factor receptor |
| RNA | Ribonucleic acid |
| SFV | Semliki forest virus |
| SMN | Survival motor neuron protein |
| VEGF | Vascular endothelial growth factor |
References
- Bordi, M.; Berg, M.J.; Mohan, P.S.; Peterhoff, C.M.; Alldred, M.J.; Che, S.; Ginsberg, S.D.; Nixon, R.A. Autophagy flux in CA1 neurons of Alzheimer hippocampus: Increased induction overburdens failing lysosomes to propel neuritic dystrophy. Autophagy 2016, 12, 2467–2483. [Google Scholar] [CrossRef]
- Ingelsson, M.; Fukumoto, H.; Newell, K.L.; Growdon, J.H.; Hedley-Whyte, E.T.; Frosch, M.P.; Albert, M.S.; Hyman, B.T.; Irizarry, M.C. Early Abeta accumulation and progressive synaptic loss, gliosis, and tangle formation in AD brain. Neurology 2004, 62, 925–931. [Google Scholar] [CrossRef]
- Monahan, J.A.; Warren, M.; Carvey, P.M. Neuroinflammation and peripheral immune infiltration in Parkinson’s disease: An autoimmune hypothesis. Cell Transplant. 2008, 17, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Brochard, V.; Combadière, B.; Prigent, A.; Laouar, Y.; Perrin, A.; Beray-Berthat, V.; Bonduelle, O.; Alvarez-Fischer, D.; Callebert, J.; Launay, J.-M.; et al. Infiltration of CD4+ lymphocytes into the brain contributes to neurodegeneration in a mouse model of Parkinson disease. J. Clin. Investig. 2008, 119, 182–192. [Google Scholar] [CrossRef]
- Rousseaux, M.W.; De Haro, M.; Lasagna-Reeves, C.A.; De Maio, A.; Park, J.; Jafar-Nejad, P.; Al-Ramahi, I.; Sharma, A.; See, L.; Lu, N.; et al. TRIM28 regulates the nuclear accumulation and toxicity of both alpha-synuclein and tau. eLife 2016, 5, 18748. [Google Scholar] [CrossRef]
- Duggan, M.; Torkzaban, B.; Ahooyi, T.M.; Khalili, K.; Gordon, J. Age-related neurodegenerative diseases. J. Cell. Physiol. 2019, 235, 3131–3141. [Google Scholar] [CrossRef]
- Min, B.; Chung, K.C. New insight into transglutaminase 2 and link to neurodegenerative diseases. BMB Rep. 2018, 51, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Flavin, W.P.; Bousset, L.; Green, Z.C.; Chu, Y.; Skarpathiotis, S.; Chaney, M.J.; Kordower, J.H.; Melki, R.; Campbell, E.M. Endocytic vesicle rupture is a conserved mechanism of cellular invasion by amyloid proteins. Acta Neuropathol. 2017, 134, 629–653. [Google Scholar] [CrossRef]
- Gorantla, N.V.; Chinnathambi, S. Tau Protein Squired by Molecular Chaperones During Alzheimer’s Disease. J. Mol. Neurosci. 2018, 66, 356–368. [Google Scholar] [CrossRef]
- Cristofani, R.; Crippa, V.; Rusmini, P.; Cicardi, M.E.; Meroni, M.; Licata, N.V.; Sala, G.; Giorgetti, E.; Grunseich, C.; Galbiati, M.; et al. Inhibition of retrograde transport modulates misfolded protein accumulation and clearance in motoneuron diseases. Autophagy 2017, 13, 1280–1303. [Google Scholar] [CrossRef]
- Soares, T.; Reis, S.; Pinho, B.; Duchen, M.R.; Oliveira, J.M. Targeting the proteostasis network in Huntington’s disease. Ageing Res. Rev. 2019, 49, 92–103. [Google Scholar] [CrossRef]
- Lim, J.; Bang, Y.; Choi, H.J. Abnormal hippocampal neurogenesis in Parkinson’s disease: Relevance to a new therapeutic target for depression with Parkinson’s disease. Arch. Pharmacal Res. 2018, 41, 943–954. [Google Scholar] [CrossRef]
- Shah, S.Z.A.; Zhao, D.; Hussain, T.; Sabir, N.; Mangi, M.H.; Yang, L. p62-Keap1-NRF2-ARE Pathway: A Contentious Player for Selective Targeting of Autophagy, Oxidative Stress and Mitochondrial Dysfunction in Prion Diseases. Front. Mol. Neurosci. 2018, 11, 310. [Google Scholar] [CrossRef] [Green Version]
- Ariosa, A.R.; Klionsky, D.J. Autophagy core machinery: Overcoming spatial barriers in neurons. J. Mol. Med. 2016, 94, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Dhawan, A.; Kadam, A.; Shinde, A. Autophagy and Mitochondria: Targets in Neurodegenerative Disorders. CNS Neurol. Disord. Drug Targets 2018, 17, 696–705. [Google Scholar] [CrossRef]
- Tramutola, A.; Lanzillotta, C.; Di Domenico, F. Targeting mTOR to reduce Alzheimer-related cognitive decline: From current hits to future therapies. Expert Rev. Neurother. 2016, 17, 33–45. [Google Scholar] [CrossRef]
- Lee, C.-C.; Chang, C.-P.; Lin, C.-J.; Lai, H.-L.; Kao, Y.-H.; Cheng, S.-J.; Chen, H.-M.; Liao, Y.-P.; Faivre, E.; Buée, L.; et al. Adenosine Augmentation Evoked by an ENT1 Inhibitor Improves Memory Impairment and Neuronal Plasticity in the APP/PS1 Mouse Model of Alzheimer’s Disease. Mol. Neurobiol. 2018, 55, 8936–8952. [Google Scholar] [CrossRef]
- Tramutola, A.; Lanzillotta, C.; Barone, E.; Arena, A.; Zuliani, I.; Mosca, L.; Blarzino, C.; Butterfield, D.A.; Perluigi, M.; Di Domenico, F. Intranasal rapamycin ameliorates Alzheimer-like cognitive decline in a mouse model of Down syndrome. Transl. Neurodegener. 2018, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Musial, T.; Molina-Campos, E.; Bean, L.A.; Ybarra, N.; Borenstein, R.; Russo, M.; Buss, E.; Justus, D.; Neuman, K.M.; Ayala, G.D.; et al. Store depletion-induced h-channel plasticity rescues a channelopathy linked to Alzheimer’s disease. Neurobiol. Learn. Mem. 2018, 154, 141–157. [Google Scholar] [CrossRef]
- Guo, N.; Xie, W.; Xiong, P.; Li, H.; Wang, S.; Chen, G.; Gao, Y.; Zhou, J.; Zhang, Y.; Bu, G.; et al. Cyclin-dependent kinase 5-mediated phosphorylation of chloride intracellular channel 4 promotes oxidative stress-induced neuronal death. Cell Death Dis. 2018, 9, 951. [Google Scholar] [CrossRef]
- Picconi, B.; De Leonibus, E.; Calabresi, P. Synaptic plasticity and levodopa-induced dyskinesia: Electrophysiological and structural abnormalities. J. Neural Transm. 2018, 125, 1263–1271. [Google Scholar] [CrossRef]
- Cacabelos, R. Parkinson’s Disease: From Pathogenesis to Pharmacogenomics. Int. J. Mol. Sci. 2017, 18, 551. [Google Scholar] [CrossRef]
- Burchfield, J.S.; Li, Q.; Wang, H.Y.; Wang, R.-F. JMJD3 as an epigenetic regulator in development and disease. Int. J. Biochem. Cell Boil. 2015, 67, 148–157. [Google Scholar] [CrossRef]
- Li, P.; Marshall, L.; Oh, G.; Jakubowski, J.L.; Groot, D.; He, Y.; Wang, T.; Petronis, A.; Labrie, V. Epigenetic dysregulation of enhancers in neurons is associated with Alzheimer’s disease pathology and cognitive symptoms. Nat. Commun. 2019, 10, 2246. [Google Scholar] [CrossRef] [Green Version]
- Vucic, S.; Ryder, J.; Mekhael, L.; Rd, H.; Mathers, S.; Needham, M.; Dw, S.; Mc, K. TEALS study group Phase 2 randomized placebo controlled double blind study to assess the efficacy and safety of tecfidera in patients with amyotrophic lateral sclerosis (TEALS Study). Medicine 2020, 99, e18904. [Google Scholar] [CrossRef]
- Petramfar, P.; Hajari, F.; Yousefi, G.; Azadi, S.; Hamedi, A. Efficacy of oral administration of licorice as an adjunct therapy on improving the symptoms of patients with Parkinson’s disease, A randomized double blinded clinical trial. J. Ethnopharmacol. 2020, 247, 112226. [Google Scholar] [CrossRef]
- Andrade, S.; Ramalho, M.J.; Loureiro, J.A.; Pereira, M.D.C. Natural Compounds for Alzheimer’s Disease Therapy: A Systematic Review of Preclinical and Clinical Studies. Int. J. Mol. Sci. 2019, 20, 2313. [Google Scholar] [CrossRef] [Green Version]
- Gasperi, V.; Sibilano, M.; Savini, I.; Catani, M.V. Niacin in the Central Nervous System: An Update of Biological Aspects and Clinical Applications. Int. J. Mol. Sci. 2019, 20, 974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farlow, M.R.; Thompson, R.E.; Wei, L.-J.; Tuchman, A.J.; Grenier, E.; Crockford, D.; Wilke, S.; Benison, J.; Alkon, D.L. A Randomized, Double-Blind, Placebo-Controlled, Phase II Study Assessing Safety, Tolerability, and Efficacy of Bryostatin in the Treatment of Moderately Severe to Severe Alzheimer’s Disease. J. Alzheimer’s Dis. 2019, 67, 555–570. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.-X.; Ma, H.-H.; Ding, H.; Li, W.-W.; Zhu, M. Preliminary optimization of a Chinese herbal medicine formula based on the neuroprotective effects in a rat model of rotenone-induced Parkinson’s disease. J. Integr. Med. 2018, 16, 290–296. [Google Scholar] [CrossRef]
- Johnstone, D.M.; Moro, C.; Stone, J.; Benabid, A.-L.; Mitrofanis, J. Turning On Lights to Stop Neurodegeneration: The Potential of Near Infrared Light Therapy in Alzheimer’s and Parkinson’s Disease. Front. Mol. Neurosci. 2016, 9, 75001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeler, A.; Elmallah, M.; Flotte, T.R. Gene Therapy 2017: Progress and Future Directions. Clin. Transl. Sci. 2017, 10, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendell, J.R.; Al-Zaidy, S.; Shell, R.; Arnold, W.D.; Rodino-Klapac, L.R.; Prior, T.W.; Lowes, L.P.; Alfano, L.; Berry, K.; Church, K.; et al. Single-Dose Gene-Replacement Therapy for Spinal Muscular Atrophy. N. Engl. J. Med. 2017, 377, 1713–1722. [Google Scholar] [CrossRef]
- Massaro, G.; Mattar, C.N.Z.; Wong, A.M.S.; Sirka, E.; Buckley, S.M.K.; Herbert, B.R.; Karlsson, S.; Perocheau, D.P.; Burke, D.; Heales, S.; et al. Fetal gene therapy for neurodegenerative disease of infants. Nat. Med. 2018, 24, 1317–1323. [Google Scholar] [CrossRef]
- Holthaus, S.-M.K.; Herranz-Martin, S.; Massaro, G.; Aristorena, M.; Hoke, J.; Hughes, M.P.; Maswood, R.; Semenyuk, O.; Basche, M.; Shah, A.Z.; et al. Neonatal brain-directed gene therapy rescues a mouse model of neurodegenerative CLN6 Batten disease. Hum. Mol. Genet. 2019, 28, 3867–3879. [Google Scholar] [CrossRef]
- Das, S.; Huang, S.; Lo, A.W. Acceleration of rare disease therapeutic development: A case study of AGIL-AADC. Drug Discov. Today 2019, 24, 678–684. [Google Scholar] [CrossRef]
- Gessler, D.J.; Gao, G. Gene Therapy for the Treatment of Neurological Disorders: Metabolic Disorders. Breast Cancer 2016, 1382, 429–465. [Google Scholar]
- Conlon, T.J.; Mavilio, F. The Pharmacology of Gene and Cell Therapy. Mol. Ther. Methods Clin. Dev. 2018, 8, 181–182. [Google Scholar] [CrossRef] [Green Version]
- Cwetsch, A.W.; Pinto, B.; Savardi, A.; Cancedda, L. In vivo methods for acute modulation of gene expression in the central nervous system. Prog. Neurobiol. 2018, 168, 69–85. [Google Scholar] [CrossRef]
- Mavilio, F.; Ferrari, G. Genetic modification of somatic stem cells. The progress, problems and prospects of a new therapeutic technology. EMBO Rep. 2008, 9 (Suppl. 1), S64–S69. [Google Scholar] [CrossRef] [Green Version]
- Vagner, T.; Dvorzhak, A.; Wójtowicz, A.M.; Harms, C.; Grantyn, R. Systemic application of AAV vectors targeting GFAP-expressing astrocytes in Z -Q175-KI Huntington’s disease mice. Mol. Cell. Neurosci. 2016, 77, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Shahaduzzaman, M.; Nash, K.; Hudson, C.; Sharif, M.; Grimmig, B.; Lin, X.; Bai, G.; Liu, H.; Ugen, K.E.; Cao, C.; et al. Anti-Human α-Synuclein N-Terminal Peptide Antibody Protects against Dopaminergic Cell Death and Ameliorates Behavioral Deficits in an AAV-α-Synuclein Rat Model of Parkinson’s Disease. PLoS ONE 2015, 10, e0116841. [Google Scholar] [CrossRef] [PubMed]
- Chapdelaine, P.; Gerard, C.; Sanchez, N.; Cherif, K.; Rousseau, J.; Ouellet, D.L.; Jauvin, D.; Tremblay, J.P. Development of an AAV9 coding for a 3XFLAG-TALEfrat#8-VP64 able to increase in vivo the human frataxin in YG8R mice. Gene Ther. 2016, 23, 606–614. [Google Scholar] [PubMed] [Green Version]
- Morabito, G.; Giannelli, S.G.; Ordazzo, G.; Bido, S.; Castoldi, V.; Indrigo, M.; Cabassi, T.; Cattaneo, S.; Luoni, M.; Cancellieri, C.; et al. AAV-PHP.B-Mediated Global-Scale Expression in the Mouse Nervous System Enables GBA1 Gene Therapy for Wide Protection from Synucleinopathy. Mol. Ther. 2017, 25, 2727–2742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foust, K.D.; Nurre, E.; Montgomery, C.L.; Hernandez, A.; Chan, C.M.; Kaspar, B.K. Intravascular AAV9 preferentially targets neonatal neurons and adult astrocytes. Nat. Biotechnol. 2008, 27, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Sands, S.A.; Yue, Y.; Zhang, K.; Levine, S.M.; Duan, N. An Engineered Galactosylceramidase Construct Improves AAV Gene Therapy for Krabbe Disease in Twitcher Mice. Hum. Gene Ther. 2019, 30, 1039–1051. [Google Scholar] [CrossRef]
- Perez, B.A.; Shutterly, A.; Chan, Y.; Byrne, B.; Corti, M. Management of Neuroinflammatory Responses to AAV-Mediated Gene Therapies for Neurodegenerative Diseases. Brain Sci. 2020, 10, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraiva, J.; Nobre, R.J.; De Almeida, L.P. Gene therapy for the CNS using AAVs: The impact of systemic delivery by AAV9. J. Control. Release 2016, 241, 94–109. [Google Scholar] [CrossRef]
- Hudry, E.; Andres-Mateos, E.; Lerner, E.P.; Volak, A.; Cohen, O.; Hyman, B.T.; Maguire, C.A.; Vandenberghe, L.H. Efficient Gene Transfer to the Central Nervous System by Single-Stranded Anc80L65. Mol. Ther. Methods Clin. Dev. 2018, 10, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Hösel, M.; Huber, A.; Bohlen, S.; Lucifora, J.; Ronzitti, G.; Puzzo, F.; Boisgerault, F.; Hacker, U.T.; Kwanten, W.; Klöting, N.; et al. Autophagy determines efficiency of liver-directed gene therapy with adeno-associated viral vectors. Hepatology 2017, 66, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Richter, M.; Psatha, N.; Li, C.; Kim, J.; Liu, J.; Ehrhardt, A.; Nilsson, S.K.; Cao, B.; Palmer, N.; et al. A Combined In Vivo HSC Transduction/Selection Approach Results in Efficient and Stable Gene Expression in Peripheral Blood Cells in Mice. Mol. Ther. Methods Clin. Dev. 2017, 8, 52–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langou, K.; Moumen, A.; Pellegrino, C.; Aebischer, J.; Medina, I.; Aebischer, P.; Raoul, C. AAV-mediated expression of wild-type and ALS-linked mutant VAPB selectively triggers death of motoneurons through a Ca2+-dependent ER-associated pathway. J. Neurochem. 2010, 114, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Zhang, T.; Kuerban, B.; Jin, Y.L.; Le, W.; Hara, H.; Fan, D.S.; Wang, Y.J.; Tabira, T.; Chui, D.H. Autophagy is involved in oral rAAV/Abeta vaccine-induced Abeta clearance in APP/PS1 transgenic mice. Neurosci. Bull. 2015, 31, 491–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, H.; Shen, L.; Li, T.; Ren, C.; Ding, S.; Wang, L.; Zhang, Z.; Liu, X.; Zhang, Q.; Geng, D.; et al. Alpha-synuclein overexpression in the olfactory bulb initiates prodromal symptoms and pathology of Parkinson’s disease. Transl. Neurodegener. 2018, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Zheng, X.; Zhang, L.; Luo, H.; Qian, L.; Fu, X.; Liu, Y.; Gao, Y.; Niu, M.; Meng, J.; et al. The Neuron-Specific Protein TMEM59L Mediates Oxidative Stress-Induced Cell Death. Mol. Neurobiol. 2016, 54, 4189–4200. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, R.; Martínez-Muriana, A.; Leiva, T.; Gregorio, D.; Ariza, L.; Morell, M.; Esteban-Pérez, J.; García-Redondo, A.; Calvo, A.; Atencia-Cibreiro, G.; et al. Neuregulin-1 promotes functional improvement by enhancing collateral sprouting in SOD1G93A ALS mice and after partial muscle denervation. Neurobiol. Dis. 2016, 95, 168–178. [Google Scholar]
- Kato, S.; Sugawara, M.; Kobayashi, K.; Kimura, K.; Inoue, K.-I.; Takada, M.; Kobayashi, K. Enhancement of the transduction efficiency of a lentiviral vector for neuron-specific retrograde gene delivery through the point mutation of fusion glycoprotein type E. J. Neurosci. Methods 2019, 311, 147–155. [Google Scholar] [CrossRef]
- Artusi, S.; Miyagawa, Y.; Goins, W.F.; Cohen, J.; Glorioso, J.C. Herpes Simplex Virus Vectors for Gene Transfer to the Central Nervous System. Disease 2018, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- Ansorena, E.; Casales, E.; Aranda, A.; Tamayo, E.; Garbayo, E.; Smerdou, C.; Blanco-Prieto, M.J.; Aymerich, M.S. A simple and efficient method for the production of human glycosylated glial cell line-derived neurotrophic factor using a Semliki Forest virus expression system. Int. J. Pharm. 2013, 440, 19–26. [Google Scholar] [CrossRef]
- Berns, K.I.; Muzyczka, N. AAV: An Overview of Unanswered Questions. Hum. Gene Ther. 2017, 28, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Ehrengruber, M.U. Alphaviral Vectors for Gene Transfer into Neurons. Mol. Neurobiol. 2002, 26, 183–202. [Google Scholar] [CrossRef]
- Ehrengruber, M.U.; Lundstrom, K.; Schweitzer, C.; Heuss, C.; Schlesinger, S.; Gähwiler, B.H. Recombinant Semliki Forest virus and Sindbis virus efficiently infect neurons in hippocampal slice cultures. Proc. Natl. Acad. Sci. USA 1999, 96, 7041–7046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VandenDriessche, T.; Thorrez, L.; Acosta-Sanchez, A.; Petrus, I.; Wang, L.; Ma, L.; De Waele, L.; Iwasaki, Y.; Gillijns, V.; Wilson, J.M.; et al. Efficacy and safety of adeno-associated viral vectors based on serotype 8 and 9 vs. lentiviral vectors for hemophilia B gene therapy. J. Thromb. Haemost. 2006, 5, 16–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoy, S.M. Onasemnogene Abeparvovec: First Global Approval. Drugs 2019, 79, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.M.; Pestronk, A.; David, W.; Rothstein, J.; Simpson, E.; Appel, S.H.; Andres, P.L.; Mahoney, K.; Allred, P.; Alexander, K.; et al. An antisense oligonucleotide against SOD1 delivered intrathecally for patients with SOD1 familial amyotrophic lateral sclerosis: A phase 1, randomised, first-in-man study. Lancet Neurol. 2013, 12, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Auricchio, A.; Smith, A.; Ali, R.R. The Future Looks Brighter After 25 Years of Retinal Gene Therapy. Hum. Gene Ther. 2017, 28, 982–987. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Zhou, S.; Nakai, H.; Thomas, C.E.; Storm, T.A.; Fuess, S.; Matsushita, T.; Allen, J.; Surosky, R.; Lochrie, M.; et al. Preclinical in vivo evaluation of pseudotyped adeno-associated virus vectors for liver gene therapy. Blood 2003, 102, 2412–2419. [Google Scholar] [CrossRef]
- Paulk, N.; Pekrun, K.; Zhu, E.; Nygaard, S.; Li, B.; Xu, J.; Chu, K.; Leborgne, C.; Dane, A.P.; Haft, A.; et al. Bioengineered AAV Capsids with Combined High Human Liver Transduction In Vivo and Unique Humoral Seroreactivity. Mol. Ther. 2017, 26, 289–303. [Google Scholar] [CrossRef] [Green Version]
- White, S.; Nicklin, S.A.; Buüning, H.; Brosnan, M.J.; Leike, K.; Papadakis, E.D.; Hallek, M.; Baker, A.H. Targeted Gene Delivery to Vascular Tissue In Vivo by Tropism-Modified Adeno-Associated Virus Vectors. Circulation 2004, 109, 513–519. [Google Scholar] [CrossRef]
- Work, L.M.; Büning, H.; Pustulka-Hunt, E.; Nicklin, S.A.; Denby, L.; Britton, N.; Leike, K.; Odenthal, M.; Drebber, U.; Hallek, M.; et al. Vascular bed-targeted in vivo gene delivery using tropism-modified adeno-associated viruses. Mol. Ther. 2006, 13, 683–693. [Google Scholar] [CrossRef]
- Halbert, C.L.; Allen, J.M.; Miller, A.D. Adeno-Associated Virus Type 6 (AAV6) Vectors Mediate Efficient Transduction of Airway Epithelial Cells in Mouse Lungs Compared to That of AAV2 Vectors. J. Virol. 2001, 75, 6615–6624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, S.Q.; Hauser, M.A.; DelloRusso, C.; Duan, D.; Crawford, R.W.; Phelps, S.F.; Harper, H.A.; Robinson, A.S.; Engelhardt, J.F.; Brooks, S.V.; et al. Modular flexibility of dystrophin: Implications for gene therapy of Duchenne muscular dystrophy. Nat. Med. 2002, 8, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Tervo, D.G.R.; Hwang, B.-Y.; Viswanathan, S.; Gaj, T.; Lavzin, M.; Ritola, K.D.; Lindo, S.; Michael, S.; Kuleshova, E.; Ojala, D.; et al. A Designer AAV Variant Permits Efficient Retrograde Access to Projection Neurons. Neuron 2016, 92, 372–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asokan, A.; Schaffer, D.V.; Samulski, R.J. The AAV Vector Toolkit: Poised at the Clinical Crossroads. Mol. Ther. 2012, 20, 699–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, J.; Thalheimer, F.B.; Höpfner, F.; Kerzel, T.; Khodosevich, K.; García-González, D.; Monyer, H.; Diester, I.; Büning, H.; Carette, J.E.; et al. GluA4-Targeted AAV Vectors Deliver Genes Selectively to Interneurons while Relying on the AAV Receptor for Entry. Mol. Ther. Methods Clin. Dev. 2019, 14, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Qing, K.; Mah, C.; Hansen, J.; Zhou, S.; Dwarki, V.; Srivastava, A. Human fibroblast growth factor receptor 1 is a co-receptor for infection by adeno-associated virus 2. Nat. Med. 1999, 5, 71–77. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Davidson, B.L.; Stein, C.S.; Martins, I.; Scudiero, D.; Monks, A.; Chiorini, J.A. Identification of PDGFR as a receptor for AAV-5 transduction. Nat. Med. 2003, 9, 1306–1312. [Google Scholar] [CrossRef] [Green Version]
- Kashiwakura, Y.; Tamayose, K.; Iwabuchi, K.; Hirai, Y.; Shimada, T.; Matsumoto, K.; Nakamura, T.; Watanabe, M.; Oshimi, K.; Daida, H. Hepatocyte Growth Factor Receptor Is a Coreceptor for Adeno-Associated Virus Type 2 Infection. J. Virol. 2005, 79, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Asokan, A.; Hamra, J.B.; Govindasamy, L.; Agbandje-McKenna, M.; Samulski, R.J. Adeno-associated virus type 2 contains an integrin alpha5beta1 binding domain essential for viral cell entry. J. Virol. 2006, 80, 8961–8969. [Google Scholar] [CrossRef] [Green Version]
- Summerford, C.; Bartlett, J.S.; Samulski, R.J. AlphaVbeta5 integrin: A co-receptor for adeno-associated virus type 2 infection. Nat. Med. 1999, 5, 78–82. [Google Scholar] [CrossRef]
- Walters, R.W.; Pilewski, J.M.; Chiorini, J.A.; Zabner, J. Secreted and Transmembrane Mucins Inhibit Gene Transfer with AAV4 More Efficiently than AAV5. J. Boil. Chem. 2002, 277, 23709–23713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akache, B.; Grimm, D.; Pandey, K.; Yant, S.R.; Xu, H.; Kay, M.A. The 37/67-kilodalton laminin receptor is a receptor for adeno-associated virus serotypes 8, 2, 3, and 9. J. Virol. 2006, 80, 9831–9836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vana, K.; Zuber, C.; Pflanz, H.; Kolodziejczak, D.; Zemora, G.; Bergmann, A.-K.; Weiss, S. LRP/LR as an alternative promising target in therapy of prion diseases, Alzheimer’s disease and cancer. Infect. Disord. Drug Targets 2009, 9, 69–80. [Google Scholar] [CrossRef]
- Cearley, C.N.; Vandenberghe, L.H.; Parente, M.K.; Carnish, E.R.; Wilson, J.M.; Wolfe, J.H. Expanded Repertoire of AAV Vector Serotypes Mediate Unique Patterns of Transduction in Mouse Brain. Mol. Ther. 2008, 16, 1710–1718. [Google Scholar] [CrossRef] [PubMed]
- Hildinger, M.; Auricchio, A.; Gao, G.; Wang, L.; Chirmule, N.; Wilson, J.M. Hybrid Vectors Based on Adeno-Associated Virus Serotypes 2 and 5 for Muscle-Directed Gene Transfer. J. Virol. 2001, 75, 6199–6203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, G.-P.; Alvira, M.R.; Wang, L.; Calcedo, R.; Johnston, J.; Wilson, J.M. Novel adeno-associated viruses from rhesus monkeys as vectors for human gene therapy. Proc. Natl. Acad. Sci. USA 2002, 99, 11854–11859. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Asokan, A.; Wu, Z.; Van Dyke, T.; DiPrimio, N.; Johnson, J.; Govindaswamy, L.; Agbandje-McKenna, M.; Leichtle, S.; Redmond, D.; et al. Engineering and Selection of Shuffled AAV Genomes: A New Strategy for Producing Targeted Biological Nanoparticles. Mol. Ther. 2008, 16, 1252–1260. [Google Scholar] [CrossRef]
- Maheshri, N.; Koerber, J.T.; Kaspar, B.K.; Schaffer, D.V. Directed evolution of adeno-associated virus yields enhanced gene delivery vectors. Nat. Biotechnol. 2006, 24, 198–204. [Google Scholar] [CrossRef]
- Körbelin, J.; Sieber, T.; Michelfelder, S.; Lunding, L.; Spies, E.; Hunger, A.; Alawi, M.; Rapti, K.; Indenbirken, D.; Müller, O.J.; et al. Pulmonary Targeting of Adeno-associated Viral Vectors by Next-generation Sequencing-guided Screening of Random Capsid Displayed Peptide Libraries. Mol. Ther. 2016, 24, 1050–1061. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.H.; Chang, M.; Davidson, B.L. Molecular signatures of disease brain endothelia provide new sites for CNS-directed enzyme therapy. Nat. Med. 2009, 15, 1215–1218. [Google Scholar] [CrossRef] [Green Version]
- Tordo, J.; O’Leary, C.; Antunes, A.; Palomar, N.; Aldrin-Kirk, P.; Basche, M.; Bennett, A.; D’Souza, Z.; Gleitz, H.; Godwin, A.; et al. A novel adeno-associated virus capsid with enhanced neurotropism corrects a lysosomal transmembrane enzyme deficiency. Brain 2018, 141, 2014–2031. [Google Scholar] [CrossRef] [PubMed]
- Zinn, E.; Pacouret, S.; Khaychuk, V.; Turunen, H.T.; Carvalho, L.S.; Andres-Mateos, E.; Shah, S.; Shelke, R.; Maurer, A.C.; Plovie, E.; et al. In Silico Reconstruction of the Viral Evolutionary Lineage Yields a Potent Gene Therapy Vector. Cell Rep. 2015, 12, 1056–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Münch, R.C.; Janicki, H.; Völker, I.; Rasbach, A.; Hallek, M.; Büning, H.; Buchholz, C.J. Displaying High-affinity Ligands on Adeno-associated Viral Vectors Enables Tumor Cell-specific and Safe Gene Transfer. Mol. Ther. 2012, 21, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidsson, M.; Wang, G.; Aldrin-Kirk, P.; Cardoso, T.; Nolbrant, S.; Hartnor, M.; Mudannayake, J.; Parmar, M.; Björklund, T. A systematic capsid evolution approach performed in vivo for the design of AAV vectors with tailored properties and tropism. Proc. Natl. Acad. Sci. USA 2019, 116, 27053–27062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osaki, T.; Shin, Y.; Sivathanu, V.; Campisi, M.; Kamm, R.D. In Vitro Microfluidic Models for Neurodegenerative Disorders. Adv. Healthc. Mater. 2017, 7, 1700489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Lee, B.K.; Jeong, G.S.; Hyun, J.K.; Lee, C.J.; Lee, S.-H. Three-dimensional brain-on-a-chip with an interstitial level of flow and its application as an in vitro model of Alzheimer’s disease. Lab Chip 2015, 15, 141–150. [Google Scholar] [CrossRef]
- Zurina, I.M.; Shpichka, A.; Saburina, I.N.; Kosheleva, N.V.A.; Gorkun, A.A.; Grebenik, E.; Kuznetsova, D.S.; Zhang, D.A.; Rochev, Y.; Butnaru, D.V.; et al. 2D/3D buccal epithelial cell self-assembling as a tool for cell phenotype maintenance and fabrication of multilayered epithelial linings in vitro. Biomed. Mater. 2018, 13, 054104. [Google Scholar] [CrossRef]
- Soriani, M.; Yi, W.; He, A.; Ml, S. Faculty of 1000 evaluation for Microfluidic blood-brain barrier model provides in vivo-like barrier properties for drug permeability screening. F1000 Post Publ. Peer Rev. Biomed. Lit. 2017, 114, 184–194. [Google Scholar]
- Dauth, S.; Maoz, B.M.; Sheehy, S.P.; Hemphill, M.A.; Murty, T.; Macedonia, M.K.; Greer, A.M.; Budnik, B.; Parker, K.K. Neurons derived from different brain regions are inherently different in vitro: A novel multiregional brain-on-a-chip. J. Neurophysiol. 2017, 117, 1320–1341. [Google Scholar] [CrossRef]
- Merkel, S.F.; Andrews, A.; Lutton, E.M.; Mu, D.; Hudry, E.; Hyman, B.T.; Maguire, C.A.; Ramirez, S.H. Trafficking of adeno-associated virus vectors across a model of the blood-brain barrier; a comparative study of transcytosis and transduction using primary human brain endothelial cells. J. Neurochem. 2016, 140, 216–230. [Google Scholar] [CrossRef] [Green Version]
- Donadon, I.; Bussani, E.; Riccardi, F.; Licastro, D.; Romano, G.; Pianigiani, G.; Pinotti, M.; Konstantinova, P.; Evers, M.; Lin, S.; et al. Rescue of spinal muscular atrophy mouse models with AAV9-Exon-specific U1 snRNA. Nucleic Acids Res. 2019, 47, 7618–7632. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.; Smith, D.A.; Morris, L.; Fletcher, C.; Colaço, A.; Huebecker, M.; Tordo, J.; Palomar, N.; Massaro, G.; Henckaerts, E.; et al. AAV9 intracerebroventricular gene therapy improves lifespan, locomotor function and pathology in a mouse model of Niemann–Pick type C1 disease. Hum. Mol. Genet. 2018, 27, 3079–3098. [Google Scholar] [CrossRef] [PubMed]
- Osman, E.Y.; Bolding, M.R.; Villalón, E.; Kaifer, K.A.; Lorson, Z.C.; Tisdale, S.; Hao, Y.; Conant, G.C.; Pires, J.C.; Pellizzoni, L.; et al. Functional characterization of SMN evolution in mouse models of SMA. Sci. Rep. 2019, 9, 9472. [Google Scholar] [CrossRef]
- Amado, D.; Rieders, J.M.; Diatta, F.; Hernandez-Con, P.; Singer, A.; Mak, J.T.; Zhang, J.; Lancaster, E.; Davidson, B.L.; Chen-Plotkin, A.S. AAV-Mediated Progranulin Delivery to a Mouse Model of Progranulin Deficiency Causes T Cell-Mediated Toxicity. Mol. Ther. 2019, 27, 465–478. [Google Scholar] [CrossRef] [Green Version]
- Commisso, B.; Ding, L.; Varadi, K.; Gorges, M.; Bayer, D.; Boeckers, T.M.; Ludolph, A.C.; Kassubek, J.; Müller, O.J.; Roselli, F. Stage-dependent remodeling of projections to motor cortex in ALS mouse model revealed by a new variant retrograde-AAV9. eLife 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Pastor, P.D.H.; Du, X.; Fazal, S.; Davies, A.N.; Gomez, C.M. Targeting the CACNA1A IRES as a Treatment for Spinocerebellar Ataxia Type 6. Cerebellum 2018, 17, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Kang, F.; Wang, S.; Han, M.; Zhang, Z.; Li, J. SIRT1 Activation Attenuates Bone Cancer Pain by Inhibiting mGluR1/5. Cell Mol. Neurobiol. 2019, 39, 1165–1175. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, S.; Lee, N.; Lee, J.; Yu, S.S.; Kim, J.-H.; Kim, S. Intrathecal delivery of recombinant AAV1 encoding hepatocyte growth factor improves motor functions and protects neuromuscular system in the nerve crush and SOD1-G93A transgenic mouse models. Acta Neuropathol. Commun. 2019, 7, 96. [Google Scholar] [CrossRef]
- Wen, D.; Cui, C.; Duan, W.; Wang, W.; Wang, Y.; Liu, Y.; Li, Z.; Li, C. The role of insulin-like growth factor 1 in ALS cell and mouse models: A mitochondrial protector. Brain Res. Bull. 2019, 144, 1–13. [Google Scholar] [CrossRef]
- Gong, Y.; Berenson, A.; Laheji, F.; Gao, G.; Wang, D.; Ng, C.; Volak, A.; Kok, R.; Kreouzis, V.; Dijkstra, I.M.; et al. Intrathecal Adeno-Associated Viral Vector-Mediated Gene Delivery for Adrenomyeloneuropathy. Hum. Gene Ther. 2019, 30, 544–555. [Google Scholar] [CrossRef]
- Janda, E.; LaScala, A.; Carresi, C.; Parafati, M.; Aprigliano, S.; Russo, V.; Savoia, C.; Ziviani, E.; Musolino, V.; Morani, F.; et al. Parkinsonian toxin-induced oxidative stress inhibits basal autophagy in astrocytes via NQO2/quinone oxidoreductase 2: Implications for neuroprotection. Autophagy 2015, 11, 1063–1080. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.M.; Smith, G.; Park, E.; Cao, H.; Brown, E.; Hayes, M.A.; Beagan, J.; McLean, J.R.; Izen, S.C.; Perez-Torres, E.; et al. Glucocerebrosidase gene therapy prevents α-synucleinopathy of midbrain dopamine neurons. Neurobiol. Dis. 2015, 82, 495–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, J.B.; Nash, K.R.; Placides, D.; Moran, P.; Selenica, M.-L.B.; Abuqalbeen, F.; Ratnasamy, K.; Slouha, N.; Rodriguez-Ospina, S.; Savlia, M.; et al. Sustained Arginase 1 Expression Modulates Pathological Tau Deposits in a Mouse Model of Tauopathy. J. Neurosci. 2015, 35, 14842–14860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, L.; Xu, Y.; Zhuo, R.; Wang, T.; Wang, K.; Huang, R.; Wang, D.; Gao, Y.; Zhu, Y.; Sheng, X.; et al. Soluble TREM2 ameliorates pathological phenotypes by modulating microglial functions in an Alzheimer’s disease model. Nat. Commun. 2019, 10, 1365. [Google Scholar] [CrossRef] [Green Version]
- Stoker, T.B.; Torsney, K.M.; Barker, R.A. Emerging Treatment Approaches for Parkinson’s Disease. Front. Mol. Neurosci. 2018, 12, 693. [Google Scholar] [CrossRef] [Green Version]
- Gutekunst, C.-A.; Tung, J.K.; McDougal, M.E.; Gross, R.E. C3 transferase gene therapy for continuous conditional RhoA inhibition. Neurosci. 2016, 339, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.; Muramatsu, S.-I.; Shimizu, N.; Yokoyama, S.; Hirai, H.; Yamada, K.; Liu, H.-X.; Higashida, C.; Hashii, M.; Higashida, A.; et al. Dopamine release via the vacuolar ATPase V0 sector c-subunit, confirmed in N18 neuroblastoma cells, results in behavioral recovery in hemiparkinsonian mice. Neurochem. Int. 2012, 61, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Higashida, H.; Yokoyama, S.; Tsuji, C.; Muramatsu, S.-I. Neurotransmitter release: Vacuolar ATPase V0 sector c-subunits in possible gene or cell therapies for Parkinson’s, Alzheimer’s, and psychiatric diseases. J. Physiol. Sci. 2016, 67, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.-H.; Ji, W.-L.; Xu, D.-E.; Yao, P.-P.; Zhao, X.-Y.; Wang, Z.-T.; Fang, L.-P.; Huang, R.; Lan, L.-J.; Chen, J.-B.; et al. Inhibition of sphingomyelin synthase 1 ameliorates alzheimer-like pathology in APP/PS1 transgenic mice through promoting lysosomal degradation of BACE1. Exp. Neurol. 2019, 311, 67–79. [Google Scholar] [CrossRef]
- Stoica, L.; Sena-Esteves, M. Adeno Associated Viral Vector Delivered RNAi for Gene Therapy of SOD1 Amyotrophic Lateral Sclerosis. Front. Mol. Neurosci. 2016, 9, 1971. [Google Scholar] [CrossRef] [Green Version]
- Frakes, A.E.; Braun, L.; Ferraiuolo, L.; Guttridge, D.C.; Kaspar, B.K. Additive amelioration of ALS by co-targeting independent pathogenic mechanisms. Ann. Clin. Transl. Neurol. 2017, 4, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.-M.; Liu, D.-D.; Li, Z.; Ling, C.; Mei, Y.-A. Neuritin Enhances Synaptic Transmission in Medial Prefrontal Cortex in Mice by Increasing CaV3.3 Surface Expression. Cereb. Cortex 2017, 27, 3842–3855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evers, M.; Miniarikova, J.; Juhas, S.; Vallès, A.; Bohuslavova, B.; Juhasova, J.; Skalnikova, H.K.; Vodicka, P.; Valekova, I.; Brouwers, C.; et al. AAV5-miHTT Gene Therapy Demonstrates Broad Distribution and Strong Human Mutant Huntingtin Lowering in a Huntington’s Disease Minipig Model. Mol. Ther. 2018, 26, 2163–2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cideciyan, A.V.; Sudharsan, R.; Dufour, V.L.; Massengill, M.T.; Iwabe, S.; Swider, M.; Lisi, B.; Sumaroka, A.; Marinho, L.F.; Appelbaum, T.; et al. Mutation-independent rhodopsin gene therapy by knockdown and replacement with a single AAV vector. Proc. Natl. Acad. Sci. USA 2018, 115, E8547–E8556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Ahn, H.J. Anti-EpCAM-conjugated adeno-associated virus serotype 2 for systemic delivery of EGFR shRNA: Its retargeting and antitumor effects on OVCAR3 ovarian cancer in vivo. Acta Biomater. 2019, 91, 258–269. [Google Scholar] [CrossRef]
- Zharikov, A.; Bai, Q.; De Miranda, B.R.; Van Laar, A.; Greenamyre, J.T.; Burton, E.A. Long-term RNAi knockdown of α-synuclein in the adult rat substantia nigra without neurodegeneration. Neurobiol. Dis. 2019, 125, 146–153. [Google Scholar] [CrossRef]
- Li, H.; Wan, C.; Li, F. Recombinant adeno-associated virus-, polyethylenimine/plasmid- and lipofectamine/carboxyfluorescein-labeled small interfering RNA-based transfection in retinal pigment epithelial cells with ultrasound and/or SonoVue. Mol. Med. Rep. 2015, 11, 3609–3614. [Google Scholar] [CrossRef] [Green Version]
- Komor, A.C.; Badran, A.H.; Liu, D.R. CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes. Cell 2017, 169, 559. [Google Scholar] [CrossRef] [Green Version]
- Reyon, D.; Tsai, S.Q.; Khayter, C.; Foden, J.A.; Sander, J.D.; Joung, J.K. FLASH assembly of TALENs for high-throughput genome editing. Nat. Biotechnol. 2012, 30, 460–465. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Raikwar, S.; Thangavel, R.; Dubova, I.; Selvakumar, G.P.; Ahmed, M.E.; Kempuraj, D.; Zaheer, S.A.; Iyer, S.S.; Zaheer, A. Targeted Gene Editing of Glia Maturation Factor in Microglia: A Novel Alzheimer’s Disease Therapeutic Target. Mol. Neurobiol. 2018, 56, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Gaj, T.; Ojala, D.S.; Ekman, F.K.; Byrne, L.C.; Limsirichai, P.; Schaffer, D.V. In vivo genome editing improves motor function and extends survival in a mouse model of ALS. Sci. Adv. 2017, 3, eaar3952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekman, F.K.; Ojala, D.S.; Adil, M.M.; Lopez, P.A.; Schaffer, D.V.; Gaj, T. CRISPR-Cas9-Mediated Genome Editing Increases Lifespan and Improves Motor Deficits in a Huntington’s Disease Mouse Model. Mol. Ther. Nucleic Acids 2019, 17, 829–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.; Sun, J.; Wang, F.; Luo, X.; Feng, J.; Gu, Q.; Liu, T.; Sun, X. MicroRNA Expression Patterns Involved in Amyloid Beta–Induced Retinal Degeneration. Investig. Opthalmol. Vis. Sci. 2017, 58, 1726–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Liu, L.; Meng, J. MicroRNA-125b promotes neurons cell apoptosis and Tau phosphorylation in Alzheimer’s disease. Neurosci. Lett. 2017, 661, 57–62. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Adachi, H.; Katsuno, M.; Minamiyama, M.; Jiang, Y.-M.; Huang, Z.; Doi, H.; Matsumoto, S.; Kondo, N.; Iida, M.; et al. Viral delivery of miR-196a ameliorates the SBMA phenotype via the silencing of CELF2. Nat. Med. 2012, 18, 1136–1141. [Google Scholar] [CrossRef]
- Boissonneault, V.; Plante, I.; Rivest, S.; Provost, P. MicroRNA-298 and microRNA-328 regulate expression of mouse beta-amyloid precursor protein-converting enzyme 1. J. Boil. Chem. 2008, 284, 1971–1981. [Google Scholar] [CrossRef] [Green Version]
- Tuszynski, M.H.; Yang, J.H.; Barba, D.; Hoi-Sang, U.; Bakay, R.A.; Pay, M.M.; Masliah, E.; Conner, J.M.; Kobalka, P.; Roy, S.; et al. Nerve Growth Factor Gene Therapy: Activation of Neuronal Responses in Alzheimer Disease. JAMA Neurol. 2015, 72, 1139–1147. [Google Scholar] [CrossRef]
- Quintino, L.; Avallone, M.; Brännstrom, E.; Kavanagh, P.; Lockowandt, M.; Jareño, P.G.; Breger, L.S.; Lundberg, C. GDNF-mediated rescue of the nigrostriatal system depends on the degree of degeneration. Gene Ther. 2018, 26, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Islamov, R.; Rizvanov, A.; Mukhamedyarov, M.; Salafutdinov, I.; Garanina, E.; Fedotova, V.; Solovyeva, V.V.; Mukhamedshina, Y.; Safiullov, Z.; Izmailov, A.; et al. Symptomatic Improvement, Increased Life-Span and Sustained Cell Homing in Amyotrophic Lateral Sclerosis After Transplantation of Human Umbilical Cord Blood Cells Genetically Modified with Adeno-Viral Vectors Expressing a Neuro-Protective Factor and a Neural Cell Adhesion Molecule. Curr. Gene Ther. 2015, 15, 266–276. [Google Scholar]
- Schubert, T.; Wissinger, B. Restoration of synaptic function in sight for degenerative retinal disease. J. Clin. Investig. 2015, 125, 2572–2575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-C.; Ma, N.-X.; Pei, Z.-F.; Wu, Z.; Do-Monte, F.H.; Keefe, S.; Yellin, E.; Chen, M.S.; Yin, J.-C.; Lee, G.; et al. A NeuroD1 AAV-Based Gene Therapy for Functional Brain Repair after Ischemic Injury through In Vivo Astrocyte-to-Neuron Conversion. Mol. Ther. 2020, 28, 217–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginn, S.L.; Amaya, A.K.; Alexander, I.E.; Edelstein, M.; Abedi, M.R. Gene therapy clinical trials worldwide to 2017: An update. J. Gene Med. 2018, 20, e3015. [Google Scholar] [CrossRef]
- Tan, J.-K.Y.; Sellers, D.L.; Pham, B.; Pun, S.H.; Horner, P.J. Non-Viral Nucleic Acid Delivery Strategies to the Central Nervous System. Front. Mol. Neurosci. 2016, 9, 41. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-P.; Ding, H.; Lu, J.-D.; Tang, Y.-H.; Deng, B.-X.; Deng, C.-Q. Autophagy in cerebral ischemia and the effects of traditional Chinese medicine. J. Integr. Med. 2015, 13, 289–296. [Google Scholar] [CrossRef]


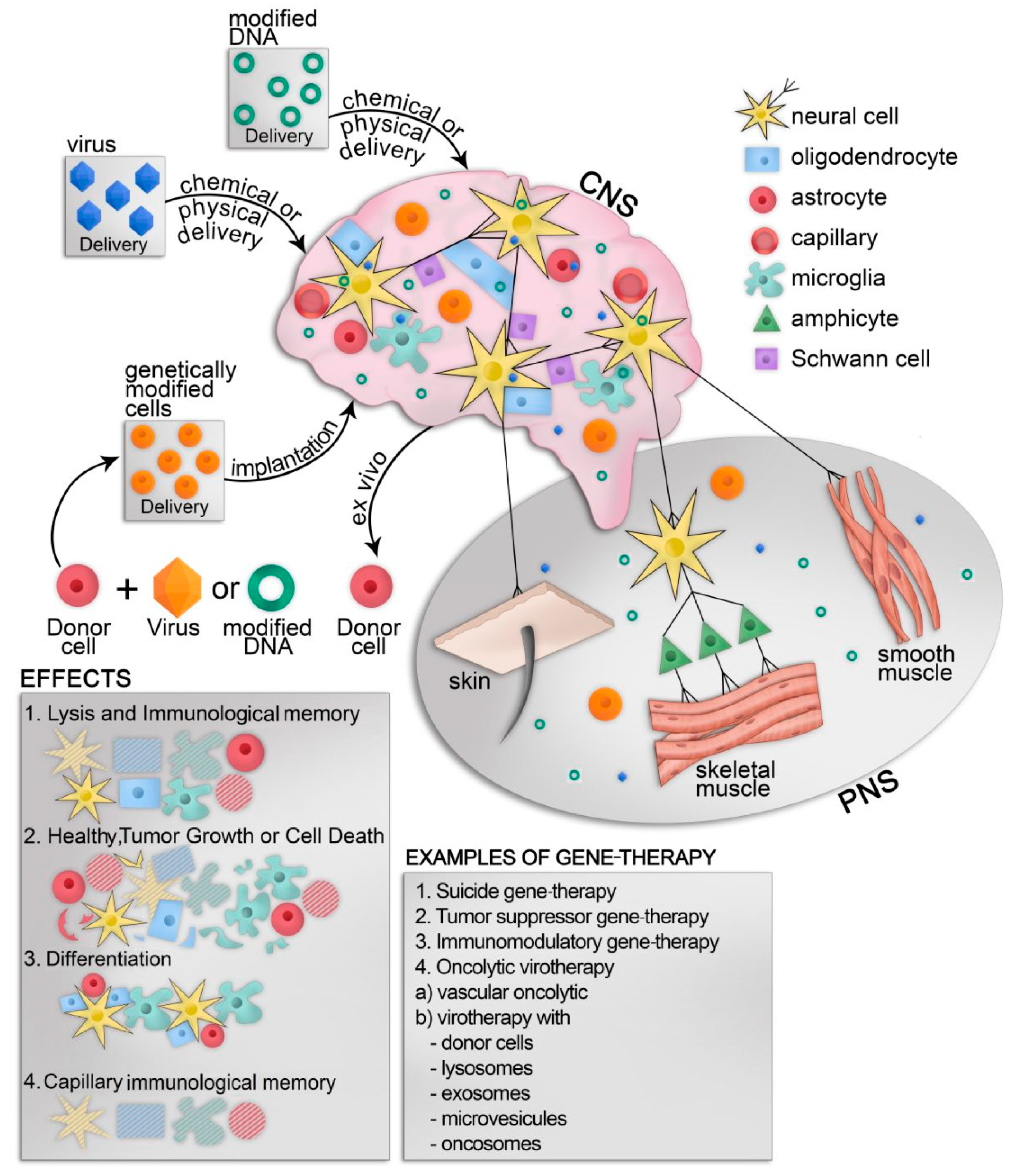{kind=link}
| Vectors | Transgene | Disease | Phase | Patients Enrolled | Outcome | Study name | Clinicaltrail.gov | Status |
|---|---|---|---|---|---|---|---|---|
| AAV | Beta-nerve growth factor (NGF) | Alzheimer’s disease | 1 | 10 | A Phase I, Dose-Escalating Study to Assess the Safety and Tolerability of CERE-110 [Adeno-Associated Virus (AAV)-Based Vector-Mediated Delivery of Beta-Nerve Growth Factor (NGF)] in Subjects with Mild to Moderate Alzheimer’s Disease | CERE-110 in Subjects With Mild to Moderate Alzheimer’s Disease | NCT00087789 CERE-110 2.0 × 1010 vg, CERE-110 1.0 × 1011 vg, CERE-110 2.0 × 1011 vg | Completed |
| AAV | Beta-nerve growth factor (NGF) | Alzheimer’s disease | 2 | 49 | A Double-Blind, Placebo-Controlled (Sham Surgery), Randomized, Multicenter Study Evaluating CERE-110 Gene Delivery in Subjects with Mild to Moderate Alzheimer’s Disease | Randomized, Controlled Study Evaluating CERE-110 in Subjects With Mild to Moderate Alzheimer’s Disease | NCT00876863 CERE-110 2.0 × 1011 vg | Completed |
| AAV | Neurotrophic (growth) factor (Neurturin) | Parkinson’s disease | 1/2 | 60 est/57 fact | A Phase 1/2 Trial Assessing the Safety and Efficacy of Bilateral Intraputaminal and Intranigral Administration of CERE-120 (Adeno-Associated Virus Serotype 2 [AAV2]-Neurturin [NTN]) in Subjects with Idiopathic Parkinson’s Disease | Safety and Efficacy of CERE-120 in Subjects With Parkinson’s Disease | NCT00985517 CERE-120 2.4 × 1012 vg | Completed |
| AAV | Glutamic acid decarboxylase (GAD) | Parkinson’s disease | 1 | 12 | Phase I Study of Subthalamic GAD Gene Transfer in Medically Refractory Parkinson’s Disease | Safety Study of Subthalamic Nucleus Gene Therapy for Parkinson’s Disease | NCT00195143 | Completed |
| AAV | Glutamic acid decarboxylase (GAD) | Parkinson’s disease | 2 | 44 (est) | Phase 2 Safety and Efficacy Study Evaluating Glutamic Acid Decarboxylase Gene Transfer to Subthalamic Nuclei in Subjects with Advanced Parkinson’s Disease | Study of AAV-GAD Gene Transfer Into the Subthalamic Nucleus for Parkinson’s Disease | NCT00643890 One-time bilateral administration of rAAV-GAD at 1 × 1012 vector genomes in 35 uL | Terminated (Financial reasons) |
| AAV | Glutamic acid decarboxylase GAD | Parkinson’s disease | 40 est/0 fact | N/A | Long Term Follow-Up Study for rAAV-GAD Treated Subjects | NCT01301573 | Terminated (Financial reasons) | |
| AAV | Aromatic L-amino acid decarboxylase (hAADC-2) | Parkinson’s disease | 1 | 10 | A Phase1 Open-Label Safety Study of Intrastriatal Infusion of Adeno-Associated Virus Encoding Human Aromatic L-Amino Acid Decarboxylase (AAV-hAADC-2) in Subjects with Parkinson’s Disease [AAV-hAADC-2-003] | A Study of AAV-hAADC-2 in Subjects With Parkinson’s Disease | NCT00229736 9 × 1010 vector genomes (vg) of AAV-hAADC-2 in a single dose of 200 µL bilaterally infused over 4 striatal targets 3 × 1011 vector genomes (vg) of AAV-hAADC-2 in a single dose of 200 µL bilaterally infused over 4 striatal targets | Completed |
| AAV | Aromatic L-amino acid decarboxylase (hAADC-2) | Parkinson’s disease | 1/2 | 6 est/2fact | A Phase I/II Study of Intra-Putaminal Infusion of Adeno-Associated Virus Encoding Human Aromatic L-Amino Acid Decarboxylase in Subjects with Parkinson’s Disease | AADC Gene Therapy for Parkinson’s Disease | NCT02418598 AAV-hAADC-2 is administered via bilateral intra-putaminal infusion. The number of vector genomes (vg) cohort 1: 3 × 1011 vg/subject cohort 2: 9 × 1011 vg/subject. | Terminated (Another clinical study for regulatory approval is planned) |
| AAV1 | Neurotrophin factor 3 (NTF3) | Charcot–Marie–Tooth disease | 1/2a | 9est/0 fact | Phase I/II a Trial Evaluating scAAV1.tMCK.NTF3 for Treatment of Charcot–Marie–Tooth Neuropathy Type 1A (CMT1A) | Phase I/II a Trial of scAAV1.tMCK.NTF3 for Treatment of CMT1A | NCT03520751 N = 3: intramuscular injection of (scAAV1.tMCK.NTF3) distributed bilaterally between both limbs at low dose (2 × 1012 vg/kg). N = 6: intramuscular injection of (scAAV1.tMCK.NTF3) distributed bilaterally between both limbs at low dose (6 × 1012 vg/kg) | Not yet recruiting |
| AAV2 | Neurotrophic (growth) factor (Neurturin) | Parkinson’s disease | 2 | 58 est/51 fact | Multicenter, Randomized, Double-Blind, Sham Surgery-Controlled Study of CERE-120 (Adeno-Associated Virus Serotype 2 [AAV2]-Neurturin [NTN]) to Assess the Efficacy and Safety of Bilateral Intraputaminal (IPu) Delivery in Subjects with Idiopathic Parkinson’s Disease | Double-Blind, Multicenter, Sham Surgery Controlled Study of CERE-120 in Subjects With Idiopathic Parkinson’s Disease | NCT00400634, CERE-120, bilaterally: 5.4 × 1011 vg | Completed |
| AAV2 | Neurotrophic (growth) factor (Neurturin) | Parkinson’s disease | 1 | 12 est | A Phase I, Open-Label Study of CERE-120 (Adeno-Associated Virus Serotype 2 [AAV2]-Neurturin [NTN] to Assess the Safety and Tolerability of Intrastriatal Delivery to Subjects with Idiopathic Parkinson’s Disease | Safety of CERE-120 (AAV2-NTN) in Subjects With Idiopathic Parkinson’s Disease | NCT00252850 | Completed |
| AAV2 | Human aromatic L-amino acid decarboxylase (AADC) gene | Parkinson’s disease | 1 | 15 est/10 fact | An Open-label Safety and Efficacy Study of VY-AADC01 Administered by MRI-Guided Convective Infusion into the Putamen of Subjects with Parkinson’s Disease with Fluctuating Responses to Levodopa | Safety Study of AADC Gene Therapy (VY-AADC01) for Parkinson’s Disease (AADC) | NCT01973543 VY-AADC01; Single dose, neurosurgically-infused, bilaterally into the striatum: 7.5 × 1011 vg, 1.5 × 1012 vg, 4.7 × 1012 vg | Active, not recruiting |
| AAV2 | Human aromatic L-amino acid decarboxylase (AADC) gene | Parkinson’s disease | 2 | 42 est | A Randomized, Placebo Surgery Controlled, Double-Blinded, Multi-center, Phase 2 Clinical Trial, Evaluating the Efficacy and Safety of VY-AADC02 in Advanced Parkinson’s Disease with Motor Fluctuations | VY-AADC02 for Parkinson’s Disease With Motor Fluctuations | NCT03562494, VY-AADC02 infusion, 2.5 × 1012 | Recruiting |
| AAV2 | Human aromatic L-amino acid decarboxylase (AADC) gene | Parkinson’s disease | 50 est | An Observational, Long-Term Extension Study for Participants of Prior VY-AADC01 or VY-AADC02 Clinical Studies | Observational, Long-term, Extension Study for Participants of Prior VY-AADC01 or VY-AADC02 Studies | NCT03733496, Participants who received VY-AADC01 or VY-AADC02 | Enrolling by invitation | |
| AAV2 | Human CLN2 | Late infantile neuronal ceroid lipofuscinosis (LINCL) | 1 | 11 est/10 fact | Administration of a Replication Deficient Adeno-Associated Virus Gene Transfer Vector Expressing the Human CLN2 cDNA to the Brain of Children with Late Infantile Neuronal Ceroid Lipofuscinosis | Safety Study of a Gene Transfer Vector for Children With Late Infantile Neuronal Ceroid Lipofuscinosis | NCT00151216 AAV2CUhCLN2, N = 5, 3 × 1012 N = 6, 3 × 1012 | Active, not recruiting |
| AAV2 | Glial cell line-derived neurotrophic factor (GDNF) | Parkinson’s disease | 1 | 28 est/25 fact | A Phase 1 Open-Label Dose Escalation Safety Study of Convection Enhanced Delivery (CED) of Adeno-Associated Virus Encoding Glial Cell Line-Derived Neurotrophic Factor (AAV2-GDNF) in Subjects with Advanced Parkinson’s Disease | AAV2-GDNF for Advanced Parkinson s Disease | NCT01621581 9 × 1010 vg, 3 × 1011 vg, 9 × 1011 vg 3 × 1012 vg | Active, not recruiting |
| AAV2 | Human ND4 | Leber’s congenital amaurosis | 3 | 90 est | Efficacy and Safety of Bilateral Intravitreal Injection of GS010: A Randomized, Double-Masked, Placebo-Controlled Trial in Subjects Affected with G11778A ND4 Leber’s Hereditary Optic Neuropathy for Up to One Year | Safety and Efficacy Study of Gene Therapy for The Treatment of Leber’s Hereditary Optic Neuropathy | NCT03293524, GS010, IVT eye, 9 × 1010 vg | Active, not recruiting |
| AAV2 | Human ND4 | Leber’s congenital amaurosis | 2 + 3 | 159 est/48 fact | Safety and Efficacy Study of Gene Therapy for The Treatment of Leber’s Hereditary Optic Neuropathy | NCT03153293, Single IVT injection, 1 × 1010 vg/0.05 mL | Active, not recruiting | |
| AAV2 | Human ND4 | Leber’s hereditary optic neuropathy | 3 | 36 est | A Randomized, Double-Masked, Sham-Controlled Clinical Trial to Evaluate the Efficacy of a Single Intravitreal Injection of GS010 in Subjects Affected for 6 Months or Less by LHON Due to the G11778A Mutation in the Mitochondrial ND4 Gene | Efficacy Study of GS010 for the Treatment of Vision Loss up to 6 Months From Onset in LHON Due to the ND4 Mutation (RESCUE) | NCT02652767, GS010 | Active, not recruiting |
| AAV2 | Human ND4 | Leber’s hereditary optic neuropathy | 3 | 37 est/36 fact | Randomized, Double-Masked, Sham-Controlled Clinical Trial to Evaluate the Efficacy of a Single Intravitreal Injection of GS010 in Subjects Affected for More Than 6 Months and to 12 Months by LHON Due to the G11778A Mutation in the ND4 Gene | Efficacy Study of GS010 for Treatment of Vision Loss From 7 Months to 1 Year From Onset in LHON Due to the ND4 Mutation (REVERSE) (REVERSE) | NCT02652780, rAAV2/2-ND4 intravitreal, 9 × 1010 vg | Completed |
| AAV2 | Human ND4 | Leber’s hereditary optic neuropathy | 74 est | Long-Term Follow-Up of ND4 LHON Subjects Treated with GS010 Ocular Gene Therapy in the RESCUE or REVERSE Phase III Clinical Trials | RESCUE and REVERSE Long-term Follow-up (RESCUE/REVERSE) | NCT03406104, GS010, intravitreal injection | Recruiting | |
| AAV2 | Human ND4 | Leber’s hereditary optic neuropathy | EAP Single Patient: Safety of Bilateral Intravitreal Injection of GS010 in a Single Subject Affected with G11778A ND4 Leber’s Hereditary Optic Neuropathy | EAP_GS010_single Patient | NCT03672968 | Available | ||
| AAV2 | Human ND4 | Leber’s hereditary optic neuropathy | 1 | 30 est/27 fact | An Open-Label Dose Escalation Study of an Adeno-Associated Virus Vector (scAAV2-P1ND4v2) for Gene Therapy of Leber’s Hereditary Optic Neuropathy (LHON) Caused by the G11778A Mutation in Mitochondrial DNA | Safety Study of an Adeno-associated Virus Vector for Gene Therapy of Leber’s Hereditary Optic Neuropathy (LHON) | NCT02161380 scAAV2-P1ND4v2 intravitreal: 1.18 × 109 vg 5.81 × 109 vg 2.4 × 1010 vg 1.0 × 1011 vg | Recruiting |
| AAV9 | CLN6 | CLN6, Batten disease | 1/2a | 13 est/6 fact | Phase I/II a Gene Transfer Clinical Trial for Variant Late Infantile Neuronal Ceroid Lipofuscinosis, Delivering the CLN6 Gene by Self-Complementary AAV9 | Gene Therapy for Children With Variant Late Infantile Neuronal Ceroid Lipofuscinosis 6 (vLINCL6) Disease | NCT02725580 AT-GTX-501 (scAAV9.CB.CLN6) | Active, not recruiting |
| AAV9 | CLN3 | CLN3, neuronal ceroid- lipofuscinosis | 1/2a | 7 est | Phase I/II a Gene Transfer Clinical Trial for Juvenile Neuronal Ceroid Lipofuscinosis, Delivering the CLN3 Gene by Self-Complementary AAV9 | Gene Therapy for Children With CLN3 Batten Disease | NCT03770572 AT-GTX-502 High dose Low dose | Recruiting |
| AAV9 | Human survival motor neuron (SMN) | Spinal muscular atrophy type 1 (SMA1) | 1 | 15 est/9 fact | Phase I Gene Transfer Clinical Trial for Spinal Muscular Atrophy Type 1 Delivering AVXS-101 | Gene Transfer Clinical Trial for Spinal Muscular Atrophy Type 1 | NCT02122952 iv N = 3, 6.7 × 1013 vg/kg N = 3, 2.0 × 1014 vg/kg | Completed |
| AAV9 | Human survival motor neuron (SMN) | SMA | 1 | 51 est/27 fact | Phase I, Open-Label, Dose Comparison Study of AVXS-101 for Sitting but Non-Ambulatory Patients with Spinal Muscular Atrophy | Study of Intrathecal Administration of Onasemnogene Abeparvovec-xioi for Spinal Muscular Atrophy (STRONG) | NCT03381729 Intrathecal Onasemnogene abeparvovec-xioi 6.0 × 1013 vg 1.2 × 1014 vg 2.4 × 1014 vg | Suspended |
| AAV9 | Human survival motor neuron (SMN) | Spinal muscular atrophy type 1 (SMA1) | 3 | 22est/15 fact | Phase 3, Open-Label, Single-Arm, Single-Dose Gene Replacement Therapy Clinical Trial for Patients with Spinal Muscular Atrophy Type 1 With One or Two SMN2 Copies Delivering AVXS-101 by Intravenous Infusion | Gene Replacement Therapy Clinical Trial for Patients With Spinal Muscular Atrophy Type 1 (STR1VE) | NCT03306277 iv Onasemnogene abeparvovec-xioi | Completed |
| AAV9 | Human survival motor neuron (SMN) | Spinal muscular atrophy type 1 (SMA1) | 3 | 44 est/30 fact | A Global Study of a Single, One-Time Dose of AVXS-101 Delivered to Infants with Genetically Diagnosed and Pre-Symptomatic Spinal Muscular Atrophy with Multiple Copies of SMN2 | Pre-Symptomatic Study of Intravenous Onasemnogene Abeparvovec-xioi in Spinal Muscular Atrophy (SMA) for Patients With Multiple Copies of SMN2 (SPR1NT) | NCT03505099 iv, Onasemnogene abeparvovec-xioi 1.1 × 1014 vg/kg | Active, not recruiting |
| AAV9 | Human survival motor neuron (SMN) | Spinal muscular atrophy (SMA) type 1 | 3 | 33est/30 fact | European, Phase 3, Open-Label, Single-Arm, Single-Dose Gene Replacement Therapy Clinical Trial for Patients with Spinal Muscular Atrophy Type 1 with One or Two SMN2 Copies Delivering AVXS-101 by Intravenous Infusion | Single-Dose Gene Replacement Therapy Clinical Trial for Patients With Spinal Muscular Atrophy Type 1 (STRIVE-EU) | NCT03461289 iv Onasemnogene Abeparvovec-xioi | Active, not recruiting |
| AAV10 | Human SGSH and SUMF1 cDNAs | Sanfilippo type A syndrome | 1 + 2 | 4 est | An Open-label, Single Arm, Monocentric, Phase I/II Clinical Study of Intracerebral Administration of Adeno-associated Viral Vector Serotype 10 Carrying the Human SGSH and SUMF1 cDNAs for the Treatment of Sanfilippo Type A Syndrome | Intracerebral Gene Therapy for Sanfilippo Type A Syndrome | NCT01474343 Intracerebral SAF-301 | Completed |
| AAV10 | Human SGSH and SUMF1 cDNAs | Sanfilippo type A syndrome | 1 + 2 | 4 est/ | Long-Term Follow-Up of Patient with Sanfilippo Type A Syndrome Who Have Previously Been Treated in the P1-SAF-301 Clinical Study Evaluating the Tolerability and Safety of the Intracerebral Administration of SAF-301 | Long-term Follow-up of Sanfilippo Type A Patients Treated by Intracerebral SAF-301 Gene Therapy | NCT02053064 Intracerebral SAF-301 Long term effect | Completed |
| AAVrh10 | Human apolipoprotein E2 (APOE2) | Alzheimer’s disease | 1 | 15 est/0 fact | Maximum Tolerated Dose of Intracisternal delivery of AAVrh.10hAPOE2 (no results) | Gene Therapy for APOE4 Homozygote of Alzheimer’s Disease | NCT03634007 AAVrh.10hAPOE2: 8.0 × 1010 gc/kg, 2.5 × 1011 gc/kg 8.0 × 1011 gc/kg | Recruiting |
| AAVrh10 | Human CLN2 | Late-infantile neuronal ceroid Lipofuscinosis | 1 | 25 est/16fact | Direct CNS Administration of a Replication Deficient Adeno-associated Virus Gene Transfer Vector Serotype rh.10 Expressing the Human CLN2 cDNA to Children with Late Infantile Neuronal Ceroid Lipofuscinosis (LINCL) | Safety Study of a Gene Transfer Vector (Rh.10) for Children With Late Infantile Neuronal Ceroid Lipofuscinosis (LINCL) | NCT01161576 AAVrh10.CUhCLN2, N = 6, 9 × 1011 N = 10, 2.85 × 1011 | Active, not recruiting |
| AAVrh10 | Human CLN2 | Late infantile neuronal ceroid lipofuscinosis | 1 + 2 | 16 est/8 fact | Improved Results on Weill Cornell LINCL Scale and Mullen Scale (no results posted) | AAVRh.10 Administered to Children With Late Infantile Neuronal Ceroid Lipofuscinosis | NCT01414985 AAVrh10.CUhCLN2, N = 2, 9 × 1011 N = 6, 2.85 × 1011 | Active, not recruiting |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mijanović, O.; Branković, A.; Borovjagin, A.V.; Butnaru, D.V.; Bezrukov, E.A.; Sukhanov, R.B.; Shpichka, A.; Timashev, P.; Ulasov, I. Battling Neurodegenerative Diseases with Adeno-Associated Virus-Based Approaches. Viruses 2020, 12, 460. https://doi.org/10.3390/v12040460
Mijanović O, Branković A, Borovjagin AV, Butnaru DV, Bezrukov EA, Sukhanov RB, Shpichka A, Timashev P, Ulasov I. Battling Neurodegenerative Diseases with Adeno-Associated Virus-Based Approaches. Viruses. 2020; 12(4):460. https://doi.org/10.3390/v12040460
Chicago/Turabian StyleMijanović, Olja, Ana Branković, Anton V. Borovjagin, Denis V. Butnaru, Evgeny A. Bezrukov, Roman B. Sukhanov, Anastasia Shpichka, Peter Timashev, and Ilya Ulasov. 2020. "Battling Neurodegenerative Diseases with Adeno-Associated Virus-Based Approaches" Viruses 12, no. 4: 460. https://doi.org/10.3390/v12040460
APA StyleMijanović, O., Branković, A., Borovjagin, A. V., Butnaru, D. V., Bezrukov, E. A., Sukhanov, R. B., Shpichka, A., Timashev, P., & Ulasov, I. (2020). Battling Neurodegenerative Diseases with Adeno-Associated Virus-Based Approaches. Viruses, 12(4), 460. https://doi.org/10.3390/v12040460