HHV-6 Infection and Chemokine RANTES Signaling Pathway Disturbance in Patients with Autoimmune Thyroiditis

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Groups
2.2. Nucleic Acid Isolation, Complementary DNA (cDNA) Synthesis and Quality Determination
2.3. Detection of HHV-6 Genomic Sequences and Gene Expression Using Nested PCR (nPCR)
2.4. HHV-6 Load Determination Using Quantitative PCR
2.5. Determination of RANTES (CCL5) Level in Peripheral Blood Plasma by ELISA
2.6. Determination of IFNγ, IL-6, TNFα and RANTES (CCL5) Levels in Peripheral Blood Plasma by Suspension Multiplex Immuno Assay (SMIA)
2.7. Immunofluorescence Labelling of Formalin-Fixed Paraffin-Embedded (FFPE) Thyroid Tissues
3. Results
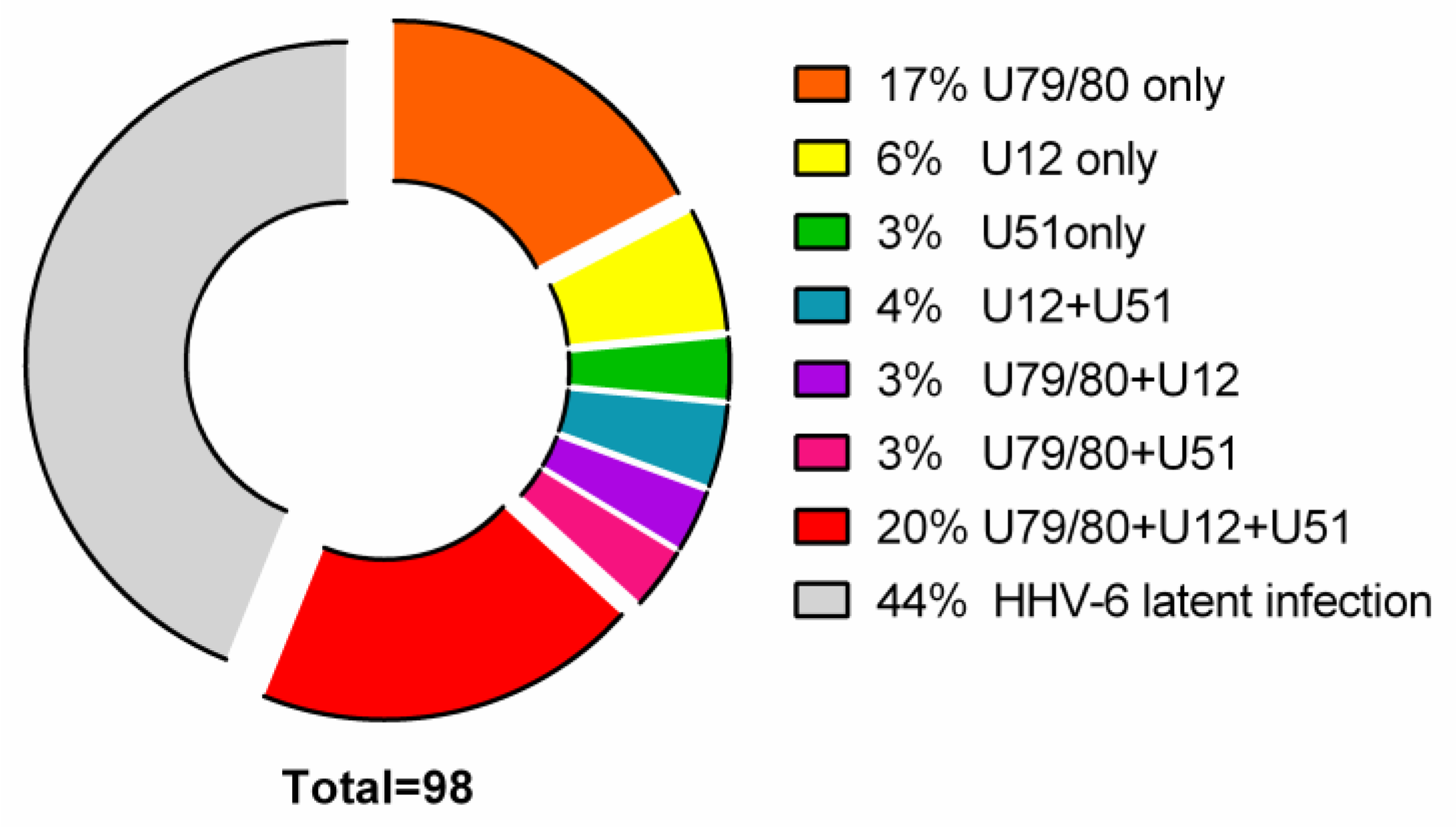
3.1. Detection of HHV-6 Virus Genomic Sequence and Its mRNAs in AIT Patients and Control Group Autopsy Specimens
- 17/98 (17%) AIT patients’ thyroid tissue samples and 8/16 (50%) PBMC samples were positive for HHV-6 U79/80 mRNA.
- 6/98 (6%) AIT patients’ thyroid tissue samples were positive for HHV-6 U12 mRNA, while none of the PBMC samples were HHV-6 U12 mRNA-positive and 3/98 (3%) thyroid tissue samples of AIT patients and 4/16 (25%) PBMC samples were positive for HHV-6 U51 mRNA.
- U79/80 + U12 mRNA was found in 3/98 (3%) AIT patients’ thyroid tissue samples, while none of the PBMC samples were HHV-6 U79/80 + U12 mRNA-positive; U79/80 + U51 mRNAs were present in 3/98 (3%) AIT patients’ thyroid tissue samples and in none of the PBMC samples.
- U79/80 + U12 + U51 mRNAs were present in 19/98 (20%) AIT patients’ thyroid tissue samples and in none of the PBMC samples.
- U12 + U51 mRNAs were found only in 4/98 (4%) AIT patients’ thyroid tissue samples.
- The presence of HHV-6 U79/80 mRNA was only detected in 1/17 autopsy samples (data from previous study [5]), and no HHV-6 U12 or U51 mRNAs were found in this study.
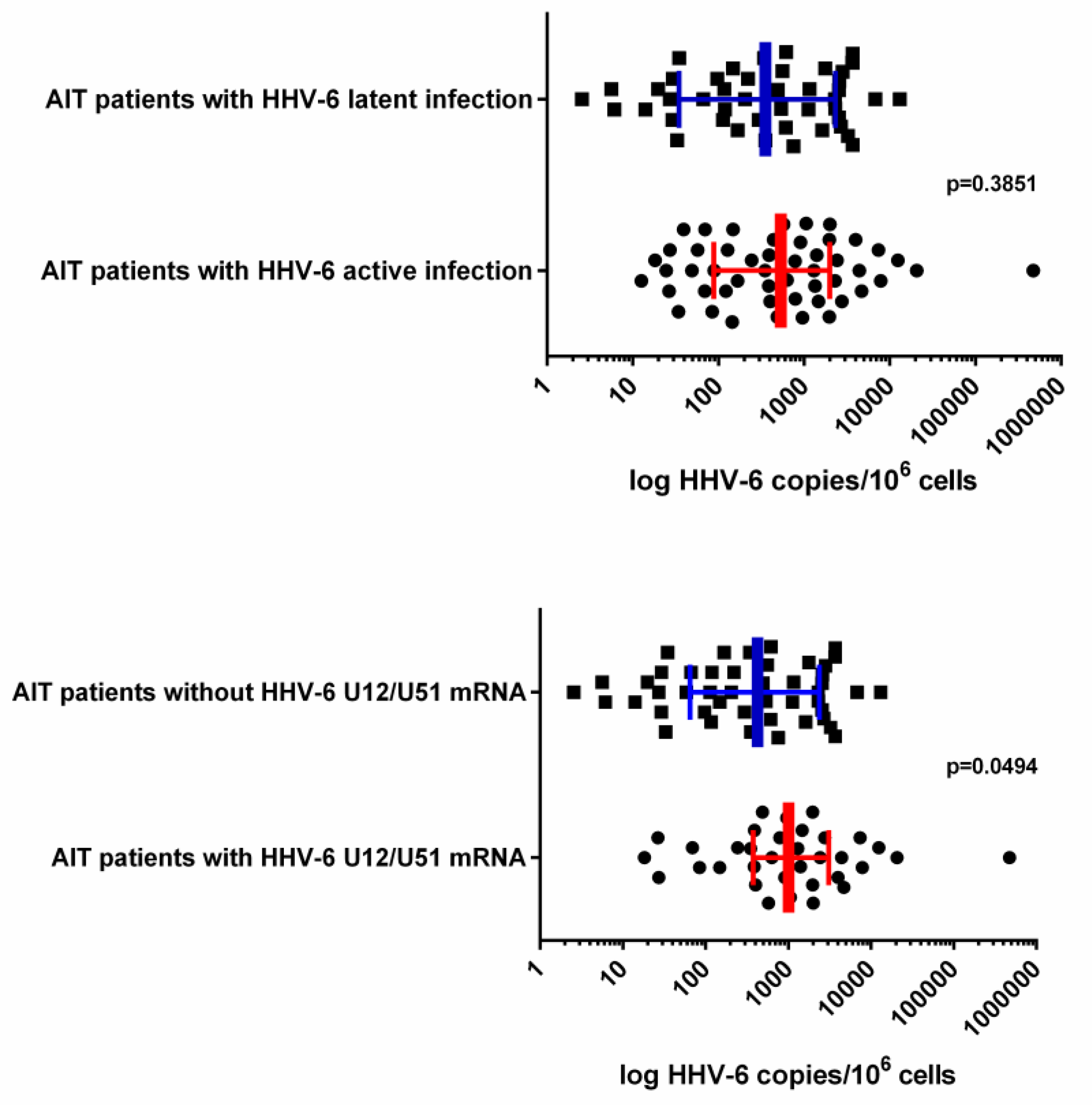
3.2. HHV-6 Load in Thyroid Tissue Samples
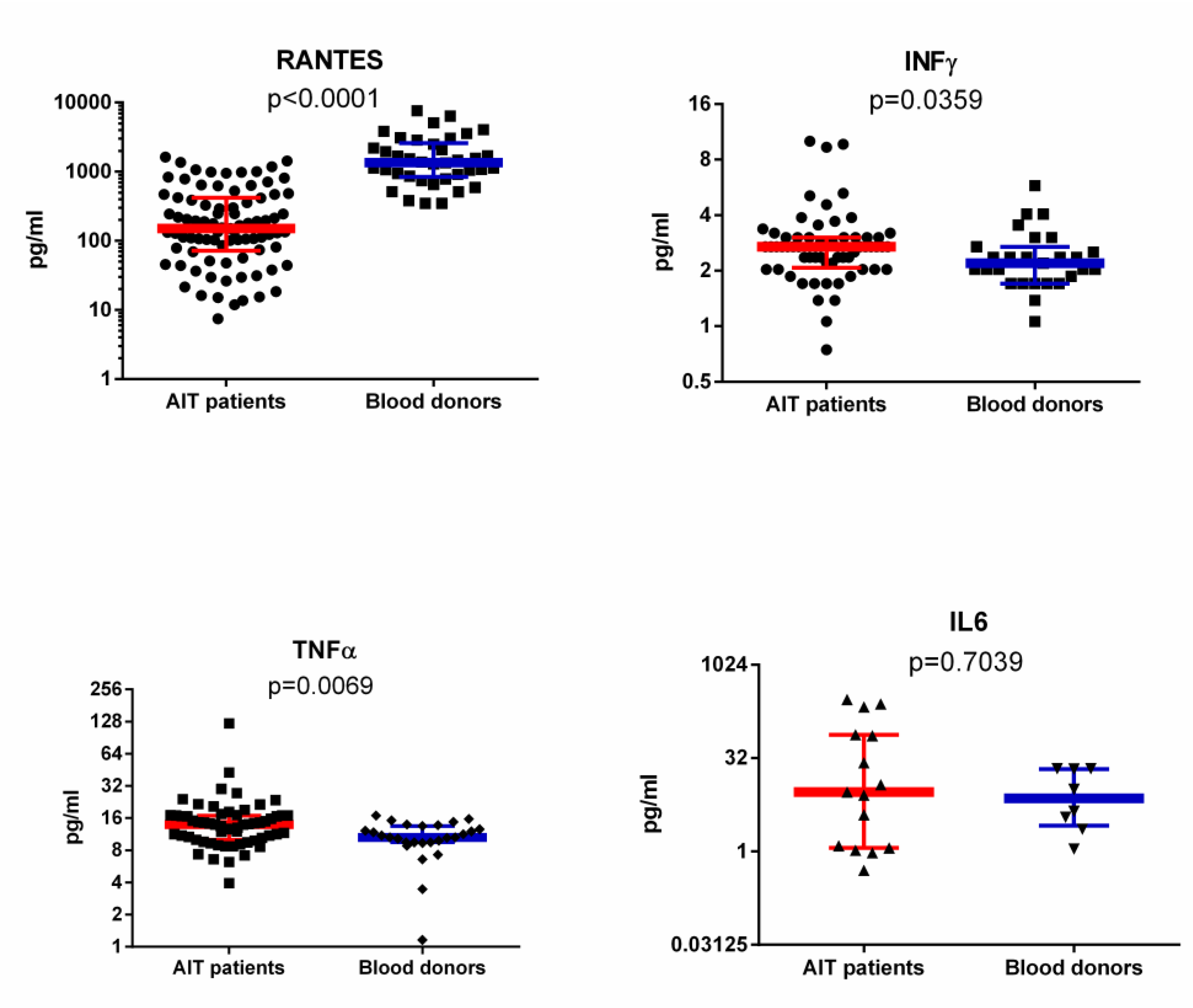
3.3. RANTES Level in Plasma of AIT Patient and Blood Donor Groups Detected with ELISA and SMIA
3.4. IFNγ, IL-6 and TNFα Levels in Plasma of AIT Patient and Blood Donor Groups
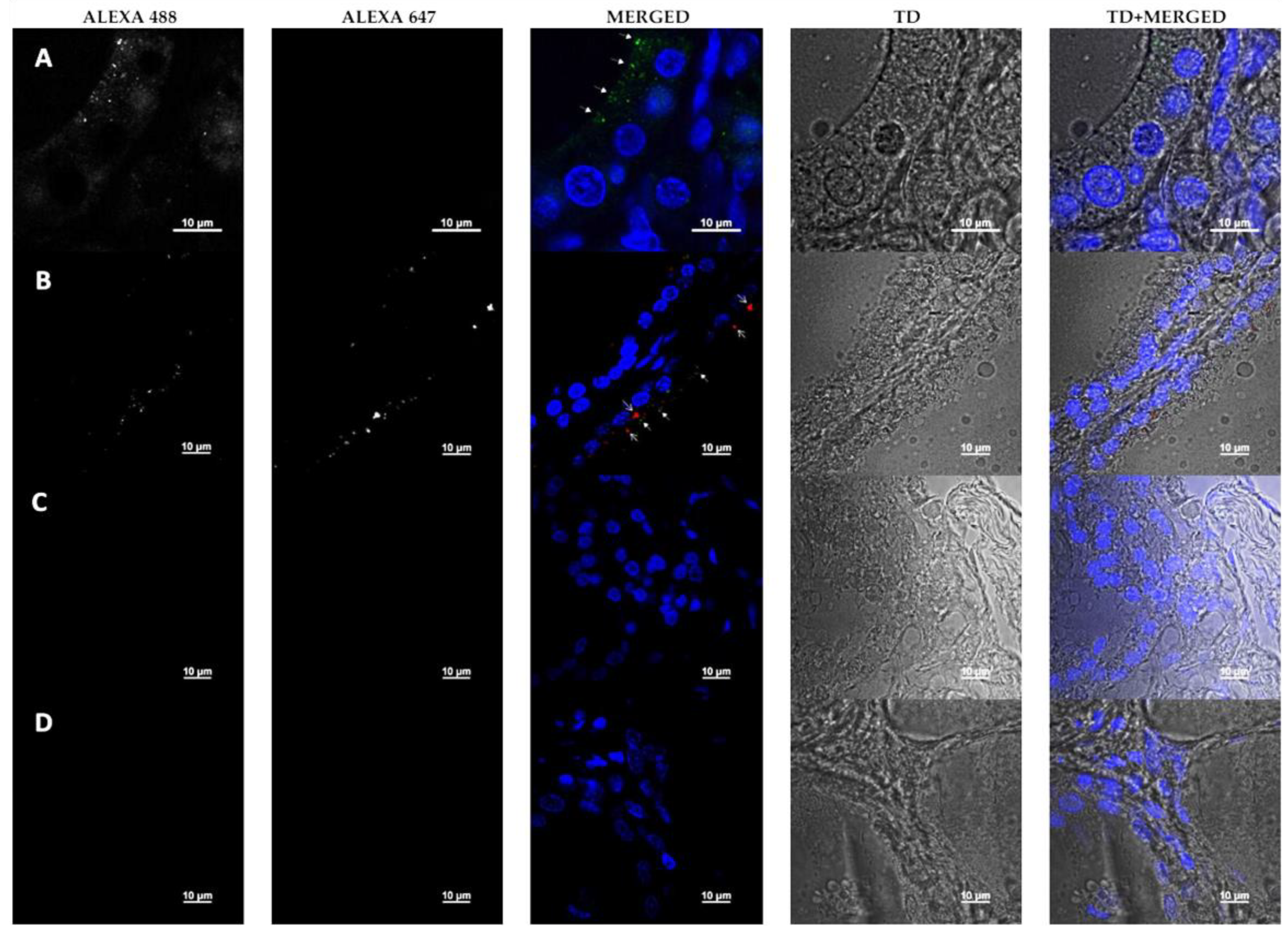
3.5. Detection of HHV-6 Antigen and RANTES by Immunofluorescence Microscopy
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Lerner, A.; Jeremias, P.; Matthias, T. The world incidence and prevalence of autoimmune diseases is increasing. Int. J. Celiac Dis. 2015, 3, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Pennell, L.M.; Galligan, C.L.; Fish, E.N. Sex affects immunity. J. Autoimmun. 2012, 38, J282–J291. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hudnall, S.D. Anatomical mapping of human herpesvirus reservoirs of infection. Mod. Pathol. 2006, 19, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Caselli, E.; Zatelli, M.C.; Rizzo, R.; Benedetti, S.; Martorelli, D.; Trasforini, G.; Cassai, E.; degli Uberti, E.C.; di Luca, D.; Dolcetti, R. Virologic and immunologic evidence supporting an association between HHV-6 and Hashimoto’s thyroiditis. PLoS Pathog. 2012, 8, e1002951. [Google Scholar] [CrossRef] [PubMed]
- Sultanova, A.; Cistjakovs, M.; Gravelsina, S.; Chapenko, S.; Roga, S.; Cunskis, E.; Nora-Krukle, Z.; Groma, V.; Ventina, I.; Murovska, M. Association of active human herpesvirus-6 (HHV-6) infection with autoimmune thyroid gland diseases. Clin. Microbiol. Infect. 2017, 23, 50.e1–50.e5. [Google Scholar] [CrossRef] [Green Version]
- Jasirwan, C.; Furusawa, Y.; Tang, H.; Maeki, T.; Mori, Y. Human herpesvirus-6A gQ1 and gQ2 are critical for human CD46 usage. Microbiol. Immunol. 2014, 58, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Yagasaki, H.; Kato, M.; Shimizu, N.; Shichino, H.; Chin, M.; Mugishima, H. Autoimmune hemolytic anemia and autoimmune neutropenia in a child with erythroblastopenia of childhood (TEC) caused by human herpesvirus-6 (HHV-6). Ann. Hematol. 2011, 90, 851–852. [Google Scholar] [CrossRef]
- Grima, P.; Chiavaroli, R.; Calabrese, P.; Tundo, P.; Grima, P. Severe hepatitis with autoimmune features following a HHV-6: A case report. Cases J. 2008, 1, 110. [Google Scholar] [CrossRef] [Green Version]
- Chapenko, S.; Millers, A.; Nora, Z.; Logina, I.; Kukaine, R.; Murovska, M. Correlation between HHV-6 reactivation and multiple sclerosis disease activity. J. Med. Virol. 2003, 69, 111–117. [Google Scholar] [CrossRef]
- Nora-Krukle, Z.; Chapenko, S.; Logina, I.; Millers, A.; Platkajis, A.; Murovska, M. Human herpesvirus 6 and 7 reactivation and disease activity in multiple sclerosis. Med. Kaunas Lith. 2011, 47, 527–531. [Google Scholar] [CrossRef]
- Tejada-Simon, M.V.; Zang, Y.C.Q.; Hong, J.; Rivera, V.M.; Zhang, J.Z. Cross-reactivity with myelin basic protein and human herpesvirus-6 in multiple sclerosis. Ann. Neurol. 2003, 53, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Frontiers|Modulation of Cellular Signaling by Herpesvirus-Encoded G Protein-Coupled Receptors|Pharmacology. Available online: https://www.frontiersin.org/articles/10.3389/fphar.2015.00040/full (accessed on 11 June 2020).
- Pontejo, S.M.; Murphy, P.M.; Pease, J.E. Chemokine Subversion by Human Herpesviruses. J. Innate Immun. 2018, 10, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Isegawa, Y.; Ping, Z.; Nakano, K.; Sugimoto, N.; Yamanishi, K. Human herpesvirus 6 open reading frame U12 encodes a functional beta-chemokine receptor. J. Virol. 1998, 72, 6104–6112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menotti, L.; Mirandola, P.; Locati, M.; Campadelli-Fiume, G. Trafficking to the plasma membrane of the seven-transmembrane protein encoded by human herpesvirus 6 U51 gene involves a cell-specific function present in T lymphocytes. J. Virol. 1999, 73, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, K.; Kondo, T.; Shimada, K.; Amo, K.; Miyagawa, H.; Yamanishi, K. Strong interaction between human herpesvirus 6 and peripheral blood monocytes/macrophages during acute infection. J. Med. Virol. 2002, 67, 364–369. [Google Scholar] [CrossRef]
- Zhen, Z.; Bradel-Tretheway, B.; Sumagin, S.; Bidlack, J.M.; Dewhurst, S. The human herpesvirus 6 G protein-coupled receptor homolog U51 positively regulates virus replication and enhances cell-cell fusion in vitro. J. Virol. 2005, 79, 11914–11924. [Google Scholar] [CrossRef] [Green Version]
- Milne, R.S.B.; Mattick, C.; Nicholson, L.; Devaraj, P.; Alcami, A.; Gompels, U.A. RANTES binding and down-regulation by a novel human herpesvirus-6 β chemokine receptor. J. Immunol. 2000, 164, 2396–2404. [Google Scholar] [CrossRef] [Green Version]
- Appay, V.; Rowland-Jones, S.L. RANTES: A versatile and controversial chemokine. Trends Immunol. 2001, 22, 83–87. [Google Scholar] [CrossRef]
- Vandamme, A.M.; Fransen, K.; Debaisieux, L.; Marissens, D.; Sprecher, S.; Vaira, D.; Vandenbroucke, A.T.; Verhofstede, C. Standardisation of primers and an algorithm for HIV-1 diagnostic PCR evaluated in patients harbouring strains of diverse geographical origin. The Belgian AIDS Reference Laboratories. J. Virol. Methods 1995, 51, 305–316. [Google Scholar] [CrossRef]
- Secchiero, P.; Carrigan, D.R.; Asano, Y.; Benedetti, L.; Crowley, R.W.; Komaroff, A.L.; Gallo, R.C.; Lusso, P. Detection of human herpesvirus 6 in plasma of children with primary infection and immunosuppressed patients by polymerase chain reaction. J. Infect. Dis. 1995, 171, 273–280. [Google Scholar] [CrossRef]
- Taniguchi, T.; Shimamoto, T.; Isegawa, Y.; Kondo, K.; Yamanishi, K. Structure of transcripts and proteins encoded by U79-80 of human herpesvirus 6 and its subcellular localization in infected cells. Virology 2000, 271, 307–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, D.; Liakos, V.; Michou, V.; Kapranos, N.; Kaltsas, G.; Tsilivakos, V.; Tsatsoulis, A. Detection of herpes virus DNA in post-operative thyroid tissue specimens of patients with autoimmune thyroid disease. Exp. Clin. Endocrinol. Diabetes 2008, 116, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Billstrom, M.A.; Johnson, G.L.; Avdi, N.J.; Worthen, G.S. Intracellular signaling by the chemokine receptor US28 during human cytomegalovirus infection. J. Virol. 1998, 72, 5535–5544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albert, V.; Subramanian, A.; Agrawal, D.; Bhoi, S.K.; Pallavi, P.; Mukhopadhayay, A.K. RANTES levels in peripheral blood, CSF and contused brain tissue as a marker for outcome in traumatic brain injury (TBI) patients. BMC Res. Notes 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.-M.; Wang, J.; Pan, H.-F.; Chen, G.-M.; Li, J.; Cen, H.; Feng, C.-C.; Ye, D.-Q. Increased serum RANTES in patients with systemic lupus erythematosus. Rheumatol. Int. 2012, 32, 1231–1233. [Google Scholar] [CrossRef]
- Fichna, M.; Żurawek, M.; Budny, B.; Komarowska, H.; Niechciał, E.; Fichna, P.; Ruchała, M. Elevated serum RANTES chemokine levels in autoimmune Addison disease. Pol. Arch. Intern. Med. 2018, 128, 216–221. [Google Scholar]
- Pavkova Goldbergova, M.; Lipkova, J.; Pavek, N.; Gatterova, J.; Vasku, A.; Soucek, M.; Nemec, P. RANTES, MCP-1 chemokines and factors describing rheumatoid arthritis. Mol. Immunol. 2012, 52, 273–278. [Google Scholar] [CrossRef]
- Akahane, M.; Watanabe, M.; Inoue, N.; Miyahara, Y.; Arakawa, Y.; Inoue, Y.; Katsumata, Y.; Hidaka, Y.; Iwatani, Y. Association of the polymorphisms of chemokine genes (IL8, RANTES, MIG, IP10, MCP1 and IL16) with the pathogenesis of autoimmune thyroid diseases. Autoimmunity 2016, 49, 312–319. [Google Scholar] [CrossRef]
- GarcÍa-López, M.Á.; Sancho, D.; Sánchez-Madrid, F.; Marazuela, M. Thyrocytes from autoimmune thyroid disorders produce the chemokines IP-10 and mig and attract CXCR3+ lymphocytes. J. Clin. Endocrinol. Metab. 2001, 86, 5008–5016. [Google Scholar] [CrossRef] [Green Version]
- Marfaing-Koka, A.; Devergne, O.; Gorgone, G.; Portier, A.; Schall, T.J.; Galanaud, P.; Emilie, D. Regulation of the production of the RANTES chemokine by endothelial cells. Synergistic induction by IFN-gamma plus TNF-alpha and inhibition by IL-4 and IL-13. J. Immunol. 1995, 154, 1870–1878. [Google Scholar]
- Hirsch, A.J.; Shenk, T. Human cytomegalovirus inhibits transcription of the CC chemokine MCP-1 gene. J. Virol. 1999, 73, 404–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelson, S.; Dal Monte, P.; Zipeto, D.; Bodaghi, B.; Laurent, L.; Oberlin, E.; Arenzana-Seisdedos, F.; Virelizier, J.L.; Landini, M.P. Modulation of RANTES production by human cytomegalovirus infection of fibroblasts. J. Virol. 1997, 71, 6495–6500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, M.B.; Worthen, G.S. Viral regulation of RANTES expression during human cytomegalovirus infection of endothelial cells. J. Virol. 2001, 75, 3383–3390. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultanova, A.; Cistjakovs, M.; Sokolovska, L.; Todorova, K.; Cunskis, E.; Murovska, M. HHV-6 Infection and Chemokine RANTES Signaling Pathway Disturbance in Patients with Autoimmune Thyroiditis. Viruses 2020, 12, 689. https://doi.org/10.3390/v12060689
Sultanova A, Cistjakovs M, Sokolovska L, Todorova K, Cunskis E, Murovska M. HHV-6 Infection and Chemokine RANTES Signaling Pathway Disturbance in Patients with Autoimmune Thyroiditis. Viruses. 2020; 12(6):689. https://doi.org/10.3390/v12060689
Chicago/Turabian StyleSultanova, Alina, Maksims Cistjakovs, Liba Sokolovska, Katerina Todorova, Egils Cunskis, and Modra Murovska. 2020. "HHV-6 Infection and Chemokine RANTES Signaling Pathway Disturbance in Patients with Autoimmune Thyroiditis" Viruses 12, no. 6: 689. https://doi.org/10.3390/v12060689
APA StyleSultanova, A., Cistjakovs, M., Sokolovska, L., Todorova, K., Cunskis, E., & Murovska, M. (2020). HHV-6 Infection and Chemokine RANTES Signaling Pathway Disturbance in Patients with Autoimmune Thyroiditis. Viruses, 12(6), 689. https://doi.org/10.3390/v12060689