Cell-Mediated Responses to Human Metapneumovirus Infection


Abstract
:1. Introduction
1.1. Airway Epithelial Cells (AECs)
1.2. Alveolar Macrophages (AMφ)
1.3. Neutrophils
1.4. Dendritic Cells (DCs)
1.5. Natural Killer Cells (NKs)
1.5.1. CD4+ T Cells
1.5.2. CD8+ T cells
1.6. B Cells
2. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Troeger, C.; Forouzanfar, M.; Rao, P.C.; Khalil, I.; Brown, A.; Swartz, S.; Fullman, N.; Mosser, J.; Thompson, R.L.; Reiner, R.C.; et al. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory tract infections in 195 countries: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 1133–1161. [Google Scholar] [CrossRef] [Green Version]
- Divarathna, M.V.; Rafeek, R.A.; Noordeen, F. A review on epidemiology and impact of human metapneumovirus infections in children using TIAB search strategy on PubMed and PubMed Central articles. Rev. Med. Virol. 2019, 30. [Google Scholar] [CrossRef] [PubMed]
- Hoogen, B.G.V.D.; De Jong, J.C.; Groen, J.; Kuiken, T.; De Groot, R.; Fouchier, R.A.; Osterhaus, A. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat. Med. 2001, 7, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Boivin, G.; Abed, Y.; Pelletier, G.; Ruel, L.; Moisan, D.; Côté, S.; Peret, T.C.T.; Erdman, D.D.; Anderson, L.J. Virological Features and Clinical Manifestations Associated with Human Metapneumovirus: A New Paramyxovirus Responsible for Acute Respiratory-Tract Infections in All Age Groups. J. Infect. Dis. 2002, 186, 1330–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterhaus, A.; Fouchier, R. Human metapneumovirus in the community. Lancet 2003, 361, 890–891. [Google Scholar] [CrossRef]
- Morris, J.A.; Blount, R.E.; Savage, R.E. Recovery of Cytopathogenic Agent from Chimpanzees with Goryza. Exp. Boil. Med. 1956, 92, 544–549. [Google Scholar] [CrossRef]
- Rudan, I.; O’Brien, K.L.; Nair, H.; Liu, L.; Theodoratou, E.; Qazi, S.; Lukšić, I.; Walker, C.L.F.; Black, R.E.; Campbell, H. Epidemiology and etiology of childhood pneumonia in 2010: Estimates of incidence, severe morbidity, mortality, underlying risk factors and causative pathogens for 192 countries. J. Glob. Health 2013, 3, 010401. [Google Scholar]
- Ulloa-Gutierrez, R. Metapneumovirus and its place in childhood. Paediatr. Child Health 2003, 8, 613–615. [Google Scholar] [CrossRef]
- Crowe, J.E. Human Metapneumovirus as a Major Cause of Human Respiratory Tract Disease. Pediatr. Infect. Dis. J. 2004, 23, S215–S221. [Google Scholar] [CrossRef]
- Hoogen, B.G.V.D.; Osterhaus, D.M.E.; Fouchier, R.A.M. Clinical impact and diagnosis of human metapneumovirus infection. Pediatr. Infect. Dis. J. 2004, 23, S25–S32. [Google Scholar] [CrossRef]
- Williams, J.V. Human metapneumovirus: An important cause of respiratory disease in children and adults. Curr. Infect. Dis. Rep. 2005, 7, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.M.; Zhu, Y.; Griffin, M.R.; Weinberg, G.A.; Hall, C.B.; Szilagyi, P.G.; Staat, M.A.; Iwane, M.; Prill, M.M.; Williams, J.V.; et al. Burden of human metapneumovirus infection in young children. N. Engl. J. Med. 2013, 368, 633–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, J.E.; Khuri-Bulos, N.; Faouri, S.; Shehabi, A.; Johnson, M.; Wang, L.; Fonnesbeck, C.; Williams, J.V.; Halasa, N. Human Metapneumovirus Infection in Jordanian Children: Epidemiology and Risk Factors for Severe Disease. Pediatr. Infect. Dis. J. 2015, 34, 1335–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.J.; Simões, E.A.; Buttery, J.P.; Dennehy, P.; Domachowske, J.; Jensen, K.; Lieberman, J.M.; Losonsky, G.A.; Yogev, R. Prevalence and Characteristics of Human Metapneumovirus Infection Among Hospitalized Children at High Risk for Severe Lower Respiratory Tract Infection. J. Pediatr. Infect. Dis. Soc. 2012, 1, 212–222. [Google Scholar] [CrossRef]
- Hoogen, B.G.V.D.; Van Doornum, G.J.J.; Fockens, J.C.; Cornelissen, J.J.; Beyer, W.E.P.; De Groot, R.; Osterhaus, A.; Fouchier, R.A.M. Prevalence and Clinical Symptoms of Human Metapneumovirus Infection in Hospitalized Patients. J. Infect. Dis. 2003, 188, 1571–1577. [Google Scholar] [CrossRef] [Green Version]
- Bosis, S.; Esposito, S.; Niesters, H.; Crovari, P.; Osterhaus, A.; Principi, N. Impact of human metapneumovirus in childhood: Comparison with respiratory syncytial virus and influenza viruses. J. Med. Virol. 2004, 75, 101–104. [Google Scholar] [CrossRef]
- Boivin, G.; De Serres, G.; Côté, S.; Gilca, R.; Abed, Y.; Rochette, L.; Bergeron, M.G.; Déry, P. Human metapneumovirus infections in hospitalized children. Emerg. Infect. Dis. 2003, 9, 634. [Google Scholar] [CrossRef]
- Peiris, J.M.; Tang, W.-H.; Chan, K.-H.; Khong, P.-L.; Guan, Y.; Lau, Y.-L.; Chiu, S.S. Children with Respiratory Disease Associated with Metapneumovirus in Hong Kong. Emerg. Infect. Dis. 2003, 9, 628–633. [Google Scholar] [CrossRef]
- McAdam, A.J.; Hasenbein, M.E.; Feldman, H.A.; Cole, S.E.; Offermann, J.T.; Riley, A.M.; Lieu, T.A. Human Metapneumovirus in Children Tested at a Tertiary-Care Hospital. J. Infect. Dis. 2004, 190, 20–26. [Google Scholar] [CrossRef]
- Leung, J.; Esper, F.; Weibel, C.; Kahn, J.S. Seroepidemiology of Human Metapneumovirus (hMPV) on the Basis of a Novel Enzyme-Linked Immunosorbent Assay Utilizing hMPV Fusion Protein Expressed in Recombinant Vesicular Stomatitis Virus. J. Clin. Microbiol. 2005, 43, 1213–1219. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.; Gonzalez, R.; Guo, L.; Wu, C.; Wu, J.; Vernet, G.; Paranhos-Baccalà, G.; Wang, J.; Hung, T. Large-scale seroprevalence analysis of human metapneumovirus and human respiratory syncytial virus infections in Beijing, China. Virol. J. 2011, 8, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Bastien, N.; Sidaway, F.; Chan, E.; Li, Y. Seroprevalence of human metapneumovirus (hMPV) in the Canadian province of Saskatchewan analyzed by a recombinant nucleocapsid protein-based enzyme-linked immunosorbent assay. J. Med. Virol. 2007, 79, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Biacchesi, S.; Skiadopoulos, M.H.; Boivin, G.; Hanson, C.T.; Murphy, B.R.; Collins, P.L.; Buchholz, U.J. Genetic diversity between human metapneumovirus subgroups. Virology 2003, 315, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hoogen, B.G.V.D.; Bestebroer, T.M.; Osterhaus, A.; Fouchier, R. Analysis of the Genomic Sequence of a Human Metapneumovirus. Virology 2002, 295, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.V.; Harris, P.A.; Tollefson, S.J.; Halburnt-Rush, L.L.; Pingsterhaus, J.M.; Edwards, K.M.; Wright, P.F.; Crowe, J.E. Human metapneumovirus and lower respiratory tract disease in otherwise healthy infants and children. N. Engl. J. Med. 2004, 350, 443–450. [Google Scholar] [CrossRef]
- Williams, J.V.; Wang, C.K.; Yang, C.; Tollefson, S.J.; House, F.S.; Heck, J.M.; Chu, M.; Brown, J.B.; Lintao, L.D.; Quinto, J.D.; et al. The role of human metapneumovirus in upper respiratory tract infections in children: A 20-year experience. J. Infect. Dis. 2005, 193, 387–395. [Google Scholar] [CrossRef]
- Williams, J.V.; Edwards, K.M.; Weinberg, G.A.; Griffin, M.R.; Hall, C.B.; Zhu, Y.; Szilagyi, P.G.; Wang, C.K.; Yang, C.; Silva, D.; et al. Population-based incidence of human metapneumovirus infection among hospitalized children. J. Infect. Dis. 2010, 201, 1890–1898. [Google Scholar] [CrossRef] [Green Version]
- Haas, L.E.M.; Thijsen, S.F.T.; Van Elden, L.J.R.; Heemstra, K.A. Human Metapneumovirus in Adults. Viruses 2013, 5, 87–110. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ren, L.; Guo, L.; Xiang, Z.; Paranhos-Baccalà, G.; Vernet, G.; Wang, J. Evolutionary Dynamics Analysis of Human Metapneumovirus Subtype A2: Genetic Evidence for Its Dominant Epidemic. PLoS ONE 2012, 7, e34544. [Google Scholar] [CrossRef] [Green Version]
- Schildgen, V.; Hoogen, B.V.D.; Fouchier, R.; Tripp, R.A.; Alvarez, R.; Manoha, C.; Williams, J.; Schildgen, O. Human Metapneumovirus: Lessons Learned over the First Decade. Clin. Microbiol. Rev. 2011, 24, 734–754. [Google Scholar] [CrossRef] [Green Version]
- Skiadopoulos, M.H.; Biacchesi, S.; Buchholz, U.J.; Riggs, J.M.; Surman, S.R.; Amaro-Carambot, E.; McAuliffe, J.M.; Elkins, W.R.; Claire, M.S.; Collins, P.L.; et al. The Two Major Human Metapneumovirus Genetic Lineages Are Highly Related Antigenically, and the Fusion (F) Protein Is a Major Contributor to This Antigenic Relatedness. J. Virol. 2004, 78, 6927–6937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoogen, B.G.V.D.; Herfst, S.; Sprong, L.; Cane, P.A.; Forleo-Neto, E.; De Swart, R.L.; Osterhaus, A.; Fouchier, R. Antigenic and Genetic Variability of Human Metapneumoviruses. Emerg. Infect. Dis. 2004, 10, 658–666. [Google Scholar] [CrossRef] [PubMed]
- MacPhail, M.; Schickli, J.H.; Tang, R.S.; Kaur, J.; Robinson, C.; Fouchier, R.A.M.; Osterhaus, A.; Spaete, R.R.; Haller, A.A. Identification of small-animal and primate models for evaluation of vaccine candidates for human metapneumovirus (hMPV) and implications for hMPV vaccine design. J. Gen. Virol. 2004, 85, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- Hoogen, B.V.D.; Herfst, S.; De Graaf, M.; Sprong, L.; Van Lavieren, R.; Van Amerongen, G.; Yüksel, S.; Fouchier, R.A.M.; Osterhaus, A.; De Swart, R.L. Experimental infection of macaques with human metapneumovirus induces transient protective immunity. J. Gen. Virol. 2007, 88, 1251–1259. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, G.; Déry, P.; Abed, Y.; Boivin, G. Respiratory Tract Reinfections by the New Human Metapneumovirus in an Immunocompromised Child. Emerg. Infect. Dis. 2002, 8, 976–978. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-F.; Wang, C.K.; Tollefson, S.J.; Piyaratna, R.; Lintao, L.D.; Chu, M.; Liem, A.; Mark, M.; Spaete, R.R.; Crowe, J.E.; et al. Genetic diversity and evolution of human metapneumovirus fusion protein over twenty years. Virol. J. 2009, 6, 138. [Google Scholar] [CrossRef] [Green Version]
- Peret, T.C.T.; Abed, Y.; Anderson, L.J.; Erdman, D.D.; Boivin, G. Sequence polymorphism of the predicted human metapneumovirus G glycoprotein. J. Gen. Virol. 2004, 85, 679–686. [Google Scholar] [CrossRef]
- Tang, R.S.; Mahmood, K.; MacPhail, M.; Guzzetta, J.M.; Haller, A.A.; Liu, H.; Kaur, J.; Lawlor, H.A.; Stillman, E.A.; Schickli, J.H.; et al. A host-range restricted parainfluenza virus type 3 (PIV3) expressing the human metapneumovirus (hMPV) fusion protein elicits protective immunity in African green monkeys. Vaccine 2005, 23, 1657–1667. [Google Scholar] [CrossRef]
- Skiadopoulos, M.H.; Biacchesi, S.; Buchholz, U.J.; Amaro-Carambot, E.; Surman, S.R.; Collins, P.L.; Murphy, B.R. Individual contributions of the human metapneumovirus F, G, and SH surface glycoproteins to the induction of neutralizing antibodies and protective immunity. Virology 2006, 345, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Ruigrok, R.W.H.; Crepin, T.; Kolakofsky, D. Nucleoproteins and nucleocapsids of negative-strand RNA viruses. Curr. Opin. Microbiol. 2011, 14, 504–510. [Google Scholar] [CrossRef]
- Rogers, M.; Williams, J.V. Quis Custodiet Ipsos Custodes? Regulation of Cell-Mediated Immune Responses Following Viral Lung Infections. Annu. Rev. Virol. 2018, 5, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Englund, J.A.; Boeckh, M.; Kuypers, J.; Nichols, W.G.; Hackman, R.C.; Morrow, R.A.; Fredricks, D.N.; Corey, L. Brief communication: Fatal human metapneumovirus infection in stem-cell transplant recipients. Ann. Intern. Med. 2006, 144, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Debiaggi, M.; Canducci, F.; Sampaolo, M.; Marinozzi, M.C.; Parea, M.; Terulla, C.; Colombo, A.; Alessandrino, E.P.; Bragotti, L.Z.; Arghittu, M.; et al. Persistent Symptomless Human Metapneumovirus Infection in Hematopoietic Stem Cell Transplant Recipients. J. Infect. Dis. 2006, 194, 474–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, J.A.; Gálvez, N.M.S.; Benavente, F.M.; Pizarro-Ortega, M.S.; Lay, M.K.; Riedel, C.; Bueno, S.M.; Gonzalez, P.A.; Kalergis, A.M. Human Metapneumovirus: Mechanisms and Molecular Targets Used by the Virus to Avoid the Immune System. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falsey, A.R.; Erdman, D.; Anderson, L.J.; Walsh, E.E. Human Metapneumovirus Infections in Young and Elderly Adults. J. Infect. Dis. 2003, 187, 785–790. [Google Scholar] [CrossRef]
- Pavlin, J.A.; Hickey, A.C.; Ulbrandt, N.; Chan, Y.-P.; Endy, T.P.; Boukhvalova, M.S.; Chunsuttiwat, S.; Nisalak, A.; Libraty, D.H.; Green, S.; et al. Human metapneumovirus reinfection among children in Thailand determined by ELISA using purified soluble fusion protein. J. Infect. Dis. 2008, 198, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Vargas, S.O.; Kozakewich, H.P.; Perez-Atayde, A.R.; McAdam, A.J. Pathology of Human Metapneumovirus Infection: Insights into the Pathogenesis of a Newly Identified Respiratory Virus. Pediatr. Dev. Pathol. 2004, 7, 478–486. [Google Scholar] [CrossRef]
- Kolli, D.; Gupta, M.R.; Sbrana, E.; Velayutham, T.S.; Chao, H.; Casola, A.; Garofalo, R.P. Alveolar Macrophages Contribute to the Pathogenesis of Human Metapneumovirus Infection while Protecting against Respiratory Syncytial Virus Infection. Am. J. Respir. Cell Mol. Boil. 2014, 51, 502–515. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.C.; Battini, L.; Tuyama, A.C.; Macip, S.; Melendi, G.A.; Horga, M.-A.; Gusella, G.L. Characterization of human metapneumovirus infection of myeloid dendritic cells. Virology 2007, 357, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Homaira, N.; Luby, S.; Petri, W.A.; Vainionpää, R.; Rahman, M.; Hossain, K.; Snider, C.B.; Rahman, M.; Alamgir, A.S.M.; Zesmin, F.; et al. Incidence of Respiratory Virus-Associated Pneumonia in Urban Poor Young Children of Dhaka, Bangladesh, 2009–2011. PLoS ONE 2012, 7, e32056. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Self, W.H.; Wunderink, R.G.; Fakhran, S.; Balk, R.; Bramley, A.M.; Reed, C.; Grijalva, C.G.; Anderson, E.J.; Courtney, D.M.; et al. Community-acquired pneumonia requiring hospitalization among U.S. adults. N. Engl. J. Med. 2015, 373, 415–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-F.; Tsao, K.-C.; Liu, Y.; Chen, Y.-C.; Yu, P.-C.; Huang, Y.-C.; Chou, C. Diagnosis of human metapneumovirus in patients hospitalized with acute lower respiratory tract infection using a metal-enhanced fluorescence technique. J. Virol. Methods 2015, 213, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Kuiken, T.; Hoogen, B.G.V.D.; Van Riel, D.; Laman, J.D.; Van Amerongen, G.; Sprong, L.; Fouchier, R.A.; Osterhaus, A. Experimental Human Metapneumovirus Infection of Cynomolgus Macaques (Macaca fascicularis) Results in Virus Replication in Ciliated Epithelial Cells and Pneumocytes with Associated Lesions throughout the Respiratory Tract. Am. J. Pathol. 2004, 164, 1893–1900. [Google Scholar] [CrossRef] [Green Version]
- Hamelin, M.-È.; Prince, G.A.; Gomez, A.M.; Kinkead, R.; Boivin, G. Human Metapneumovirus Infection Induces Long?Term Pulmonary Inflammation Associated with Airway Obstruction and Hyperresponsiveness in Mice. J. Infect. Dis. 2006, 193, 1634–1642. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.V.; Tollefson, S.J.; Johnson, J.E.; Crowe, J.E. The Cotton Rat (Sigmodon hispidus) Is a Permissive Small Animal Model of Human Metapneumovirus Infection, Pathogenesis, and Protective Immunity. J. Virol. 2005, 79, 10944–10951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.; Bao, X.; Liu, T.; Lai, S.; Li, K.; Garofalo, R.P.; Casola, A. Role of retinoic acid inducible gene-I in human metapneumovirus-induced cellular signalling. J. Gen. Virol. 2008, 89, 1978–1986. [Google Scholar] [CrossRef]
- Bao, X.; Liu, T.; Spetch, L.; Kolli, D.; Garofalo, R.; Casola, A. Airway epithelial cell response to human metapneumovirus infection. Virology 2007, 368, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Sinha, M.; Liu, T.; Hong, C.; Luxon, B.; Garofalo, R.; Casola, A. Identification of human metapneumovirus-induced gene networks in airway epithelial cells by microarray analysis. Virology 2008, 374, 114–127. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.J.; Takeuchi, O.; Akira, S. IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat. Immunol. 2005, 6, 981–988. [Google Scholar] [CrossRef]
- Seth, R.B.; Sun, L.; Ea, C.-K.; Chen, Z.J. Identification and Characterization of MAVS, a Mitochondrial Antiviral Signaling Protein that Activates NF-κB and IRF3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-stimulated genes: A complex web of host defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baños-Lara, M.D.R.; Piao, B.; Guerrero-Plata, A. Differential Mucin Expression by Respiratory Syncytial Virus and Human Metapneumovirus Infection in Human Epithelial Cells. Mediat. Inflamm. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loevenich, S.; Malmo, J.; Liberg, A.M.; Sherstova, T.; Li, Y.; Rian, K.; Johnsen, I.B.; Anthonsen, M.W. Cell-Type-Specific Transcription of Innate Immune Regulators in response to HMPV Infection. Mediat. Inflamm. 2019, 2019, 4964239–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoggins, J.W.; MacDuff, N.A.; Imanaka, N.; Gainey, M.D.; Shrestha, B.; Eitson, J.L.; Mar, K.B.; Richardson, R.B.; Ratushny, A.V.; Litvak, V.; et al. Pan-viral specificity of IFN-induced genes reveals new roles for cGAS in innate immunity. Nature 2013, 505, 691–695. [Google Scholar] [CrossRef] [PubMed]
- McMichael, T.M.; Zhang, Y.; Kenney, A.; Zhang, L.; Zani, A.; Lu, M.; Chemudupati, M.; Li, J.; Yount, J.S. IFITM3 Restricts Human Metapneumovirus Infection. J. Infect. Dis. 2018. [Google Scholar] [CrossRef] [Green Version]
- Weber, F.; Haller, O. Viral suppression of the interferon system. Biochimie 2007, 89, 836–842. [Google Scholar] [CrossRef]
- Dinwiddie, D.L.; Harrod, K. Human Metapneumovirus Inhibits IFN-α Signaling through Inhibition of STAT1 Phosphorylation. Am. J. Respir. Cell Mol. Boil. 2008, 38, 661–670. [Google Scholar] [CrossRef]
- Ren, J.; Kolli, D.; Liu, T.; Xu, R.; Garofalo, R.P.; Casola, A.; Bao, X. Human Metapneumovirus Inhibits IFN-β Signaling by Downregulating Jak1 and Tyk2 Cellular Levels. PLoS ONE 2011, 6, e24496. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Liu, T.; Shan, Y.; Li, K.; Garofalo, R.P.; Casola, A. Human Metapneumovirus Glycoprotein G Inhibits Innate Immune Responses. PLoS Pathog. 2008, 4, e1000077. [Google Scholar] [CrossRef]
- Uche, I.K.; Guerrero-Plata, A. Interferon-Mediated Response to Human Metapneumovirus Infection. Viruses 2018, 10, 505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baggiolini, M.; Dewald, B.; Moser, B. Human Chemokines: An Update. Annu. Rev. Immunol. 1997, 15, 675–705. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte Chemoattractant Protein-1 (MCP-1): An Overview. J. Interf. Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Futosi, K.; Fodor, S.; Mócsai, A. Neutrophil cell surface receptors and their intracellular signal transduction pathways. Int. Immunopharmacol. 2013, 17, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Hamelin, M.-E.; Yim, K.; Kuhn, K.H.; Cragin, R.P.; Boukhvalova, M.; Blanco, J.C.G.; Prince, G.A.; Boivin, G. Pathogenesis of Human Metapneumovirus Lung Infection in BALB/c Mice and Cotton Rats. J. Virol. 2005, 79, 8894–8903. [Google Scholar] [CrossRef] [Green Version]
- Lay, M.K.; Céspedes, P.F.; Palavecino, C.E.; León, M.A.; Díaz, R.A.; Salazar, F.J.; Méndez, G.P.; Bueno, S.M.; Kalergis, A.M. Human metapneumovirus infection activates the TSLP pathway that drives excessive pulmonary inflammation and viral replication in mice. Eur. J. Immunol. 2015, 45, 1680–1695. [Google Scholar] [CrossRef]
- Inngjerdingen, M.; Damaj, B.; Maghazachi, A.A.; Maghazachi, A. Human NK cells express CC chemokine receptors 4 and 8 and respond to thymus and activation-regulated chemokine, macrophage-derived chemokine, and I-309. J. Immunol. 2000, 164, 4048–4054. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-J. Thymic stromal lymphopoietin: Master switch for allergic inflammation. J. Exp. Med. 2006, 203, 269–273. [Google Scholar] [CrossRef]
- Li, Y.; Lund, C.; Nervik, I.; Loevenich, S.; Døllner, H.; Anthonsen, M.W.; Johnsen, I.B. Characterization of signaling pathways regulating the expression of pro-inflammatory long form thymic stromal lymphopoietin upon human metapneumovirus infection. Sci. Rep. 2018, 8, 883. [Google Scholar] [CrossRef] [Green Version]
- De Graaf, M.; Herfst, S.; Aarbiou, J.; Burgers, P.C.; Zaaraoui-Boutahar, F.; Bijl, M.; Van Ijcken, W.F.J.; Schrauwen, E.J.A.; Osterhaus, A.; Luider, T.M.; et al. Small Hydrophobic Protein of Human Metapneumovirus Does Not Affect Virus Replication and Host Gene Expression In Vitro. PLoS ONE 2013, 8, e58572. [Google Scholar] [CrossRef] [Green Version]
- De Lamballerie, C.N.; Pizzorno, A.; Dubois, J.; Julien, T.; Padey, B.; Bouveret, M.; Traversier, A.; Legras-Lachuer, C.; Lina, B.; Boivin, G.; et al. Characterization of cellular transcriptomic signatures induced by different respiratory viruses in human reconstituted airway epithelia. Sci. Rep. 2019, 9, 11493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Chahar, H.S.; Komaravelli, N.; Dossumekova, A.; Casola, A. Human metapneumovirus infection of airway epithelial cells is associated with changes in core metabolic pathways. Virology 2019, 531, 183–191. [Google Scholar] [CrossRef]
- Sanchez, E.L.; Lagunoff, M. Viral activation of cellular metabolism. Virology 2015, 479, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosakote, Y.M.; Jantzi, P.D.; Esham, D.L.; Spratt, H.; Kurosky, A.; Casola, A.; Garofalo, R.P. Viral-mediated Inhibition of Antioxidant Enzymes Contributes to the Pathogenesis of Severe Respiratory Syncytial Virus Bronchiolitis. Am. J. Respir. Crit. Care Med. 2011, 183, 1550–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelton, P.; Jaiswal, A.K. The transcription factor NF-E2-related Factor 2 (Nrf2): A protooncogene? FASEB J. 2012, 27, 414–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanciuc, T.; Sbrana, E.; Casola, A.; Garofalo, R.P. Protective Role of Nuclear Factor Erythroid 2-Related Factor 2 Against Respiratory Syncytial Virus and Human Metapneumovirus Infections. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Hay, S.; Kannourakis, G. A time to kill: Viral manipulation of the cell death program. J. Gen. Virol. 2002, 83, 1547–1564. [Google Scholar] [CrossRef]
- Hilleman, M.R. Strategies and mechanisms for host and pathogen survival in acute and persistent viral infections. Proc. Natl. Acad. Sci. USA 2004, 101, 14560–14566. [Google Scholar] [CrossRef] [Green Version]
- Marsico, S.; Caccuri, F.; Mazzuca, P.; Apostoli, P.; Roversi, S.; Lorenzin, G.; Zani, A.; Fiorentini, S.; Giagulli, C.; Caruso, A. Human lung epithelial cells support human metapneumovirus persistence by overcoming apoptosis. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef] [Green Version]
- Young, L.S.; Dawson, C.W.; Eliopoulos, A.G. Viruses and apoptosis. Br. Med. Bull. 1997, 53, 509–521. [Google Scholar] [CrossRef]
- Alvarez, R.; Harrod, K.; Shieh, W.-J.; Zaki, S.; Tripp, R.A. Human Metapneumovirus Persists in BALB/c Mice despite the Presence of Neutralizing Antibodies. J. Virol. 2004, 78, 14003–14011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, R.; Tripp, R.A. The Immune Response to Human Metapneumovirus Is Associated with Aberrant Immunity and Impaired Virus Clearance in BALB/c Mice. J. Virol. 2005, 79, 5971–5978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Haas, D.L.; Poore, S.; Isakovic, S.; Gahan, M.E.; Mahalingam, S.; Fu, Z.F.; Tripp, R.A. Human Metapneumovirus Establishes Persistent Infection in the Lungs of Mice and Is Reactivated by Glucocorticoid Treatment. J. Virol. 2009, 83, 6837–6848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abed, Y.; Boivin, G. Persistent Human Metapneumovirus Infection in Immunocompromised Child. Emerg. Infect. Dis. 2008, 14, 854–856. [Google Scholar] [CrossRef]
- Newton, A.H.; Cardani, A.; Braciale, T.J. The host immune response in respiratory virus infection: Balancing virus clearance and immunopathology. Semin. Immunopathol. 2016, 38, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Pribul, P.K.; Harker, J.A.; Wang, B.; Wang, H.; Tregoning, J.; Schwarze, J.; Openshaw, P.J. Alveolar Macrophages Are a Major Determinant of Early Responses to Viral Lung Infection but Do Not Influence Subsequent Disease Development. J. Virol. 2008, 82, 4441–4448. [Google Scholar] [CrossRef] [Green Version]
- Benoit, A.; Huang, Y.; Proctor, J.; Rowden, G.; Anderson, R. Effects of alveolar macrophage depletion on liposomal vaccine protection against respiratory syncytial virus (RSV). Clin. Exp. Immunol. 2006, 145, 147–154. [Google Scholar] [CrossRef]
- Li, Y.; Østerhus, S.; Johnsen, I.B. Human Metapneumovirus Infection Inhibits Cathelicidin Antimicrobial Peptide Expression in Human Macrophages. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Cheemarla, N.; Baños-Lara, M.D.R.; Naidu, S.; Guerrero-Plata, A. Neutrophils regulate the lung inflammatory response via γδ T cell infiltration in an experimental mouse model of human metapneumovirus infection. J. Leukoc. Boil. 2017, 101, 1383–1392. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.C.; Tollefson, S.J.; Johnson, M.; Gilchuk, P.; Boyd, K.L.; Shepherd, B.; Joyce, S.; Williams, J.V. Acute Clearance of Human Metapneumovirus Occurs Independently of Natural Killer Cells. J. Virol. 2014, 88, 10963–10969. [Google Scholar] [CrossRef] [Green Version]
- Kolli, D.; Bataki, E.L.; Spetch, L.; Guerrero-Plata, A.; Jewell, A.M.; Piedra, P.A.; Milligan, G.N.; Garofalo, R.P.; Casola, A. T Lymphocytes Contribute to Antiviral Immunity and Pathogenesis in Experimental Human Metapneumovirus Infection. J. Virol. 2008, 82, 8560–8569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, M.; Lamens, K.; Shafagati, N.; Johnson, M.; Oury, T.D.; Joyce, S.; Williams, J.V. CD4+ Regulatory T Cells Exert Differential Functions during Early and Late Stages of the Immune Response to Respiratory Viruses. J. Immunol. 2018, 201, 1253–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melendi, G.A.; Zavala, F.; Buchholz, U.J.; Boivin, G.; Collins, P.L.; Kleeberger, S.R.; Polack, F.P. Mapping and Characterization of the Primary and Anamnestic H-2d-Restricted Cytotoxic T-Lymphocyte Response in Mice against Human Metapneumovirus. J. Virol. 2007, 81, 11461–11467. [Google Scholar] [CrossRef] [Green Version]
- Culshaw, S.; Millington, O.R.; Brewer, J.M.; McInnes, I. Murine neutrophils present Class II restricted antigen. Immunol. Lett. 2008, 118, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulendran, B.; Maddur, M.S. Innate Immune Sensing and Response to Influenza. Curr. Top. Microbiol. Immunol. 2014, 386, 23–71. [Google Scholar] [CrossRef]
- Cheemarla, N.; Guerrero-Plata, A. Immune Response to Human Metapneumovirus Infection: What We Have Learned from the Mouse Model. Pathogens 2015, 4, 682–696. [Google Scholar] [CrossRef] [Green Version]
- Cavallaro, E.; Liang, K.; Lawrence, M.D.; Forsyth, K.D.; Dixon, D.-L. Neutrophil infiltration and activation in bronchiolitic airways are independent of viral etiology. Pediatr. Pulmonol. 2016, 52, 238–246. [Google Scholar] [CrossRef]
- Daley, J.M.; Thomay, A.A.; Connolly, M.D.; Reichner, J.; Albina, J. Use of Ly6G-specific monoclonal antibody to deplete neutrophils in mice. J. Leukoc. Boil. 2007, 83, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Cortjens, B.; Ingelse, S.A.; Calis, J.C.J.; Vlaar, A.P.; Koenderman, L.; Bem, R.A.; Van Woensel, J.B. Neutrophil subset responses in infants with severe viral respiratory infection. Clin. Immunol. 2017, 176, 100–106. [Google Scholar] [CrossRef]
- Pillay, J.; Kamp, V.M.; Van Hoffen, E.; Visser, T.; Tak, T.; Lammers, J.-W.; Ulfman, L.H.; Leenen, L.P.; Pickkers, P.; Koenderman, L. A subset of neutrophils in human systemic inflammation inhibits T cell responses through Mac-1. J. Clin. Investig. 2011, 122, 327–336. [Google Scholar] [CrossRef]
- Zilio, S.; Serafini, P. Neutrophils and Granulocytic MDSC: The Janus God of Cancer Immunotherapy. Vaccines 2016, 4, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheemarla, N.; Guerrero-Plata, A. Human Metapneumovirus Attachment Protein Contributes to Neutrophil Recruitment into the Airways of Infected Mice. Viruses 2017, 9, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baños-Lara, M.D.R.; Harvey, L.; Mendoza, A.; Simms, D.; Chouljenko, V.N.; Wakamatsu, N.; Kousoulas, K.G.; Guerrero-Plata, A. Impact and Regulation of Lambda Interferon Response in Human Metapneumovirus Infection. J. Virol. 2014, 89, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Plata, A. Dendritic Cells in Human Pneumovirus and Metapneumovirus Infections. Viruses 2013, 5, 1553–1570. [Google Scholar] [CrossRef] [PubMed]
- Neyt, K.; Lambrecht, B.N. The role of lung dendritic cell subsets in immunity to respiratory viruses. Immunol. Rev. 2013, 255, 57–67. [Google Scholar] [CrossRef]
- Guerrero-Plata, A.; Kolli, D.; Hong, C.; Casola, A.; Garofalo, R.P. Subversion of pulmonary dendritic cell function by paramyxovirus infections. J. Immunol. 2009, 182, 3072–3083. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Plata, A.; Casola, A.; Suárez, G.; Yu, X.; Spetch, L.; Peeples, M.E.; Garofalo, R.P. Differential Response of Dendritic Cells to Human Metapneumovirus and Respiratory Syncytial Virus. Am. J. Respir. Cell Mol. Boil. 2006, 34, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Kolli, D.; Esham, D.; Velayutham, T.S.; Casola, A. Human Metapneumovirus Small Hydrophobic Protein Inhibits Interferon Induction in Plasmacytoid Dendritic Cells. Viruses 2018, 10, 278. [Google Scholar] [CrossRef] [Green Version]
- Goutagny, N.; Jiang, Z.; Tian, J.; Parroche, P.; Schickli, J.; Monks, B.G.; Ulbrandt, N.; Ji, H.; Kiener, P.A.; Coyle, A.J.; et al. Cell type-specific recognition of human metapneumoviruses (HMPVs) by retinoic acid-inducible gene I (RIG-I) and TLR7 and viral interference of RIG-I ligand recognition by HMPV-B1 phosphoprotein. J. Immunol. 2009, 184, 1168–1179. [Google Scholar] [CrossRef] [Green Version]
- Smyth, L.A.; Boardman, D.; Tung, S.L.; Lechler, R.; Lombardi, G. MicroRNAs affect dendritic cell function and phenotype. Immunology 2015, 144, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Banos-Lara, M.; Zabaleta, J.; Garai, J.; Baddoo, M.; Guerrero-Plata, A. Comparative analysis of miRNA profile in human dendritic cells infected with respiratory syncytial virus and human metapneumovirus. BMC Res. Notes 2018, 11, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Nouën, C.; Hillyer, P.; Munir, S.; Winter, C.C.; Mccarty, T.; Bukreyev, A.; Collins, P.L.; Rabin, R.L.; Buchholz, U.J. Effects of Human Respiratory Syncytial Virus, Metapneumovirus, Parainfluenza Virus 3 and Influenza Virus on CD4+ T Cell Activation by Dendritic Cells. PLoS ONE 2010, 5, e15017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Céspedes, P.F.; Gonzalez, P.A.; Kalergis, A.M. Human metapneumovirus keeps dendritic cells from priming antigen-specific naive T cells. Immunology 2013, 139, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Hoogen, B.V.D.; Van Boheemen, S.; De Rijck, J.; Van Nieuwkoop, S.; Smith, D.J.; Laksono, B.; Gultyaev, A.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Excessive production and extreme editing of human metapneumovirus defective interfering RNA is associated with type I IFN induction. J. Gen. Virol. 2014, 95, 1625–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, A.S. Defective Interfering Viruses. Annu. Rev. Microbiol. 1973, 27, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Biacchesi, S.; Skiadopoulos, M.H.; Yang, L.; Lamirande, E.W.; Tran, K.C.; Murphy, B.R.; Collins, P.L.; Buchholz, U.J. Recombinant Human Metapneumovirus Lacking the Small Hydrophobic SH and/or Attachment G Glycoprotein: Deletion of G Yields a Promising Vaccine Candidate. J. Virol. 2004, 78, 12877–12887. [Google Scholar] [CrossRef] [Green Version]
- Biacchesi, S.; Skiadopoulos, M.H.; Tran, K.C.; Murphy, B.R.; Collins, P.L.; Buchholz, U.J. Recovery of human metapneumovirus from cDNA: Optimization of growth in vitro and expression of additional genes. Virology 2004, 321, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Le, V.B.; Dubois, J.; Couture, C.; Cavanagh, M.H.; Uyar, O.; Pizzorno, A.; Rosa-Calatrava, M.; Hamelin, M.E.; Boivin, G. Human metapneumovirus activates NOD-like receptor protein 3 inflammasome via its smallhydrophobic protein which plays adetrimental role during infection in mice. PLoS Pathog. 2019, 15, e1007689. [Google Scholar] [CrossRef]
- Kolli, D.; Bao, X.; Liu, T.; Hong, C.; Wang, T.; Garofalo, R.P.; Casola, A. Human metapneumovirus glycoprotein G inhibits TLR4-dependent signaling in monocyte-derived dendritic cells. J. Immunol. 2011, 187, 47–54. [Google Scholar] [CrossRef]
- Preston, F.M.; Straub, C.P.; Ramirez, R.; Mahalingam, S.; Spann, K. siRNA against the G gene of human metapneumovirus. Virol. J. 2012, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Dossumbekova, A.; Kolli, D.; Esham, D.L.; Liu, T.; Casola, A. Human Metapneumovirus Small Hydrophobic Protein Inhibits Interferon Induction in Plasmacytoid Dendritic Cells via TLR7 Signaling Pathway. J. Allergy Clin. Immunol. 2017, 139, AB16. [Google Scholar] [CrossRef]
- Hastings, A.K.; Amato, K.R.; Wen, S.C.; Peterson, L.S.; Williams, J.V. Human metapneumovirus small hydrophobic (SH) protein downregulates type I IFN pathway signaling by affecting STAT1 expression and phosphorylation. Virology 2016, 494, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Biacchesi, S.; Pham, Q.N.; Skiadopoulos, M.H.; Murphy, B.R.; Collins, P.L.; Buchholz, U.J. Infection of Nonhuman Primates with Recombinant Human Metapneumovirus Lacking the SH, G, or M2-2 Protein Categorizes Each as a Nonessential Accessory Protein and Identifies Vaccine Candidates. J. Virol. 2005, 79, 12608–12613. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Liu, G.; Go, J.; Kolli, D.; Zhang, G.; Bao, X. Human Metapneumovirus M2-2 Protein Inhibits Innate Immune Response in Monocyte-Derived Dendritic Cells. PLoS ONE 2014, 9, e91865. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, Y.; Sakai, M.; Funayama, M.; Itoh, M.; Gotoh, B. Human Metapneumovirus M2-2 Protein Acts as a Negative Regulator of Alpha Interferon Production by Plasmacytoid Dendritic Cells. J. Virol. 2017, 91, e00579-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Li, C.; Feng, L.; Pan, W.; Li, L.; Wang, Q.; Li, J.; Li, N.; Han, L.; Zheng, X.; et al. Proton Channel Activity of Influenza A Virus Matrix Protein 2 Contributes to Autophagy Arrest. J. Virol. 2015, 90, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Choi, E.; Zhang, K.; Chen, Y.; Ye, S.; Deng, X.; Zhang, K.; Bao, X. Detection of Nuclear Protein Profile Changes by Human Metapneumovirus M2-2 Protein Using Quantitative Differential Proteomics. Vaccines 2017, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.; Wu, W.; Chen, Y.; Yan, W.; Li, L.; Choudhury, A.; Bao, X. The role of M2-2 PDZ-binding motifs in pulmonary innate immune responses to human metapneumovirus. J. Med. Virol. 2020, 2. [Google Scholar] [CrossRef]
- Obeidy, P.; Sharland, A.F. NKG2D and its ligands. Int. J. Biochem. Cell Boil. 2009, 41, 2364–2367. [Google Scholar] [CrossRef]
- Stern-Ginossar, N.; Mandelboim, O. An integrated view of the regulation of NKG2D ligands. Immunology 2009, 128, 1–6. [Google Scholar] [CrossRef]
- Uzhachenko, R.V.; Shanker, A. CD8+ T Lymphocyte and NK Cell Network: Circuitry in the Cytotoxic Domain of Immunity. Front. Immunol. 2019, 10, 1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huck, B.; Neumann-Haefelin, D.; Schmitt-Graeff, A.; Weckmann, M.; Mattes, J.; Ehl, S.; Falcone, V. Human metapneumovirus induces more severe disease and stronger innate immune response in BALB/c mice as compared with respiratory syncytial virus. Respir. Res. 2007, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaya, M.; Barral, P.; Burbage, M.; Aggarwal, S.; Montaner, B.; Navia, A.W.; Aid, M.; Tsui, C.; Maldonado, P.; Nair, U.; et al. Initiation of Antiviral B Cell Immunity Relies on Innate Signals from Spatially Positioned NKT Cells. Cell 2018, 172, 517–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kaer, L. Regulation of Immune Responses by CD1d-Restricted Natural Killer T Cells. Immunol. Res. 2004, 30, 139–154. [Google Scholar] [CrossRef]
- Diab, M.; Glasner, A.; Isaacson, B.; Bar-On, Y.; Drori, Y.; Yamin, R.; Duev-Cohen, A.; Danziger, O.; Zamostiano, R.; Mandelboim, M.; et al. NK-cell receptors NKp46 and NCR1 control human metapneumovirus infection. Eur. J. Immunol. 2017, 47, 692–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, S.; Paget, C.; Trottein, F. Role of Non-conventional T Lymphocytes in Respiratory Infections: The Case of the Pneumococcus. PLoS Pathog. 2014, 10, e1004300. [Google Scholar] [CrossRef] [PubMed]
- Herd, K.A.; Nissen, M.; Hopkins, P.M.; Sloots, T.P.; Tindle, R.W. Major Histocompatibility Complex Class I Cytotoxic T Lymphocyte Immunity to Human Metapneumovirus (hMPV) in Individuals with Previous hMPV Infection and Respiratory Disease. J. Infect. Dis. 2008, 197, 584–592. [Google Scholar] [CrossRef]
- Laham, F.R.; Israele, V.; Casellas, J.M.; Garcia, A.M.; Prugent, C.M.L.; Hoffman, S.J.; Hauer, D.; Thumar, B.; Name, M.I.; Pascual, A.; et al. Differential Production of Inflammatory Cytokines in Primary Infection with Human Metapneumovirus and with Other Common Respiratory Viruses of Infancy. J. Infect. Dis. 2004, 189, 2047–2056. [Google Scholar] [CrossRef]
- Douville, R.N.; Bastien, N.; Li, Y.; Pochard, P.; Simons, F.E.R.; HayGlass, K.T. Human Metapneumovirus Elicits Weak IFN-γ Memory Responses Compared with Respiratory Syncytial Virus. J. Immunol. 2006, 176, 5848–5855. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, S.; Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T. Regulatory T cells: How do they suppress immune responses? Int. Immunol. 2009, 21, 1105–1111. [Google Scholar] [CrossRef]
- Liu, Y.; Qin, T.; Zhao, X.; Dong, S.; Zhu, J.; Peng, D.; Zhong, J.; Li, T.; Chen, X. Skewed balance of regulatory T cell and inflammatory T cell in IL-17 defect with human metapneumovirus infection. Cell. Immunol. 2018, 331, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Tzannou, I.; Nicholas, S.K.; Lulla, P.; Aguayo-Hiraldo, P.I.; Misra, A.; Martinez, C.A.; Machado, A.; Orange, J.S.; Piedra, P.A.; Vera, J.F.; et al. Immunologic Profiling of Human Metapneumovirus for the Development of Targeted Immunotherapy. J. Infect. Dis. 2017, 216, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, J.; Gilchuk, P.; Hastings, A.K.; Tollefson, S.J.; Johnson, M.; Downing, M.B.; Boyd, K.L.; Johnson, J.E.; Kim, A.S.; Joyce, S.; et al. Viral acute lower respiratory infections impair CD8+ T cells through PD-1. J. Clin. Investig. 2012, 122, 2967–2982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, J.; Rogers, M.; Hastings, A.K.; Tollefson, S.J.; Williams, J.V. Programmed death-1 impairs secondary effector lung CD8⁺ T cells during respiratory virus reinfection. J. Immunol. 2014, 193, 5108–5117. [Google Scholar] [CrossRef] [Green Version]
- Hastings, A.K.; Erickson, J.J.; Schuster, J.E.; Boyd, K.L.; Tollefson, S.J.; Johnson, M.; Gilchuk, P.; Joyce, S.; Williams, J.V. Role of Type I Interferon Signaling in Human Metapneumovirus Pathogenesis and Control of Viral Replication. J. Virol. 2015, 89, 4405–4420. [Google Scholar] [CrossRef] [Green Version]
- Falsey, A.R.; Hennessey, P.A.; Formica, M.A.; Criddle, M.M.; Biear, J.M.; Walsh, E.E. Humoral immunity to human metapneumovirus infection in adults. Vaccine 2009, 28, 1477–1480. [Google Scholar] [CrossRef]
- El Najjar, F.; Cifuentes-Muñoz, N.; Chen, J.; Zhu, H.; Buchholz, U.J.; Moncman, C.L.; Dutch, R.E. Human metapneumovirus Induces Reorganization of the Actin Cytoskeleton for Direct Cell-to-Cell Spread. PLoS Pathog. 2016, 12, e1005922. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Qian, X.; Lattmann, S.; El Sahili, A.; Yeo, T.H.; Jia, H.; Cressey, T.; Ludeke, B.; Noton, S.; Kalocsay, M.; et al. Structure of the human metapneumovirus polymerase phosphoprotein complex. Nature 2019, 577, 275–279. [Google Scholar] [CrossRef]
- Langley, J.M.; Macdonald, L.D.; Weir, G.M.; MacKinnon-Cameron, D.; Ye, L.; McNeil, S.A.; Schepens, B.; Saelens, X.; Stanford, M.M.; Halperin, S.A. A Respiratory Syncytial Virus Vaccine Based on the Small Hydrophobic Protein Ectodomain Presented with a Novel Lipid-Based Formulation Is Highly Immunogenic and Safe in Adults: A First-in-Humans Study. J. Infect. Dis. 2018, 218, 378–387. [Google Scholar] [CrossRef]
- Chlibek, R.; Smetana, J.; Pauksens, K.; Rombo, L.; Hoek, J.A.R.V.D.; Richardus, J.H.; Plassmann, G.; Schwarz, T.F.; Ledent, E.; Heineman, T.C. Safety and immunogenicity of three different formulations of an adjuvanted varicella-zoster virus subunit candidate vaccine in older adults: A phase II, randomized, controlled study. Vaccine 2014, 32, 1745–1753. [Google Scholar] [CrossRef]
- Rogers, M.C.; Williams, J.V. Reining in the CD8+ T cell: Respiratory virus infection and PD-1-mediated T-cell impairment. PLoS Pathog. 2019, 15, e1007387. [Google Scholar] [CrossRef] [PubMed] [Green Version]



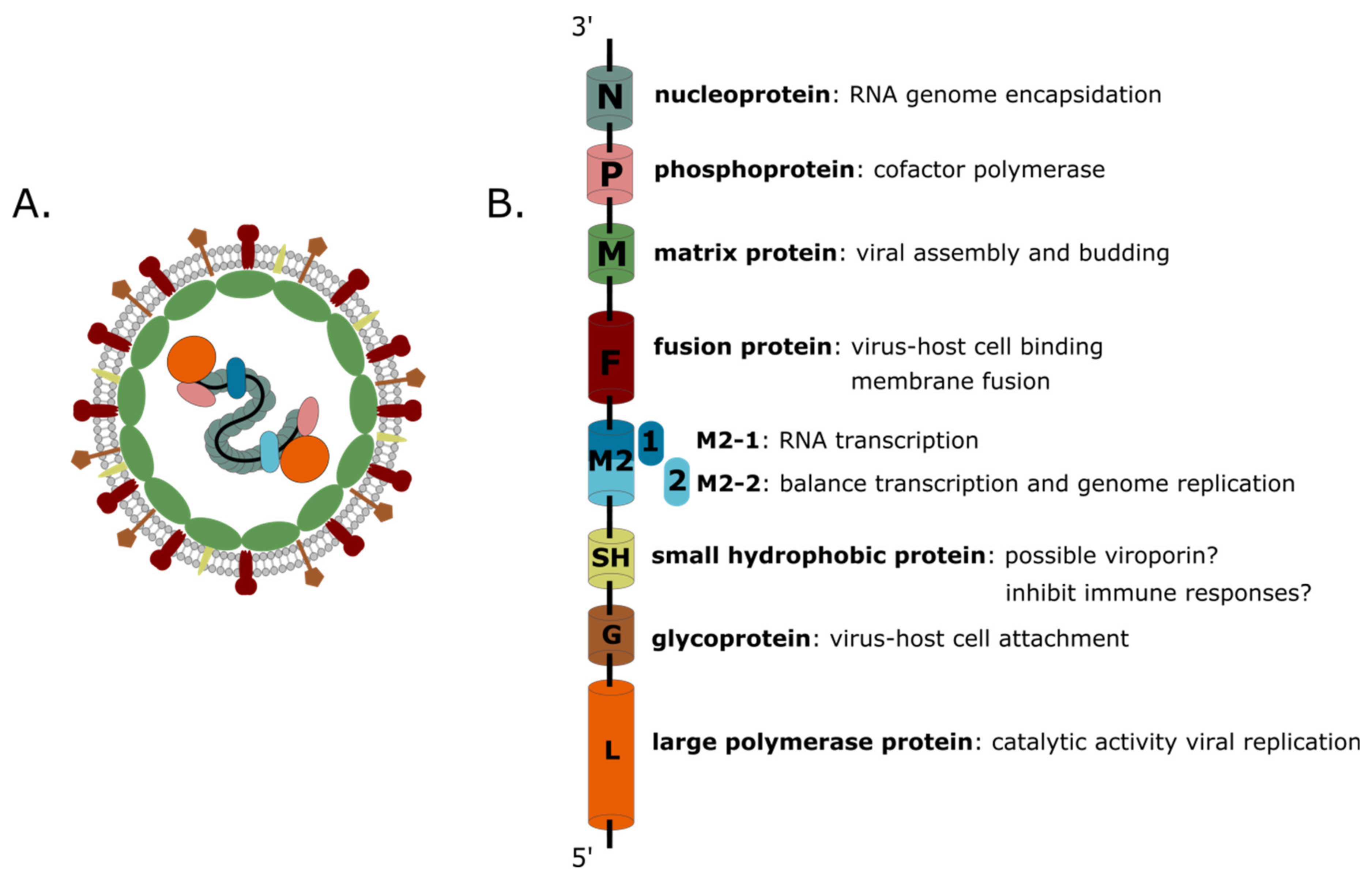{kind=link}
{kind=link}
| Cell Type | Strategy for Cell Type Depletion | Time of Depletion | Lung Viral Load | Lung Inflammation and Histopathology | Mouse Strain | Reference |
|---|---|---|---|---|---|---|
| Innate Immunity | ||||||
| AMφ | L-CL2MBP liposomes | Before hMPV inoculation | Decreased | Decreased | BALB/c | [48] |
| After hMPV inoculation | Unchanged | Not investigated | BALB/c | [48] | ||
| Neutrophils | anti-Ly6G monoclonal antibody | Before hMPV inoculation | Unchanged Decreased | Increased Decreased | BALB/c BALB/c | [76] [99] |
| NK Cells | Anti CD49b/Pan NK cell antibody | Before hMPV inoculation | Increased | Not investigated | BALB/c | [91] |
| Anti NK1.1 antibody | Before hMPV inoculation | Unchanged | Unchanged | C57BL/6 | [100] | |
| NKT Cells | CD1d-/- mice | Before hMPV inoculation | Unchanged | Unchanged | C57BL/6 | [100] |
| Adaptive Immunity | ||||||
| T Cells | Anti CD4 and anti CD8 antibody | Before hMPV inoculation | Increased | Decreased | BALB/c | [101] |
| Anti CD3ε + anti αβTCR antibody | Before hMPV inoculation | Increased | Not investigated | BALB/c | [91] | |
| CD4+ T Cells | Anti CD4 antibody | Before hMPV inoculation | Unchanged | Decreased | BALB/c | [101] |
| Tregs | FoxP3DTR mice | Throughout hMPV infection | Increased | Increased | C57BL/6 | [102] |
| Before hMPV inoculation (early) | Increased | Increased | C57BL/6 | [102] | ||
| After hMPV inoculation (late) | Unchanged | Increased | C57BL/6 | [102] | ||
| Anti CD25 antibody | Before hMPV inoculation | Decreased | Not investigated | C57BL/6 | [102] | |
| CD8+ T Cells | Anti CD8 antibody | Before hMPV inoculation | Unchanged | Decreased | BALB/c | [101] |
| Adoptive transfer hMPV specific CTLs in Rag1−/− mice | Before hMPV inoculation | Decreased | Not investigated | BALB/c | [103] | |
| γδ T Cells | TCR-δ KO mice (B6.129P2-Tcrdtm1Mom/J) | Before hMPV inoculation | Not investigated | Decreased | C57BL/6 | [99] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballegeer, M.; Saelens, X. Cell-Mediated Responses to Human Metapneumovirus Infection. Viruses 2020, 12, 542. https://doi.org/10.3390/v12050542
Ballegeer M, Saelens X. Cell-Mediated Responses to Human Metapneumovirus Infection. Viruses. 2020; 12(5):542. https://doi.org/10.3390/v12050542
Chicago/Turabian StyleBallegeer, Marlies, and Xavier Saelens. 2020. "Cell-Mediated Responses to Human Metapneumovirus Infection" Viruses 12, no. 5: 542. https://doi.org/10.3390/v12050542
APA StyleBallegeer, M., & Saelens, X. (2020). Cell-Mediated Responses to Human Metapneumovirus Infection. Viruses, 12(5), 542. https://doi.org/10.3390/v12050542