Pesticide–Virus Interactions in Honey Bees: Challenges and Opportunities for Understanding Drivers of Bee Declines


{kind=link}
Abstract
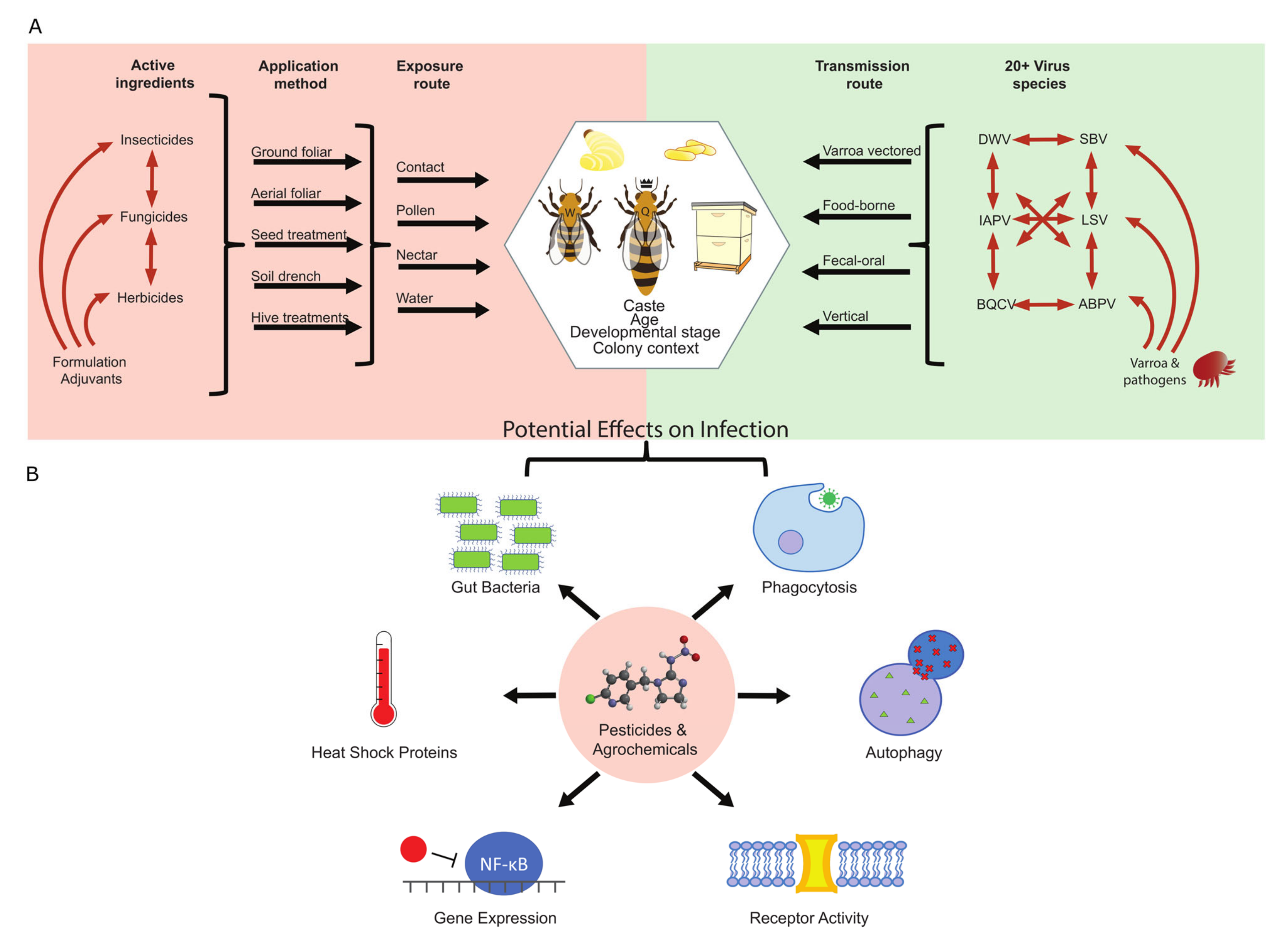
:1. Introduction
2. Pesticides
2.1. General Background on Classes of Pesticides
2.2. How Bees Detoxify Pesticides
2.3. Active Ingredients vs. Formulations
2.4. Treating for Varroa mites
3. Honey Bee Viruses
3.1. Sublethal Infection
3.2. Viral Transmission
3.3. Antiviral Immune Pathways
4. Interaction of Pesticides and Honey Bee Viruses
4.1. How Pesticides Can Impact Antiviral Pathways
4.2. Laboratory- vs. Field-Based Studies
5. Future Directions
Funding
Acknowledgments
Conflicts of Interest
References
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic Valuation of the Vulnerability of World Agriculture Confronted with Pollinator Decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- United States Department of Agriculture/National Agricultural Statistics Survey. Honey Bee Colonies; United States Department of Agriculture/National Agricultural Statistics Survey: Washington, DC, USA, 2019. [Google Scholar]
- Naug, D. Nutritional Stress Due to Habitat Loss May Explain Recent Honeybee Colony Collapses. Biol. Conserv. 2009, 142, 2369–2372. [Google Scholar] [CrossRef]
- Paudel, Y.P.; Mackereth, R.; Hanley, R.; Qin, W. Honey Bees (Apis Mellifera L.) and Pollination Issues: Current Status, Impacts, and Potential Drivers of Decline. J. Agric. Sci. 2015, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.-P.; van Engelsdorp, D. Drivers of Colony Losses. Curr. Opin. Insect Sci. 2018, 26, 142–148. [Google Scholar] [CrossRef]
- Smith, K.M.; Loh, E.H.; Rostal, M.K.; Zambrana-Torrelio, C.M.; Mendiola, L.; Daszak, P. Pathogens, Pests, and Economics: Drivers of Honey Bee Colony Declines and Losses. EcoHealth 2013, 10, 434–445. [Google Scholar] [CrossRef]
- McMenamin, A.J.; Genersch, E. Honey Bee Colony Losses and Associated Viruses. Curr. Opin. Insect Sci. 2015, 8, 121–129. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Nazzi, F.; Brown, S.P.; Annoscia, D.; Del Piccolo, F.; Di Prisco, G.; Varricchio, P.; Della Vedova, G.; Cattonaro, F.; Caprio, E.; Pennacchio, F. Synergistic Parasite-Pathogen Interactions Mediated by Host Immunity Can Drive the Collapse of Honeybee Colonies. PLoS Pathog. 2012, 8, e1002735. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.-Q.; Gong, H.-R.; Huang, S.-K.; Sohr, A.; Hu, F.-L.; Chen, Y.P. Evidence of the Synergistic Interaction of Honey Bee Pathogens Nosema Ceranae and Deformed Wing Virus. Vet. Microbiol. 2015, 177, 1–6. [Google Scholar] [CrossRef]
- Grassl, J.; Holt, S.; Cremen, N.; Peso, M.; Hahne, D.; Baer, B. Synergistic Effects of Pathogen and Pesticide Exposure on Honey Bee (Apis Mellifera) Survival and Immunity. J. Invertebr. Pathol. 2018, 159, 78–86. [Google Scholar] [CrossRef]
- Zhu, Y.C.; Yao, J.; Adamczyk, J.; Luttrell, R. Synergistic Toxicity and Physiological Impact of Imidacloprid Alone and Binary Mixtures with Seven Representative Pesticides on Honey Bee (Apis Mellifera). PLoS ONE 2017, 12, e0176837. [Google Scholar] [CrossRef] [PubMed]
- Aufauvre, J.; Biron, D.G.; Vidau, C.; Fontbonne, R.; Roudel, M.; Diogon, M.; Viguès, B.; Belzunces, L.P.; Delbac, F.; Blot, N. Parasite-Insecticide Interactions: A Case Study of Nosema Ceranae and Fipronil Synergy on Honeybee. Sci. Rep. 2012, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Retschnig, G.; Neumann, P.; Williams, G.R. Thiacloprid–Nosema Ceranae Interactions in Honey Bees: Host Survivorship but Not Parasite Reproduction Is Dependent on Pesticide Dose. J. Invertebr. Pathol. 2014, 118, 18–19. [Google Scholar] [CrossRef] [PubMed]
- Nazzi, F.; Pennacchio, F. Disentangling Multiple Interactions in the Hive Ecosystem. Trends Parasitol. 2014, 30, 556–561. [Google Scholar] [CrossRef] [PubMed]
- DeGrandi-Hoffman, G.; Chen, Y.; Huang, E.; Huang, M.H. The Effect of Diet on Protein Concentration, Hypopharyngeal Gland Development and Virus Load in Worker Honey Bees (Apis Mellifera L.). J. Insect Physiol. 2010, 56, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Dantec, C.; Parrinello, H.; Le Conte, Y. Nutrigenomics in Honey Bees: Digital Gene Expression Analysis of Pollen’s Nutritive Effects on Healthy and Varroa-Parasitized Bees. BMC Genomics 2011, 12, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of Pollen Nutrition on Honey Bee Health: Do Pollen Quality and Diversity Matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolezal, A.G.; Carrillo-Tripp, J.; Judd, T.M.; Allen Miller, W.; Bonning, B.C.; Toth, A.L. Interacting Stressors Matter: Diet Quality and Virus Infection in Honeybee Health. R. Soc. Open Sci. 2019, 6, 181803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.M.; Ellis, M.D.; Mullin, C.A.; Frazier, M. Pesticides and Honey Bee Toxicity–USA. Apidologie 2010, 41, 312–331. [Google Scholar] [CrossRef] [Green Version]
- Long, E.Y.; Krupke, C.H. Non-Cultivated Plants Present a Season-Long Route of Pesticide Exposure for Honey Bees. Nat. Commun. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Chauzat, M.-P.; Faucon, J.-P.; Martel, A.-C.; Lachaize, J.; Cougoule, N.; Aubert, M. A Survey of Pesticide Residues in Pollen Loads Collected by Honey Bees in France. J. Econ. Entomol. 2006, 99, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; Pettis, J.S. High Levels of Miticides and Agrochemicals in North American Apiaries: Implications for Honey Bee Health. PLoS ONE 2010, 5, e9754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beekman, M.; Ratnieks, F.L.W. Long-Range Foraging by the Honey-Bee, Apis mellifera L. Funct. Ecol. 2000, 14, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Tosi, S.; Nieh, J.C. Lethal and Sublethal Synergistic Effects of a New Systemic Pesticide, Flupyradifurone (Sivanto®), on Honeybees. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190433. [Google Scholar] [CrossRef] [Green Version]
- Mullin, C.A.; Chen, J.; Fine, J.D.; Frazier, M.T.; Frazier, J.L. The Formulation Makes the Honey Bee Poison. Pestic. Biochem. Physiol. 2015, 120, 27–35. [Google Scholar] [CrossRef]
- US Environmental Protection Agency. Guidance on Exposure and Efffects Testing for Assessing Risks to Bees; US Environmental Protection Agency: Washington, DC, USA, 2016. [Google Scholar]
- Decourtye, A.; Devillers, J.; Aupinel, P.; Brun, F.; Bagnis, C.; Fourrier, J.; Gauthier, M. Honeybee Tracking with Microchips: A New Methodology to Measure the Effects of Pesticides. Ecotoxicology 2011, 20, 429–437. [Google Scholar] [CrossRef]
- Aliouane, Y.; El Hassani, A.K.; Gary, V.; Armengaud, C.; Lambin, M.; Gauthier, M. Subchronic Exposure of Honeybees to Sublethal Doses of Pesticides: Effects on Behavior. Environ. Toxicol. Chem. 2009, 28, 113–122. [Google Scholar] [CrossRef]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.-F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A Common Pesticide Decreases Foraging Success and Survival in Honey Bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef]
- El Hassani, A.K.; Dacher, M.; Gary, V.; Lambin, M.; Gauthier, M.; Armengaud, C. Effects of Sublethal Doses of Acetamiprid and Thiamethoxam on the Behavior of the Honeybee (Apis Mellifera). Arch. Environ. Contam. Toxicol. 2008, 54, 653–661. [Google Scholar] [CrossRef]
- Decourtye, A.; Devillers, J.; Genecque, E.; Menach, K.L.; Budzinski, H.; Cluzeau, S.; Pham-Delègue, M.H. Comparative Sublethal Toxicity of Nine Pesticides on Olfactory Learning Performances of the Honeybee Apis Mellifera. Arch. Environ. Contam. Toxicol. 2005, 48, 242–250. [Google Scholar] [CrossRef]
- Williamson, S.M.; Wright, G.A. Exposure to Multiple Cholinergic Pesticides Impairs Olfactory Learning and Memory in Honeybees. J. Exp. Biol. 2013, 216, 1799–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siviter, H.; Koricheva, J.; Brown, M.J.F.; Leadbeater, E. Quantifying the Impact of Pesticides on Learning and Memory in Bees. J. Appl. Ecol. 2018, 55, 2812–2821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.Y.; Anelli, C.M.; Sheppard, W.S. Sub-Lethal Effects of Pesticide Residues in Brood Comb on Worker Honey Bee (Apis Mellifera) Development and Longevity. PLoS ONE 2011, 6, e14720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid Clothianidin Adversely Affects Insect Immunity and Promotes Replication of a Viral Pathogen in Honey Bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijverberg, H.P.M.; Vanden Bercken, J. Neurotoxicological Effects and the Mode of Action of Pyrethroid Insecticides. Crit. Rev. Toxicol. 1990, 21, 105–126. [Google Scholar] [CrossRef] [PubMed]
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V.; et al. Systemic Insecticides (Neonicotinoids and Fipronil): Trends, Uses, Mode of Action and Metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef]
- Johnson, R.M. Honey Bee Toxicology. Annu. Rev. Entomol. 2015, 60, 415–434. [Google Scholar] [CrossRef] [Green Version]
- Casida, J.E.; Durkin, K.A. Neuroactive Insecticides: Targets, Selectivity, Resistance, and Secondary Effects. Annu. Rev. Entomol. 2013, 58, 99–117. [Google Scholar] [CrossRef]
- Hardstone, M.C.; Scott, J.G. Is Apis Mellifera More Sensitive to Insecticides than Other Insects? Pest Manag. Sci. 2010, 66, 1171–1180. [Google Scholar] [CrossRef]
- Sanchez-Bayo, F.; Goka, K. Pesticide Residues and Bees—A Risk Assessment. PLoS ONE 2014, 9, e94482. [Google Scholar] [CrossRef] [Green Version]
- Urlacher, E.; Monchanin, C.; Rivière, C.; Richard, F.-J.; Lombardi, C.; Michelsen-Heath, S.; Hageman, K.J.; Mercer, A.R. Measurements of Chlorpyrifos Levels in Forager Bees and Comparison with Levels That Disrupt Honey Bee Odor-Mediated Learning Under Laboratory Conditions. J. Chem. Ecol. 2016, 42, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Latham, M.; Hester, P.G.; Frommer, R.L.; Brock, C. Impact of Naled on Honey Bee Apis Mellifera L. Survival and Productivity: Aerial ULV Application Using a Flat-Fan Nozzle System. Arch. Environ. Contam. Toxicol. 2003, 45, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Barnett, E.A.; Charlton, A.J.; Fletcher, M.R. Incidents of Bee Poisoning with Pesticides in the United Kingdom, 1994–2003. Pest Manag. Sci. 2007, 63, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of Pyrethroid Neurotoxicity: Implications for Cumulative Risk Assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef]
- Johnson, R.M.; Dahlgren, L.; Siegfried, B.D.; Ellis, M.D. Acaricide, Fungicide and Drug Interactions in Honey Bees (Apis Mellifera). PLoS ONE 2013, 8, e54092. [Google Scholar] [CrossRef] [Green Version]
- Dai, P.-L.; Wang, Q.; Sun, J.-H.; Liu, F.; Wang, X.; Wu, Y.-Y.; Zhou, T. Effects of Sublethal Concentrations of Bifenthrin and Deltamethrin on Fecundity, Growth, and Development of the Honeybee Apis Mellifera Ligustica. Environ. Toxicol. Chem. 2010, 29, 644–649. [Google Scholar] [CrossRef]
- Liao, C.; He, X.; Wang, Z.; Barron, A.B.; Zhang, B.; Zeng, Z.; Wu, X. Short-Term Exposure to Lambda-Cyhalothrin Negatively Affects the Survival and Memory-Related Characteristics of Worker Bees Apis Mellifera. Arch. Environ. Contam. Toxicol. 2018, 75, 59–65. [Google Scholar] [CrossRef]
- Schneider, C.W.; Tautz, J.; Grünewald, B.; Fuchs, S. RFID Tracking of Sublethal Effects of Two Neonicotinoid Insecticides on the Foraging Behavior of Apis Mellifera. PLoS ONE 2012, 7, e30023. [Google Scholar] [CrossRef] [Green Version]
- Guez, D. A Common Pesticide Decreases Foraging Success and Survival in Honey Bees: Questioning the Ecological Relevance. Front. Physiol. 2013, 4, 37. [Google Scholar] [CrossRef] [Green Version]
- Cresswell, J.E. A Meta-Analysis of Experiments Testing the Effects of a Neonicotinoid Insecticide (Imidacloprid) on Honey Bees. Ecotoxicology 2011, 20, 149–157. [Google Scholar] [CrossRef]
- Tsvetkov, N.; Samson-Robert, O.; Sood, K.; Patel, H.S.; Malena, D.A.; Gajiwala, P.H.; Maciukiewicz, P.; Fournier, V.; Zayed, A. Chronic Exposure to Neonicotinoids Reduces Honey Bee Health near Corn Crops. Science 2017, 356, 1395–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabea, E.I.; Nasr, H.M.; Badawy, M.E.I. Toxic Effect and Biochemical Study of Chlorfluazuron, Oxymatrine, and Spinosad on Honey Bees (Apis Mellifera). Arch. Environ. Contam. Toxicol. 2010, 58, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.E. Effects of a Juvenile Hormone Analogue on Honey Bee Foraging Behaviour and Alarm Pheromone Production. J. Insect Physiol. 1985, 31, 277–282. [Google Scholar] [CrossRef]
- Tasei, J.-N. Effects of Insect Growth Regulators on Honey Bees and Non-Apis Bees. A Review. Apidologie 2001, 32, 527–545. [Google Scholar] [CrossRef] [Green Version]
- Fine, J.D.; Mullin, C.A.; Frazier, M.T.; Reynolds, R.D. Field Residues and Effects of the Insect Growth Regulator Novaluron and Its Major Co-Formulant N-Methyl-2-Pyrrolidone on Honey Bee Reproduction and Development. J. Econ. Entomol. 2017, 110, 1993–2001. [Google Scholar] [CrossRef] [PubMed]
- Pener, M.P.; Dhadialla, T.S. Chapter One—An Overview of Insect Growth Disruptors; Applied Aspects. In Advances in Insect Physiology; Dhadialla, T.S., Ed.; Insect Growth Disruptors; Academic Press: Cambridge, MA, USA, 2012; Volume 43, pp. 1–162. [Google Scholar] [CrossRef]
- Johansen, C.A. Pesticides and Pollinators. Annu. Rev. Entomol. 1977, 22, 177–192. [Google Scholar] [CrossRef] [Green Version]
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Diet Effects on Honeybee Immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef] [Green Version]
- Motta, E.V.; Raymann, K.; Moran, N.A. Glyphosate Perturbs the Gut Microbiota of Honey Bees. Proc. Natl. Acad. Sci. USA 2018, 115, 10305–10310. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, D.E.; Ilina, N.; Pagano, E.A.; Zavala, J.A.; Farina, W.M. Glyphosate Affects the Larval Development of Honey Bees Depending on the Susceptibility of Colonies. PLoS ONE 2018, 13, e0205074. [Google Scholar] [CrossRef] [Green Version]
- Herbert, L.T.; Vázquez, D.E.; Arenas, A.; Farina, W.M. Effects of Field-Realistic Doses of Glyphosate on Honeybee Appetitive Behaviour. J. Exp. Biol. 2014, 217, 3457–3464. [Google Scholar] [CrossRef] [Green Version]
- Liao, L.-H.; Wu, W.-Y.; Berenbaum, M.R. Behavioral Responses of Honey Bees ( Apis Mellifera ) to Natural and Synthetic Xenobiotics in Food. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ostiguy, N.; Drummond, F.A.; Aronstein, K.; Eitzer, B.; Ellis, J.D.; Spivak, M.; Sheppard, W.S. Honey Bee Exposure to Pesticides: A Four-Year Nationwide Study. Insects 2019, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Helmer, S.H.; Kerbaol, A.; Aras, P.; Jumarie, C.; Boily, M. Effects of Realistic Doses of Atrazine, Metolachlor, and Glyphosate on Lipid Peroxidation and Diet-Derived Antioxidants in Caged Honey Bees (Apis Mellifera). Environ. Sci. Pollut. Res. 2015, 22, 8010–8021. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. Disruption of Quercetin Metabolism by Fungicide Affects Energy Production in Honey Bees (Apis Mellifera). Proc. Natl. Acad. Sci. USA 2017, 114, 2538–2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandame, R.; Belzunces, L.P. Joint Actions of Deltamethrin and Azole Fungicides on Honey Bee Thermoregulation. Neurosci. Lett. 1998, 251, 57–60. [Google Scholar] [CrossRef]
- Wade, A.; Lin, C.-H.; Kurkul, C.; Regan, E.R.; Johnson, R.M. Combined Toxicity of Insecticides and Fungicides Applied to California Almond Orchards to Honey Bee Larvae and Adults. Insects 2019, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Forkpah, C.; Dixon, L.R.; Fahrbach, S.E.; Rueppell, O. Xenobiotic Effects on Intestinal Stem Cell Proliferation in Adult Honey Bee (Apis Mellifera L) Workers. PLoS ONE 2014, 9, e91180. [Google Scholar] [CrossRef]
- Johnson, R.T.; Dickersin, K. Publication Bias against Negative Results from Clinical Trials: Three of the Seven Deadly Sins. Nat. Clin. Pract. Neurol. 2007, 3, 590–591. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular Mechanisms of Metabolic Resistance to Synthetic and Natural Xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect P450 Inhibitors and Insecticides: Challenges and Opportunities. Pest Manag. Sci. 2015, 71, 793–800. [Google Scholar] [CrossRef]
- Jones, D.G. Piperonyl Butoxide; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Zhu, W.; Schmehl, D.R.; Mullin, C.A.; Frazier, J.L. Four Common Pesticides, Their Mixtures and a Formulation Solvent in the Hive Environment Have High Oral Toxicity to Honey Bee Larvae. PLoS ONE 2014, 9, e77547. [Google Scholar] [CrossRef] [PubMed]
- Cousin, M.; Silva-Zacarin, E.; Kretzschmar, A.; Maataoui, M.E.; Brunet, J.-L.; Belzunces, L.P. Size Changes in Honey Bee Larvae Oenocytes Induced by Exposure to Paraquat at Very Low Concentrations. PLoS ONE 2013, 8, e65693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregorc, A.; Ellis, J.D. Cell Death Localization in Situ in Laboratory Reared Honey Bee (Apis Mellifera L.) Larvae Treated with Pesticides. Pestic. Biochem. Physiol. 2011, 99, 200–207. [Google Scholar] [CrossRef]
- Mussen, E.C.; Lopez, J.E.; Peng, C.Y. Effects of Selected Fungicides on Growth and Development of Larval Honey Bees, Apis Mellifera L. (Hymenoptera: Apidae). Environ. Entomol. 2004, 33, 1151–1154. [Google Scholar] [CrossRef] [Green Version]
- Atkins, E.L.; Kellum, D. Comparative Morphogenic and Toxicity Studies on the Effect of Pesticides on Honeybee Brood. J. Apic. Res. 1986, 25, 242–255. [Google Scholar] [CrossRef]
- Dahlgren, L.; Johnson, R.M.; Siegfried, B.D.; Ellis, M.D. Comparative Toxicity of Acaricides to Honey Bee (Hymenoptera: Apidae) Workers and Queens. J. Econ. Entomol. 2012, 105, 1895–1902. [Google Scholar] [CrossRef]
- Rinkevich, F.D.; Margotta, J.W.; Pittman, J.M.; Danka, R.G.; Tarver, M.R.; Ottea, J.A.; Healy, K.B. Genetics, Synergists, and Age Affect Insecticide Sensitivity of the Honey Bee, Apis Mellifera. PLoS ONE 2015, 10, e0139841. [Google Scholar] [CrossRef] [Green Version]
- Smirle, M.J.; Winston, M.L. Intercolony Variation in Pesticide Detoxification by the Honey Bee (Hymenoptera: Apidae). J. Econ. Entomol. 1987, 80, 5–8. [Google Scholar] [CrossRef]
- Wahl, O.; Ulm, K. Influence of Pollen Feeding and Physiological Condition on Pesticide Sensitivity of the Honey Bee Apis Mellifera Carnica. Oecologia 1983, 59, 106–128. [Google Scholar] [CrossRef]
- Liao, L.-H.; Wu, W.-Y.; Berenbaum, M.R. Impacts of Dietary Phytochemicals in the Presence and Absence of Pesticides on Longevity of Honey Bees (Apis Mellifera). Insects 2017, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. CYP9Q-Mediated Detoxification of Acaricides in the Honey Bee (Apis Mellifera). Proc. Natl. Acad. Sci. USA 2011, 108, 12657–12662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honeybee Genome Sequencing Consortium. Insights into Social Insects from the Genome of the Honeybee Apis Mellifera. Nature 2006, 443, 931–949. [Google Scholar] [CrossRef]
- Claudianos, C.; Ranson, H.; Johnson, R.M.; Biswas, S.; Schuler, M.A.; Berenbaum, M.R.; Feyereisen, R.; Oakeshott, J.G. A Deficit of Detoxification Enzymes: Pesticide Sensitivity and Environmental Response in the Honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef] [Green Version]
- Fine, J.D.; Cox-Foster, D.L.; Mullin, C.A. An Inert Pesticide Adjuvant Synergizes Viral Pathogenicity and Mortality in Honey Bee Larvae. Sci. Rep. 2017, 7, 40499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettis, J.S. A Scientific Note on Varroa Destructor Resistance to Coumaphos in the United States. Apidologie 2004, 35, 91–92. [Google Scholar] [CrossRef] [Green Version]
- Haarmann, T.; Spivak, M.; Weaver, D.; Weaver, B.; Glenn, T. Effects of Fluvalinate and Coumaphos on Queen Honey Bees (Hymenoptera: Apidae) in Two Commercial Queen Rearing Operations. J. Econ. Entomol. 2002, 95, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.A.; Hood, W.M.; Pietravalle, S.; Delaplane, K.S. Field-Level Sublethal Effects of Approved Bee Hive Chemicals on Honey Bees (Apis Mellifera L). PLoS ONE 2013, 8, e76536. [Google Scholar] [CrossRef] [Green Version]
- Frost, E.H.; Shutler, D.; Hillier, N.K. Effects of Fluvalinate on Honey Bee Learning, Memory, Responsiveness to Sucrose, and Survival. J. Exp. Biol. 2013, 216, 2931–2938. [Google Scholar] [CrossRef] [Green Version]
- Floris, I.; Satta, A.; Cabras, P.; Garau, V.L.; Angioni, A. Comparison between Two Thymol Formulations in the Control of Varroa Destructor: Effectiveness, Persistence, and Residues. J. Econ. Entomol. 2004, 97, 187–191. [Google Scholar] [CrossRef]
- Charpentier, G.; Vidau, C.; Ferdy, J.-B.; Tabart, J.; Vetillard, A. Lethal and Sub-lethal Effects of Thymol on Honeybee (Apis Mellifera) Larvae Reared in Vitro. Pest Manag. Sci. 2014, 70, 140–147. [Google Scholar] [CrossRef]
- Guzmán-Novoa, E.; Eccles, L.; Calvete, Y.; Mcgowan, J.; Kelly, P.G.; Correa-Benítez, A. Varroa Destructor Is the Main Culprit for the Death and Reduced Populations of Overwintered Honey Bee (Apis Mellifera) Colonies in Ontario, Canada. Apidologie 2010, 41, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Le Conte, Y.; Ellis, M.; Ritter, W. Varroa Mites and Honey Bee Health: Can Varroa Explain Part of the Colony Losses? Apidologie 2010, 41, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Amdam, G.V.; Hartfelder, K.; Norberg, K.; Hagen, A.; Omholt, S.W. Altered Physiology in Worker Honey Bees (Hymenoptera: Apidae) Infested with the Mite Varroa Destructor (Acari: Varroidae): A Factor in Colony Loss During Overwintering? J. Econ. Entomol. 2004, 97, 741–747. [Google Scholar] [CrossRef]
- Kralj, J.; Brockmann, A.; Fuchs, S.; Tautz, J. The Parasitic Mite Varroa Destructor Affects Non-Associative Learning in Honey Bee Foragers, Apis mellifera L. J. Comp. Physiol. A 2007, 193, 363–370. [Google Scholar] [CrossRef]
- Aronstein, K.A.; Saldivar, E.; Vega, R.; Westmiller, S.; Douglas, A.E. How Varroa Parasitism Affects the Immunological and Nutritional Status of the Honey Bee, Apis Mellifera. Insects 2012, 3, 601–615. [Google Scholar] [CrossRef] [Green Version]
- Dolezal, A.G.; Carrillo-Tripp, J.; Miller, W.A.; Bonning, B.C.; Toth, A.L. Intensively Cultivated Landscape and Varroa Mite Infestation Are Associated with Reduced Honey Bee Nutritional State. PLoS ONE 2016, 11, e0153531. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and Control of Varroa Destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- Nazzi, F.; Le Conte, Y. Ecology of Varroa destructor, the Major Ectoparasite of the Western Honey Bee, Apis mellifera. Annu. Rev. Entomol. 2016, 61, 417–432. [Google Scholar] [CrossRef] [Green Version]
- Grozinger, C.M.; Flenniken, M.L. Bee Viruses: Ecology, Pathogenicity, and Impacts. Annu. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef]
- Bonning, B.C. The Dicistroviridae: An Emerging Family of Invertebrate Viruses. Virol. Sin. 2009, 24, 415. [Google Scholar] [CrossRef]
- De Miranda, J.R.; Cordoni, G.; Budge, G. The Acute Bee Paralysis Virus–Kashmir Bee Virus–Israeli Acute Paralysis Virus Complex. J. Invertebr. Pathol. 2010, 103, S30–S47. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Lavi, S.; Mozes-Koch, R.; Gantman, Y.; Peretz, Y.; Edelbaum, O.; Tanne, E.; Sela, I. Isolation and Characterization of Israeli Acute Paralysis Virus, a Dicistrovirus Affecting Honeybees in Israel: Evidence for Diversity Due to Intra- and Inter-Species Recombination. J. Gen. Virol. 2007, 88, 3428–3438. [Google Scholar] [CrossRef] [PubMed]
- Boncristiani, H.F.; Evans, J.D.; Chen, Y.; Pettis, J.; Murphy, C.; Lopez, D.L.; Simone-Finstrom, M.; Strand, M.; Tarpy, D.R.; Rueppell, O. In Vitro Infection of Pupae with Israeli Acute Paralysis Virus Suggests Disturbance of Transcriptional Homeostasis in Honey Bees (Apis Mellifera). PLoS ONE 2013, 8, e73429. [Google Scholar] [CrossRef] [PubMed]
- De Miranda, J.R.; Genersch, E. Deformed Wing Virus. J. Invertebr. Pathol. 2010, 103 (Suppl. 1), S48–S61. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Wood, G.R.; Fannon, J.M.; Moore, J.D.; Bull, J.C.; Chandler, D.; Mead, A.; Burroughs, N.; Evans, D.J. A Virulent Strain of Deformed Wing Virus (DWV) of Honeybees (Apis Mellifera) Prevails after Varroa Destructor-Mediated, or In Vitro, Transmission. PLoS Pathog. 2014, 10, e1004230. [Google Scholar] [CrossRef] [Green Version]
- Yañez, O.; Chávez-Galarza, J.; Tellgren-Roth, C.; Pinto, M.A.; Neumann, P.; de Miranda, J.R. The Honeybee (Apis Mellifera) Developmental State Shapes the Genetic Composition of the Deformed Wing Virus-A Quasispecies during Serial Transmission. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global Honey Bee Viral Landscape Altered by a Parasitic Mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Childers, A.K.; Chen, Y.; Madella, S.; Nessa, A.; van Engelsdorp, D.; Evans, J.D. Recent Spread of Varroa Destructor Virus-1, a Honey Bee Pathogen, in the United States. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kevill, J.L.; de Souza, F.S.; Sharples, C.; Oliver, R.; Schroeder, D.C.; Martin, S.J. DWV-A Lethal to Honey Bees (Apis Mellifera): A Colony Level Survey of DWV Variants (A, B, and C) in England, Wales, and 32 States across the US. Viruses 2019, 11, 426. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.P.; Pettis, J.S.; Corona, M.; Chen, W.P.; Li, C.J.; Spivak, M.; Visscher, P.K.; DeGrandi-Hoffman, G.; Boncristiani, H.; Zhao, Y.; et al. Israeli Acute Paralysis Virus: Epidemiology, Pathogenesis and Implications for Honey Bee Health. PLoS Pathog. 2014, 10, e1004261. [Google Scholar] [CrossRef]
- Geffre, A.C.; Gernat, T.; Harwood, G.P.; Jones, B.M.; Gysi, D.M.; Hamilton, A.R.; Bonning, B.C.; Toth, A.L.; Robinson, G.E.; Dolezal, A.G. Honey Bee Virus Causes Context-Dependent Changes in Host Social Behavior. Proc. Natl. Acad. Sci. USA 2020, 117, 10406–10413. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Evans, J.; Feldlaufer, M. Horizontal and Vertical Transmission of Viruses in the Honey Bee, Apis Mellifera. J. Invertebr. Pathol. 2006, 92, 152–159. [Google Scholar] [CrossRef]
- Amiri, E.; Seddon, G.; Zuluaga Smith, W.; Strand, M.K.; Tarpy, D.R.; Rueppell, O. Israeli Acute Paralysis Virus: Honey Bee Queen–Worker Interaction and Potential Virus Transmission Pathways. Insects 2019, 10, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, C.; Schröder, M.; Gisder, S.; Genersch, E. Vertical-Transmission Routes for Deformed Wing Virus of Honeybees (Apis Mellifera). J. Gen. Virol. 2007, 88, 2329–2336. [Google Scholar] [CrossRef] [PubMed]
- Levitt, A.L.; Singh, R.; Cox-Foster, D.L.; Rajotte, E.; Hoover, K.; Ostiguy, N.; Holmes, E.C. Cross-Species Transmission of Honey Bee Viruses in Associated Arthropods. Virus Res. 2013, 176, 232–240. [Google Scholar] [CrossRef]
- Genersch, E.; Yue, C.; Fries, I.; de Miranda, J.R. Detection of Deformed Wing Virus, a Honey Bee Viral Pathogen, in Bumble Bees (Bombus Terrestris and Bombus Pascuorum) with Wing Deformities. J. Invertebr. Pathol. 2006, 91, 61–63. [Google Scholar] [CrossRef]
- Fürst, M.A.; McMahon, D.P.; Osborne, J.L.; Paxton, R.J.; Brown, M.J.F. Disease Associations between Honeybees and Bumblebees as a Threat to Wild Pollinators. Nature 2014, 506, 364–366. [Google Scholar] [CrossRef]
- Flenniken, M.L.; Andino, R. Non-Specific DsRNA-Mediated Antiviral Response in the Honey Bee. PLoS ONE 2013, 8, e77263. [Google Scholar] [CrossRef]
- Cerutti, H.; Casas-Mollano, J.A. On the Origin and Functions of RNA-Mediated Silencing: From Protists to Man. Curr. Genet. 2006, 50, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Maori, E.; Paldi, N.; Shafir, S.; Kalev, H.; Tsur, E.; Glick, E.; Sela, I. IAPV, a Bee-Affecting Virus Associated with Colony Collapse Disorder Can Be Silenced by DsRNA Ingestion. Insect Mol. Biol. 2009, 18, 55–60. [Google Scholar] [CrossRef]
- Brutscher, L.M.; Daughenbaugh, K.F.; Flenniken, M.L. Virus and DsRNA-Triggered Transcriptional Responses Reveal Key Components of Honey Bee Antiviral Defense. Sci. Rep. 2017, 7, 6448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingsolver, M.B.; Huang, Z.; Hardy, R.W. Insect Antiviral Innate Immunity: Pathways, Effectors, and Connections. J. Mol. Biol. 2013, 425, 4921–4936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkling, S.H.; van Rij, R.P. Beyond RNAi: Antiviral Defense Strategies in Drosophila and Mosquito. J. Insect Physiol. 2013, 59, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Khush, R.S.; Leulier, F.; Lemaitre, B. Drosophila Immunity: Two Paths to NF-ΚB. Trends Immunol. 2001, 22, 260–264. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.; Jan, E.; Sarnow, P.; Schneider, D. The Imd Pathway Is Involved in Antiviral Immune Responses in Drosophila. PLoS ONE 2009, 4, e7436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, Á.G.; Naylor, H.; Esteves, S.S.; Pais, I.S.; Martins, N.E.; Teixeira, L. The Toll-Dorsal Pathway Is Required for Resistance to Viral Oral Infection in Drosophila. PLoS Pathog. 2014, 10, e1004507. [Google Scholar] [CrossRef] [Green Version]
- Zambon, R.A.; Nandakumar, M.; Vakharia, V.N.; Wu, L.P. The Toll Pathway Is Important for an Antiviral Response in Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 7257–7262. [Google Scholar] [CrossRef] [Green Version]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes Aegypti Toll Pathway Controls Dengue Virus Infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef]
- Shelly, S.; Lukinova, N.; Bambina, S.; Berman, A.; Cherry, S. Autophagy Is an Essential Component of Drosophila Immunity against Vesicular Stomatitis Virus. Immunity 2009, 30, 588–598. [Google Scholar] [CrossRef] [Green Version]
- Lamiable, O.; Arnold, J.; de Faria, I.J.S.; Olmo, R.P.; Bergami, F.; Meignin, C.; Hoffmann, J.A.; Marques, J.T.; Imler, J.-L. Analysis of the Contribution of Hemocytes and Autophagy to Drosophila Antiviral Immunity. J. Virol. 2016, 90, 5415–5426. [Google Scholar] [CrossRef] [Green Version]
- McMenamin, A.J.; Daughenbaugh, K.F.; Flenniken, M.L. The Heat Shock Response in the Western Honey Bee (Apis Mellifera) Is Antiviral. Viruses 2020, 12, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, S.; Kobayashi, M.; Yoda, M.; Sakaguchi, Y.; Katsuma, S.; Suzuki, T.; Tomari, Y. Hsc70/Hsp90 Chaperone Machinery Mediates ATP-Dependent RISC Loading of Small RNA Duplexes. Mol. Cell 2010, 39, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Cremer, S.; Armitage, S.A.O.; Schmid-Hempel, P. Social Immunity. Curr. Biol. 2007, 17, R693–R702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, S.; Pull, C.D.; Fürst, M.A. Social Immunity: Emergence and Evolution of Colony-Level Disease Protection. Annu. Rev. Entomol. 2018, 63, 105–123. [Google Scholar] [CrossRef] [PubMed]
- López, J.H.; Schuehly, W.; Crailsheim, K.; Riessberger-Gallé, U. Trans-Generational Immune Priming in Honeybees. Proc. R. Soc. Lond. B Biol. Sci. 2014, 281, 20140454. [Google Scholar] [CrossRef] [PubMed]
- Salmela, H.; Amdam, G.V.; Freitak, D. Transfer of Immunity from Mother to Offspring Is Mediated via Egg-Yolk Protein Vitellogenin. PLoS Pathog. 2015, 11, e1005015. [Google Scholar] [CrossRef] [Green Version]
- Harwood, G.; Amdam, G.; Freitak, D. The Role of Vitellogenin in the Transfer of Immune Elicitors from Gut to Hypopharyngeal Glands in Honey Bees (Apis Mellifera). J. Insect Physiol. 2019, 112, 90–100. [Google Scholar] [CrossRef]
- Maori, E.; Garbian, Y.; Kunik, V.; Mozes-Koch, R.; Malka, O.; Kalev, H.; Sabath, N.; Sela, I.; Shafir, S. A Transmissible RNA Pathway in Honey Bees. Cell Rep. 2019, 27, 1949–1959. [Google Scholar] [CrossRef] [Green Version]
- Coulon, M.; Schurr, F.; Martel, A.-C.; Cougoule, N.; Bégaud, A.; Mangoni, P.; Dalmon, A.; Alaux, C.; Le Conte, Y.; Thiéry, R.; et al. Metabolisation of Thiamethoxam (a Neonicotinoid Pesticide) and Interaction with the Chronic Bee Paralysis Virus in Honeybees. Pestic. Biochem. Physiol. 2018, 144, 10–18. [Google Scholar] [CrossRef]
- Coulon, M.; Schurr, F.; Martel, A.-C.; Cougoule, N.; Begaud, A.; Mangoni, P.; Di Prisco, G.; Dalmon, A.; Alaux, C.; Ribiere-Chabert, M. Influence of Chronic Exposure to Thiamethoxam and Chronic Bee Paralysis Virus on Winter Honey Bees. PLoS ONE 2019, 14, e0220703. [Google Scholar] [CrossRef] [Green Version]
- Diao, Q.; Li, B.; Zhao, H.; Wu, Y.; Guo, R.; Dai, P.; Chen, D.; Wang, Q.; Hou, C. Enhancement of Chronic Bee Paralysis Virus Levels in Honeybees Acute Exposed to Imidacloprid: A Chinese Case Study. Sci. Total Environ. 2018, 630, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Doublet, V.; Labarussias, M.; de Miranda, J.R.; Moritz, R.F.A.; Paxton, R.J. Bees under Stress: Sublethal Doses of a Neonicotinoid Pesticide and Pathogens Interact to Elevate Honey Bee Mortality across the Life Cycle. Environ. Microbiol. 2015, 17, 969–983. [Google Scholar] [CrossRef] [PubMed]
- O’Neal, S.T.; Reeves, A.M.; Fell, R.D.; Brewster, C.C.; Anderson, T.D. Chlorothalonil Exposure Alters Virus Susceptibility and Markers of Immunity, Nutrition, and Development in Honey Bees. J. Insect Sci. 2019, 19, 14. [Google Scholar] [CrossRef] [PubMed]
- Rolke, D.; Fuchs, S.; Grünewald, B.; Gao, Z.; Blenau, W. Large-Scale Monitoring of Effects of Clothianidin-Dressed Oilseed Rape Seeds on Pollinating Insects in Northern Germany: Effects on Honey Bees (Apis Mellifera). Ecotoxicology 2016, 25, 1648–1665. [Google Scholar] [CrossRef] [Green Version]
- Osterman, J.; Wintermantel, D.; Locke, B.; Jonsson, O.; Semberg, E.; Onorati, P.; Forsgren, E.; Rosenkranz, P.; Rahbek-Pedersen, T.; Bommarco, R.; et al. Clothianidin Seed-Treatment Has No Detectable Negative Impact on Honeybee Colonies and Their Pathogens. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Simon-Delso, N.; San Martin, G.; Bruneau, E.; Minsart, L.-A.; Mouret, C.; Hautier, L. Honeybee Colony Disorder in Crop Areas: The Role of Pesticides and Viruses. PLoS ONE 2014, 9, e103073. [Google Scholar] [CrossRef]
- Wintermantel, D.; Locke, B.; Andersson, G.K.S.; Semberg, E.; Forsgren, E.; Osterman, J.; Pedersen, T.R.; Bommarco, R.; Smith, H.G.; Rundlöf, M.; et al. Field-Level Clothianidin Exposure Affects Bumblebees but Generally Not Their Pathogens. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rajak, P.; Dutta, M.; Roy, S. Altered Differential Hemocyte Count in 3rd Instar Larvae of Drosophila Melanogaster as a Response to Chronic Exposure of Acephate. Interdiscip. Toxicol. 2015, 8, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Walderdorff, L.; Laval-Gilly, P.; Wechtler, L.; Bonnefoy, A.; Falla-Angel, J. Phagocytic Activity of Human Macrophages and Drosophila Hemocytes after Exposure to the Neonicotinoid Imidacloprid. Pestic. Biochem. Physiol. 2019, 160, 95–101. [Google Scholar] [CrossRef]
- Walderdorff, L.; Laval-Gilly, P.; Bonnefoy, A.; Falla-Angel, J. Imidacloprid Intensifies Its Impact on Honeybee and Bumblebee Cellular Immune Response When Challenged with LPS (Lippopolysacharide) of Escherichia Coli. J. Insect Physiol. 2018, 108, 17–24. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Wang, X. Exploring the Multilevel Hazards of Thiamethoxam Using Drosophila Melanogaster. J. Hazard. Mater. 2020, 384, 121419. [Google Scholar] [CrossRef] [PubMed]
- Pellacani, C.; Costa, L.G. Role of Autophagy in Environmental Neurotoxicity. Environ. Pollut. 2018, 235, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Schmehl, D.R.; Teal, P.E.A.; Frazier, J.L.; Grozinger, C.M. Genomic Analysis of the Interaction between Pesticide Exposure and Nutrition in Honey Bees (Apis Mellifera). J. Insect Physiol. 2014, 71, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.; Son, T.-G.; Kim, S.-Y.; Lee, K.-Y. Differential Responses of Apis Mellifera Heat Shock Protein Genes to Heat Shock, Flower-Thinning Formulations, and Imidacloprid. J. Asia-Pac. Entomol. 2015, 18, 583–589. [Google Scholar] [CrossRef]
- Reid, W.R.; Zhang, L.; Gong, Y.; Li, T.; Liu, N. Gene Expression Profiles of the Southern House Mosquito Culex Quinquefasciatus during Exposure to Permethrin. Insect Sci. 2018, 25, 439–453. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Alto, B.W.; Shin, D.; Yu, F. The Effect of Permethrin Resistance on Aedes Aegypti Transcriptome Following Ingestion of Zika Virus Infected Blood. Viruses 2018, 10, 470. [Google Scholar] [CrossRef] [Green Version]
- Games, P.D.; Alves, S.N.; Katz, B.B.; Tomich, J.M.; Serrão, J.E. Differential Protein Expression in the Midgut of Culex Quinquefasciatus Mosquitoes Induced by the Insecticide Temephos. Med. Vet. Entomol. 2016, 30, 253–263. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Toll-like Receptors and Their Crosstalk with Other Innate Receptors in Infection and Immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [Green Version]
- Daisley, B.A.; Trinder, M.; McDowell, T.W.; Welle, H.; Dube, J.S.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Neonicotinoid-Induced Pathogen Susceptibility Is Mitigated by Lactobacillus Plantarum Immune Stimulation in a Drosophila Melanogaster Model. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Chmiel, J.A.; Daisley, B.A.; Burton, J.P.; Reid, G. Deleterious Effects of Neonicotinoid Pesticides on Drosophila Melanogaster Immune Pathways. mBio 2019, 10, e01395-19. [Google Scholar] [CrossRef] [Green Version]
- Raymann, K.; Moran, N.A. The Role of the Gut Microbiome in Health and Disease of Adult Honey Bee Workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Yan, Z.; Ma, S.; Yang, Y.; Wang, Q.; Hou, C.; Wu, Y.; Liu, Y.; Diao, Q. The Herbicide Glyphosate Negatively Affects Midgut Bacterial Communities and Survival of Honey Bee during Larvae Reared in Vitro. J. Agric. Food Chem. 2018, 66, 7786–7793. [Google Scholar] [CrossRef] [PubMed]
- Raymann, K.; Motta, E.V.; Girard, C.; Riddington, I.M.; Dinser, J.A.; Moran, N.A. Imidacloprid Decreases Honey Bee Survival Rates but Does Not Affect the Gut Microbiome. Appl. Env. Microbiol. 2018, 84, e00545-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swale, D.R.; Engers, D.W.; Bollinger, S.R.; Gross, A.; Inocente, E.A.; Days, E.; Kanga, F.; Johnson, R.M.; Yang, L.; Bloomquist, J.R.; et al. An Insecticide Resistance-Breaking Mosquitocide Targeting Inward Rectifier Potassium Channels in Vectors of Zika Virus and Malaria. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Eleftherianos, I.; Won, S.; Chtarbanova, S.; Squiban, B.; Ocorr, K.; Bodmer, R.; Beutler, B.; Hoffmann, J.A.; Imler, J.-L. ATP-Sensitive Potassium Channel (KATP)–Dependent Regulation of Cardiotropic Viral Infections. Proc. Natl. Acad. Sci. USA 2011, 108, 12024–12029. [Google Scholar] [CrossRef] [Green Version]
- O’Neal, S.T.; Swale, D.R.; Bloomquist, J.R.; Anderson, T.D. ATP-Sensitive Inwardly Rectifying Potassium Channel Modulators Alter Cardiac Function in Honey Bees. J. Insect Physiol. 2017, 99, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Brandt, A.; Gorenflo, A.; Siede, R.; Meixner, M.; Büchler, R. The Neonicotinoids Thiacloprid, Imidacloprid, and Clothianidin Affect the Immunocompetence of Honey Bees (Apis Mellifera L.). J. Insect Physiol. 2016, 86, 40–47. [Google Scholar] [CrossRef]
- Bitsadze, N.; Jaronski, S.; Khasdan, V.; Abashidze, E.; Abashidze, M.; Latchininsky, A.; Samadashvili, D.; Sokhadze, I.; Rippa, M.; Ishaaya, I.; et al. Joint Action of Beauveria Bassiana and the Insect Growth Regulators Diflubenzuron and Novaluron, on the Migratory Locust, Locusta Migratoria. J. Pest Sci. 2013, 86, 293–300. [Google Scholar] [CrossRef]
- Boucias, D.G.; Nordin, G.L. Methoprene: Nucleopolyhedrosis Virus Interactions in Hyphantria Cunea (Drury). J. Kans. Entomol. Soc. 1980, 53, 56–60. [Google Scholar]
- Furlong, M.J.; Groden, E. Evaluation of Synergistic Interactions Between the Colorado Potato Beetle (Coleoptera: Chrysomelidae) Pathogen Beauveria Bassiana and the Insecticides, Imidacloprid, and Cyromazine. J. Econ. Entomol. 2001, 94, 344–356. [Google Scholar] [CrossRef]
- Cutler, G.C.; Scott-Dupree, C.D.; Sultan, M.; McFarlane, A.D.; Brewer, L. A Large-Scale Field Study Examining Effects of Exposure to Clothianidin Seed-Treated Canola on Honey Bee Colony Health, Development, and Overwintering Success. PeerJ 2014, 2, e652. [Google Scholar] [CrossRef] [PubMed]
- Paleolog, J.; Wilde, J.; Siuda, M.; Bąk, B.; Wójcik, Ł.; Strachecka, A. Imidacloprid Markedly Affects Hemolymph Proteolysis, Biomarkers, DNA Global Methylation, and the Cuticle Proteolytic Layer in Western Honeybees. Apidologie 2020, 51, 1–11. [Google Scholar] [CrossRef] [Green Version]
- De Smet, L.; Hatjina, F.; Ioannidis, P.; Hamamtzoglou, A.; Schoonvaere, K.; Francis, F.; Meeus, I.; Smagghe, G.; de Graaf, D.C. Stress Indicator Gene Expression Profiles, Colony Dynamics and Tissue Development of Honey Bees Exposed to Sub-Lethal Doses of Imidacloprid in Laboratory and Field Experiments. PLoS ONE 2017, 12, e0171529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, M.; Cerrutti, N.; Aupinel, P.; Decourtye, A.; Gayrard, M.; Odoux, J.-F.; Pissard, A.; Rüger, C.; Bretagnolle, V. Reconciling Laboratory and Field Assessments of Neonicotinoid Toxicity to Honeybees. Proc. R. Soc. B Biol. Sci. 2015, 282, 20152110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.; et al. Country-Specific Effects of Neonicotinoid Pesticides on Honey Bees and Wild Bees. Science 2017, 356, 1393–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carleton, J. Environmental Fate and Ecological Risk Assessment for Foliar, Soil Drench, and Seed Treatment Uses of the New Insecticide Flupyradifurone (BYI 02960); United States Environmental Protection Agency: Washington, DC, USA, 2014. [Google Scholar]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harwood, G.P.; Dolezal, A.G. Pesticide–Virus Interactions in Honey Bees: Challenges and Opportunities for Understanding Drivers of Bee Declines. Viruses 2020, 12, 566. https://doi.org/10.3390/v12050566
Harwood GP, Dolezal AG. Pesticide–Virus Interactions in Honey Bees: Challenges and Opportunities for Understanding Drivers of Bee Declines. Viruses. 2020; 12(5):566. https://doi.org/10.3390/v12050566
Chicago/Turabian StyleHarwood, Gyan P., and Adam G. Dolezal. 2020. "Pesticide–Virus Interactions in Honey Bees: Challenges and Opportunities for Understanding Drivers of Bee Declines" Viruses 12, no. 5: 566. https://doi.org/10.3390/v12050566
APA StyleHarwood, G. P., & Dolezal, A. G. (2020). Pesticide–Virus Interactions in Honey Bees: Challenges and Opportunities for Understanding Drivers of Bee Declines. Viruses, 12(5), 566. https://doi.org/10.3390/v12050566