The Emergence of a vv + MDV Can Break through the Protections Provided by the Current Vaccines

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Vaccines
2.2. Experimental Design
2.3. Sample Collection and Processing
2.4. Evaluation of Viral Loads in GX18NNM4-Challenged Birds by Real-Time PCR
2.5. Expression Analyses of the PD-1 and PD-L1 mRNA by Real-Time RT-PCR
2.6. Statistical Analysis
2.7. Ethical Statement
3. Results
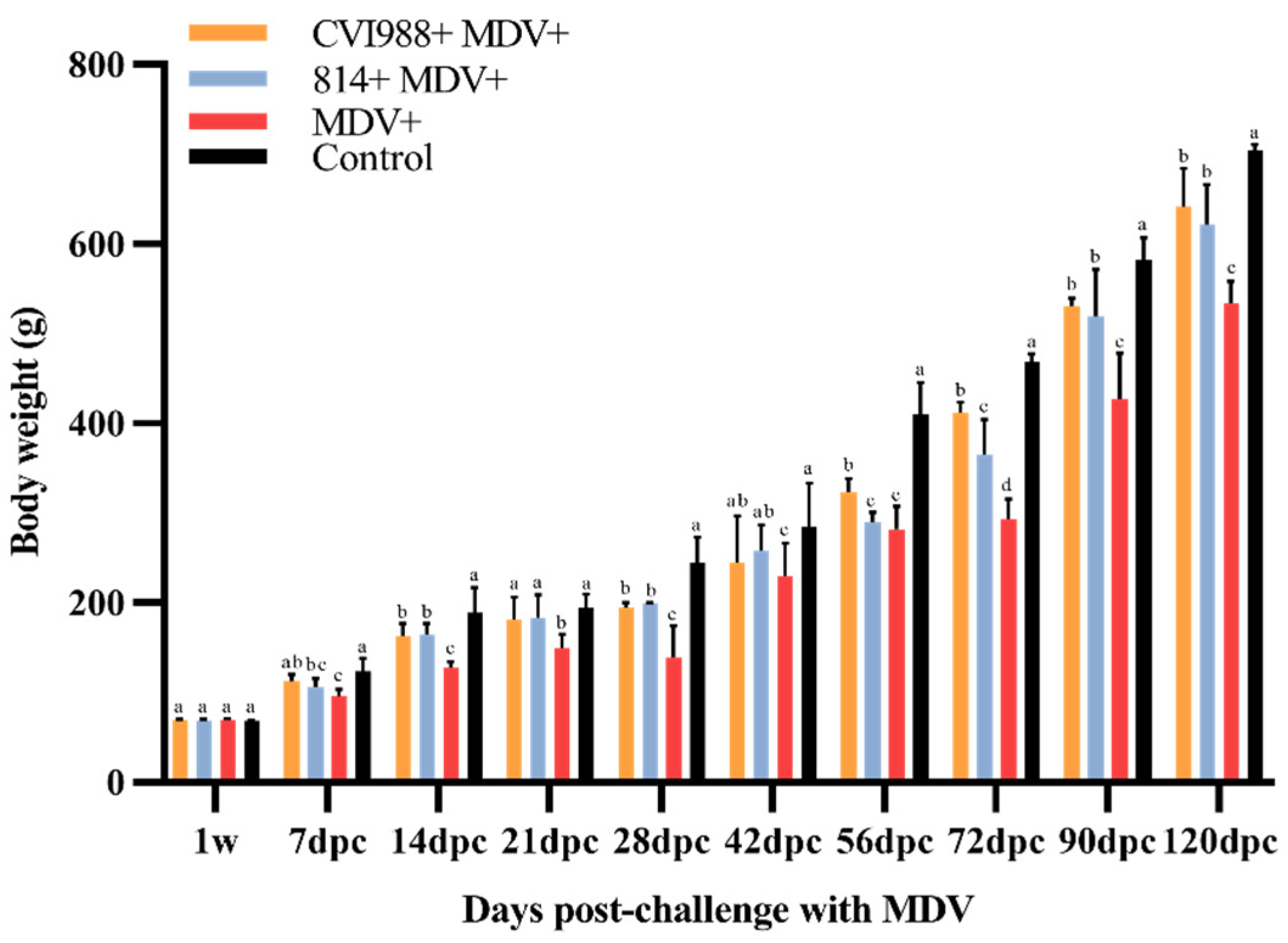
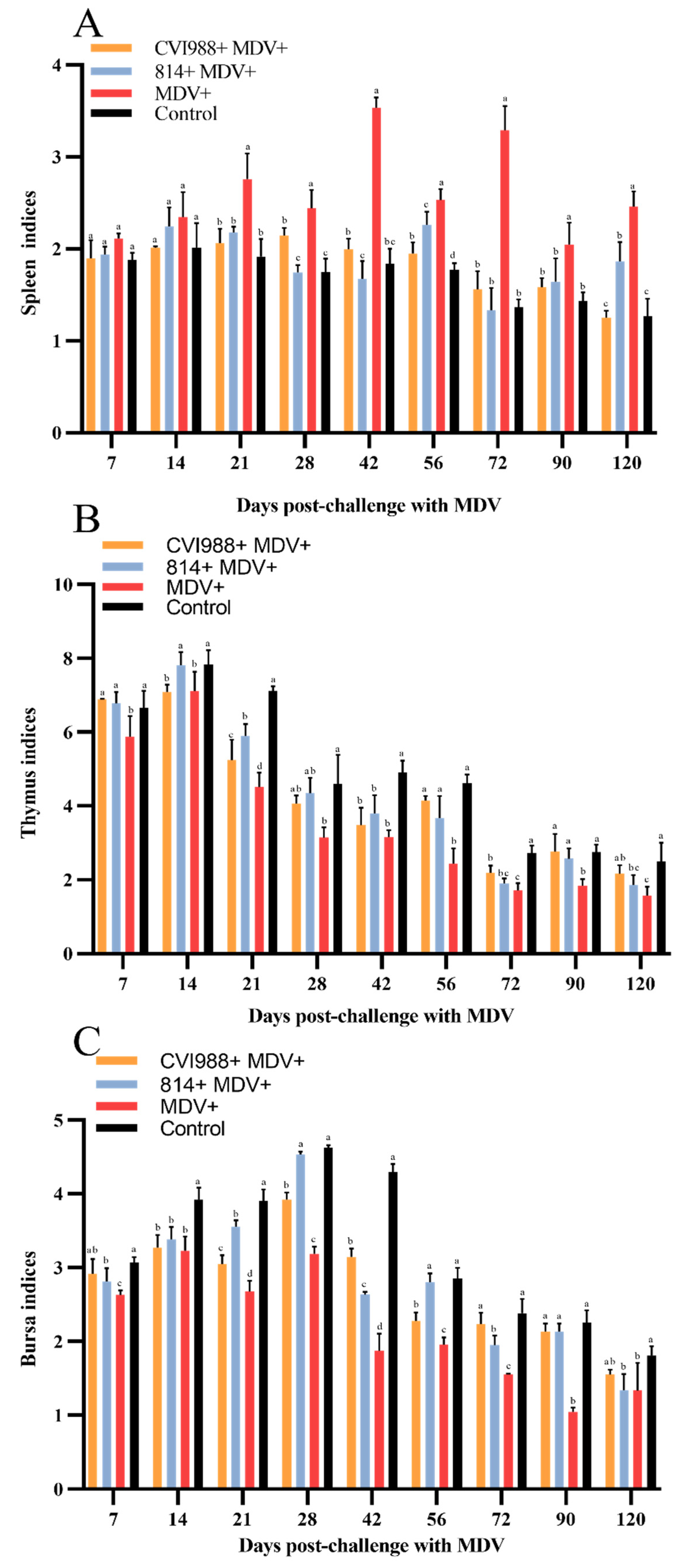
3.1. Birds Infected with GX18NNM4 Show Significantly Inhibited Body Weight Gain and Severe Damage to the Immune Organs
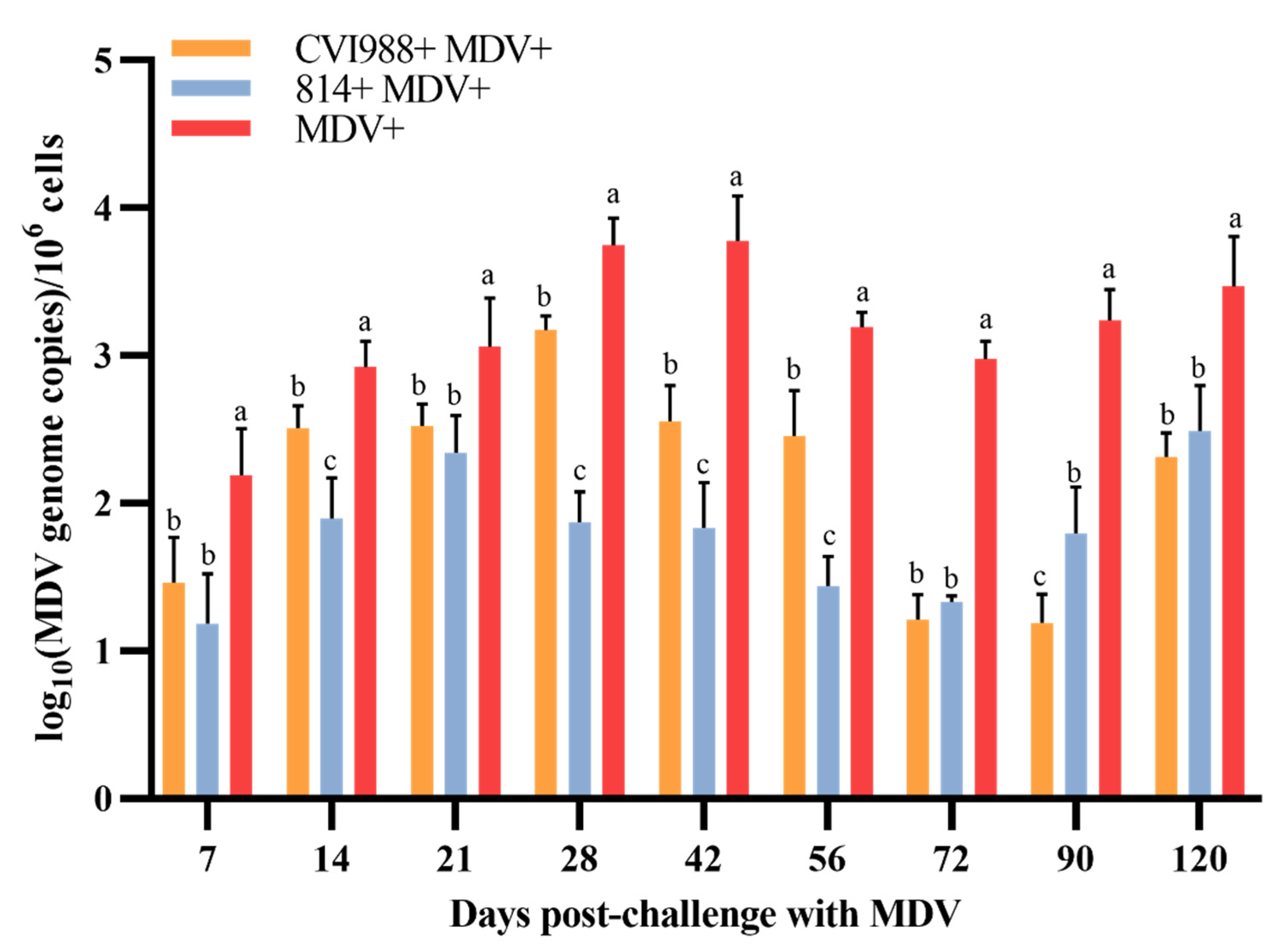
3.2. The Viral Load in PBLs Was the Highest in Unvaccinated Challenged Birds
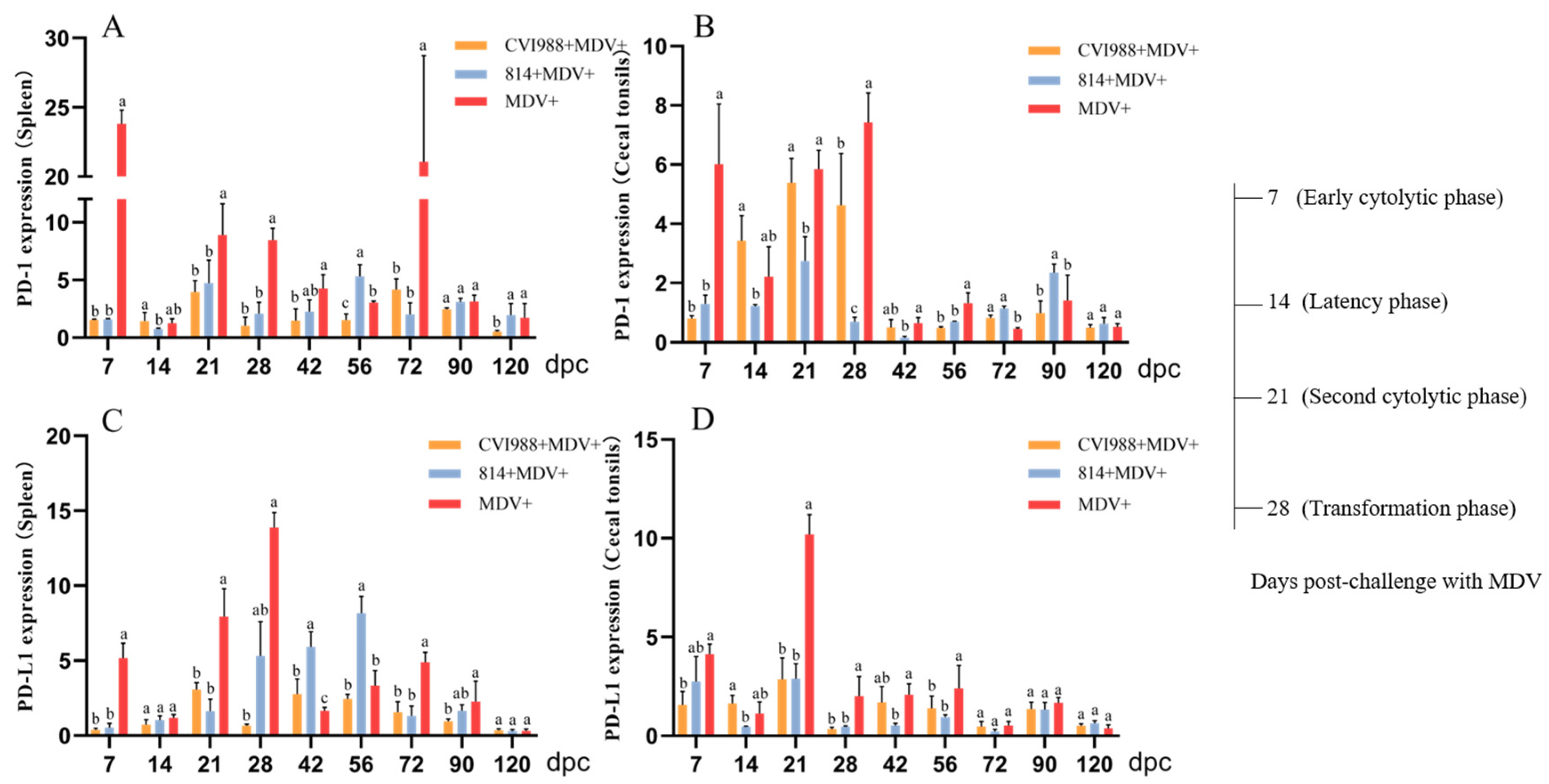
3.3. The Expression of PD-1 and PD-L1 Was Significantly Increased in the Unvaccinated Challenged Birds
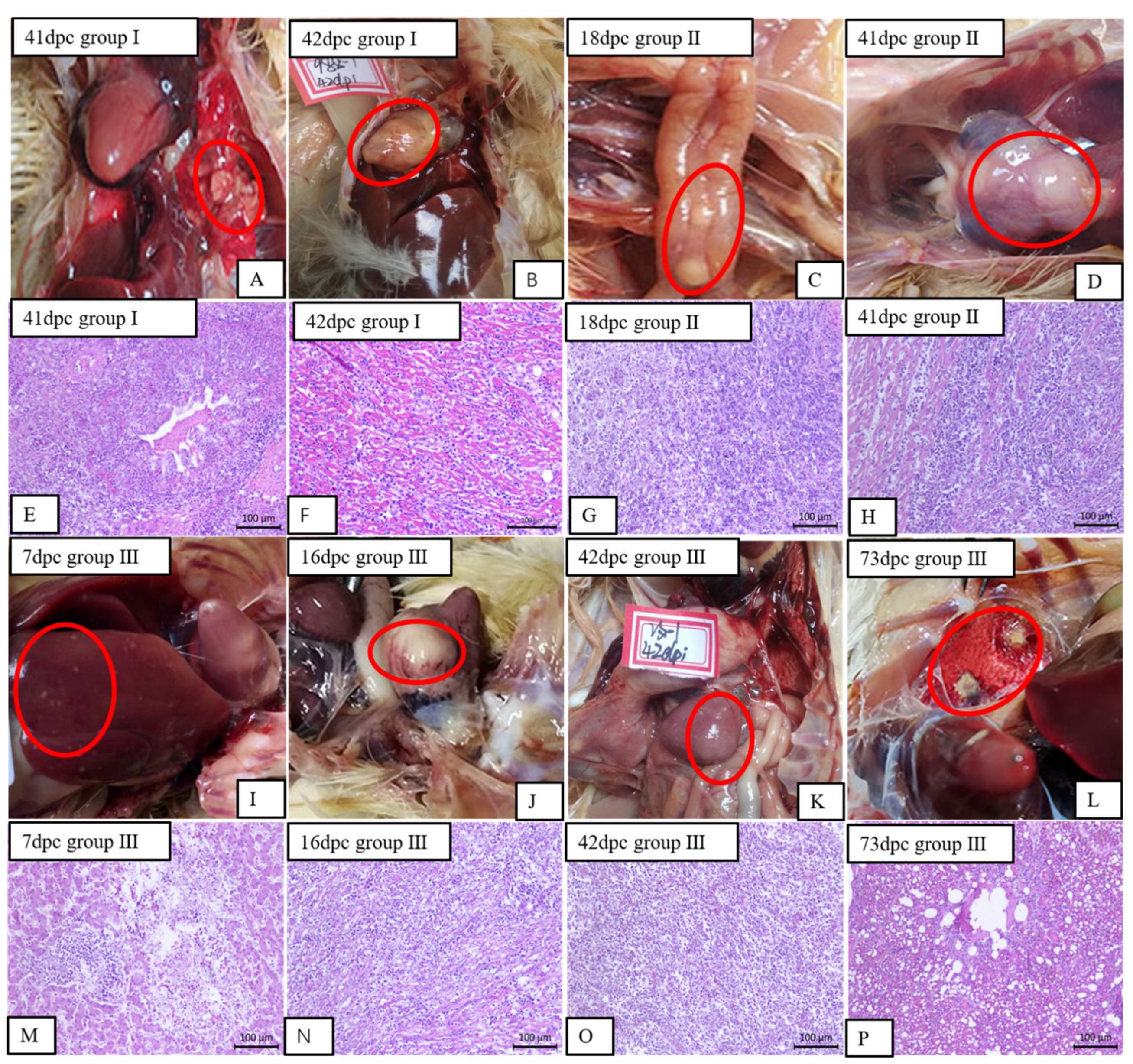
3.4. The Gross Lesions and Histopathological Observations of the Experimentally Infected Birds
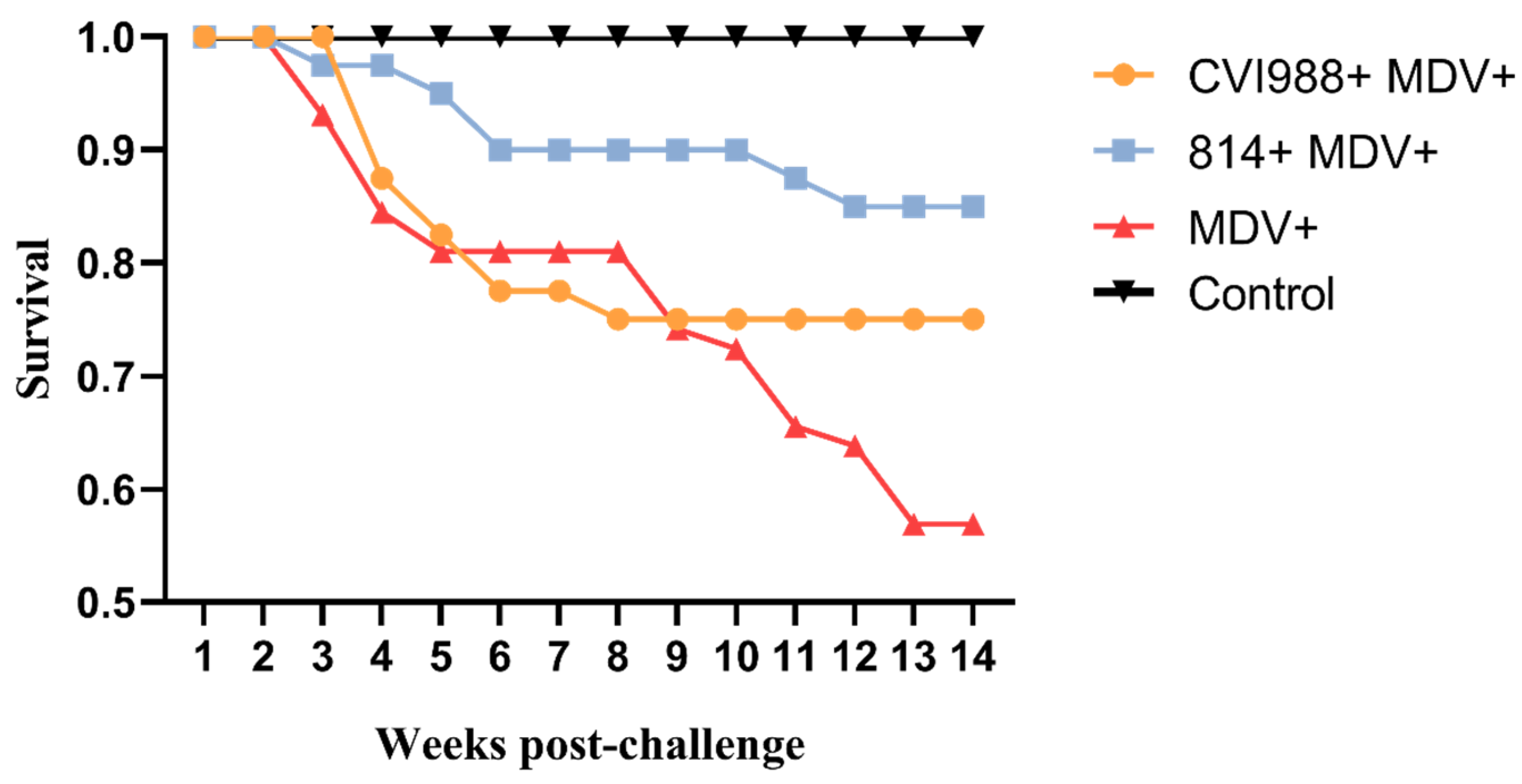
3.5. Birds Infected with GX18NNM4 Can Exhibit Severe Tumor Incidence and Break through the Protections Provided by the Current Vaccines
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nolan, L.K.; Swayne, D.E.; Saif, Y.M.; Fadly, A.M.; Glisson, J.R.; Mcdougald, L.R. Diseases of Poultry; Wiley: Hoboken, NJ, USA, 2016; pp. 513–673. [Google Scholar]
- Biggs, P.M. The History and Biology of Marek’s Disease Virus. Curr. Top. Microbiol. Immunol. 2001, 255, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Osterrieder, N.; Kamil, J.P.; Schumacher, D.; Tischer, B.K.; Trapp, S. Marek’s disease virus: From miasma to model. Nat. Rev. Microbiol. 2006, 4, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Bertzbach, L.D.; Conradie, A.M.; You, Y.; Kaufer, B.B. Latest Insights into Marek’s Disease Virus Pathogenesis and Tumorigenesis. Cancers 2020, 12, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, C.; Fehler, F. 5-Marek’s disease: A worldwide problem. In Marek’s Disease; Davison, F., Nair, V., Eds.; Academic Press: Oxford, UK, 2004; pp. 49–61. [Google Scholar]
- Baigent, S.; Smith, L.P.; Nair, V.; Currie, R.J. Vaccinal control of Marek’s disease: Current challenges, and future strategies to maximize protection. Vet. Immunol. Immunopathol. 2006, 112, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Schat, K.A.; Calnek, B.W. Characterization of an Apparently Nononcogenic Marek’s Disease Virus. J. Natl. Cancer Inst. 1978, 60, 1075–1082. [Google Scholar] [CrossRef]
- Cui, H.; Gao, H.; Cui, X.; Zhao, Y.; Shi, X.; Li, Q.; Yan, S.; Gao, M.; Wang, M.; Liu, C.; et al. Avirulent Marek’s Disease Virus Type 1 Strain 814 Vectored Vaccine Expressing Avian Influenza (AI) Virus H5 Haemagglutinin Induced Better Protection Than Turkey Herpesvirus Vectored AI Vaccine. PLoS ONE 2013, 8, e53340. [Google Scholar] [CrossRef] [Green Version]
- Witter, R.L.; Kreager, K.S. Serotype 1 Viruses Modified by Backpassage or Insertional Mutagenesis: Approaching the Threshold of Vaccine Efficacy in Marek’s Disease. Avian Dis. 2004, 48, 768–782. [Google Scholar] [CrossRef]
- Nair, V. Successful control of Marek’s disease by vaccination. Dev. Biol. 2004, 119, 147–154. [Google Scholar]
- Witter, R.L. Protective Efficacy of Marek’s Disease Vaccines. Curr. Top. Microbiol. Immunol. 2001, 255, 57–90. [Google Scholar] [CrossRef]
- Witter, R.L. Increased Virulence of Marek’s Disease Virus Field Isolates. Avian Dis. 1997, 41, 149. [Google Scholar] [CrossRef]
- Witter, R.L.; Calnek, B.W.; Buscaglia, C.; Gimeno, I.M.; Schat, K.A. Classification of Marek’s disease viruses according to pathotype: Philosophy and methodology. Avian Pathol. 2005, 34, 75–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calnek, B.W.; Witter, R.L. Marek’s Disease—A Model for Herpesvirus Oncology. Crit. Rev. Microbiol. 1985, 12, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Shek, W.R.; Calnek, B.W.; A Schat, K.; Chen, C.H. Characterization of Marek’s disease virus-infected lymphocytes: Discrimination between cytolytically and latently infected cells. J. Natl. Cancer Inst. 1983, 70, 485–491. [Google Scholar] [PubMed]
- Calnek, B.W. Pathogenesis of Marek’s Disease Virus Infection. Curr. Top. Microbiol. Immunol. 2001, 255, 25–55. [Google Scholar] [PubMed]
- Freeman, G.J.; Long, A.J.; Iwai, Y.; Bourque, K.; Chernova, T.; Nishimura, H.; Fitz, L.J.; Malenkovich, N.; Okazaki, T.; Byrne, M.C.; et al. Engagement of the Pd-1 Immunoinhibitory Receptor by a Novel B7 Family Member Leads to Negative Regulation of Lymphocyte Activation. J. Exp. Med. 2000, 192, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Freeman, G.J.; Wherry, E.J.; Ahmed, R.; Sharpe, A.H. Reinvigorating exhausted HIV-specific T cells via PD-1–PD-1 ligand blockade. J. Exp. Med. 2006, 203, 2223–2227. [Google Scholar] [CrossRef] [Green Version]
- Greenwald, R.J.; Freeman, G.J.; Sharpe, A. The B7 Family Revisited. Annu. Rev. Immunol. 2005, 23, 515–548. [Google Scholar] [CrossRef]
- Reddy, V.; Mwangi, W.; Sadigh, Y.; Nair, V. In vitro Interactions of Chicken Programmed Cell Death 1 (PD-1) and PD-1 Ligand-1 (PD-L1). Front. Cell. Infect. Microbiol. 2019, 9, 436. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, P.; Lin, L.; Shi, M.; Gu, Z.; Huang, T.; Mo, M.-L.; Wei, T.; Zhang, H.; Wei, P.; et al. The emergence of the infection of subgroup J avian leucosis virus escalated the tumour incidence in commercial Yellow chickens in Southern China in recent years. Transbound. Emerg. Dis. 2018, 66, 312–316. [Google Scholar] [CrossRef] [Green Version]
- Teng, L.-Q.; Wei, P.; Song, Z.-B.; Yang, N.-L. Evaluation of the pathogenicity of a field isolate of Marek’s disease virus integrated with retroviral long terminal repeat sequence. Chin. J. Virol. 2009, 25, 376–381. [Google Scholar]
- Buscaglia, C. Mixed Infections of Marek’s Disease and Reticuloendotheliosis Viruses in Layer Flocks in Argentina. Avian Dis. 2013, 57, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Consigli, R.A.; Zabielski, J.; Weil, R. Plaque Assay for Polyoma Virus on Primary Mouse Kidney Cell Cultures. Appl. Microbiol. 1973, 26, 627–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, P.; Cui, Z. The Comparison of meq Gene Sequences Amplified from Different Pathotypes of Mared′s Disease Virus. Chin. J. Prev. Vet. Med. 2002, 24, 9–13. [Google Scholar]
- Lin, L.; Li, H.; Wei, P.; Wang, P.; Yang, Y.; Huang, T. Full-length genome sequence analysis of four subgroup J avian leukosis virus strains isolated from chickens with clinical hemangioma. Virus Genes 2017, 53, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.-R.; Zhang, Y.; Zhou, L.; Lv, H.-C.; Zhang, F.; Li, K.; Gao, Y.; Qi, X.; Cui, H.-Y.; Wang, Y.; et al. Co-Infection with Marek’s Disease Virus and Reticuloendotheliosis Virus Increases Illness Severity and Reduces Marek’s Disease Vaccine Efficacy. Viruses 2017, 9, 158. [Google Scholar] [CrossRef]
- Wang, P.; Lin, L.; Shi, M.; Li, H.; Gu, Z.; Li, M.; Gao, Y.; Teng, H.; Mo, M.; Wei, T.; et al. Vertical transmission of ALV from ALV-J positive parents caused severe immunosuppression and significantly reduced marek’s disease vaccine efficacy in three-yellow chickens. Vet. Microbiol. 2020, 244, 108683. [Google Scholar] [CrossRef] [PubMed]
- Del Moral, M.G.; Fonfría, J.; Varas, A.; Jimenez, E.; Moreno, J.; Zapata, A.G. Appearance and development of lymphoid cells in the chicken (Gallus gallus) caecal tonsil. Anat. Rec. 1998, 250, 182–189. [Google Scholar] [CrossRef]
- Heidari, M.; Fitzgerald, S.D.; Zhang, H. Immune Responses in Cecal Tonsils of Marek’s Disease Virus–Infected Chickens. Avian Dis. 2015, 59, 213–226. [Google Scholar] [CrossRef]
- Matsuyama-Kato, A.; Murata, S.; Isezaki, M.; Kano, R.; Takasaki, S.; Ichii, O.; Konnai, S.; Ohashi, K. Molecular characterization of immunoinhibitory factors PD-1/PD-L1 in chickens infected with Marek’s disease virus. J. Virol. 2012, 9, 94. [Google Scholar] [CrossRef] [Green Version]
- Gurung, A.; Kamble, N.M.; Kaufer, B.B.; Pathan, A.; Behboudi, S. Association of Marek’s Disease induced immunosuppression with activation of a novel regulatory T cells in chickens. PLoS Pathog. 2017, 13, e1006745. [Google Scholar] [CrossRef] [Green Version]
- López-Osorio, S.; Piedrahita, D.; Espinal-Restrepo, M.A.; Nieto, G.R.; Nair, V.; Williams, S.M.; Baigent, S.; Ventura-Polite, C.; Aranzazu-Taborda, D.A.; Chaparro-Gutiérrez, J.J. Molecular characterization of Marek’s disease virus in a poultry layer farm from Colombia. Poult. Sci. 2017, 96, 1598–1608. [Google Scholar] [CrossRef] [PubMed]
- Teng, L.-Q.; Wei, P.; Song, Z.-B.; He, J.-J.; Cui, Z.-Z. Molecular epidemiological investigation of Marek’s disease virus from Guangxi, China. Arch. Virol. 2011, 156, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, Z.; Lan, X.; Zhang, F.; Wang, Q.; Li, K.; Pan, Q.; Gao, Y.; Qi, X.; Cui, H.-Y.; et al. A high frequency of Gallid herpesvirus-2 co-infection with Reticuloendotheliosis virusis associated with high tumor rates in Chinese chicken farms. Vet. Microbiol. 2019, 237, 108418. [Google Scholar] [CrossRef]
- Wei, P. Challenge and Development Opportunity of Quality Chicken Industry in China: A Review about 9 Issues on the Topic. China Poult. 2019, 41, 1–6. [Google Scholar]
- Witter, R.L. Control strategies for Marek’s disease: A perspective for the future. Poult. Sci. 1998, 77, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-P.; Li, Z.-J.; Bao, K.-Y.; Lv, H.-C.; Gao, Y.; Gao, H.-L.; Qi, X.; Cui, H.; Wang, Y.-Q.; Ren, X.; et al. Pathogenic characteristics of Marek’s disease virus field strains prevalent in China and the effectiveness of existing vaccines against them. Vet. Microbiol. 2015, 177, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Zhao, Z.; Wei, P.; Wei, X.; Li, Y.; Mo, M. Isolation and Identification of a Field Isolate of Marek’s Disease Virus with Acute Oncogenicity. Chin. J. Virol. 2007, 23, 218–223. [Google Scholar]
- Wang, P.; Shi, M.; He, C.; Lin, L.; Li, H.; Gu, Z.; Li, M.; Gao, Y.; Huang, T.; Mo, M.; et al. A novel recombinant avian leukosis virus isolated from gamecocks induced pathogenicity in Three-Yellow chickens: A potential infection source of avian leukosis virus to the commercial chickens. Poult. Sci. 2019, 98, 6497–6504. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | No. of Birds | Vaccine (Day 1) | MDV Challenge (Day 7) | PFU per Volume (μL) |
|---|---|---|---|---|
| I | 40 | CVI988/Rispens | GX18NNM4 | 1000 per 200 |
| II | 40 | 814 | GX18NNM4 | 1000 per 200 |
| III | 58 | DMEM | GX18NNM4 | 1000 per 200 |
| IV | 37 | DMEM | DMEM | 0 per 200 |
| Gene | Type | Sequence (5′-3′) | Accession Number | Reference |
|---|---|---|---|---|
| pp38 | Forward | ATAAAGGGTGATGGGAAGGC | Our group’s designs (unpublished) | |
| Reverse | CGTCAAGATGTTCATTCCCTG | |||
| Probe | TCCTCCCACTGTGACAGCC | |||
| PD-1 | Forward | GGACTACGGTGTGCTGGAGTT | XM422723 | Matsuyama-Kato et al., 2012 |
| Reverse | TCTTTCCTCGCTCTGGTGTG | |||
| PD-L1 | Forward | TTCAGGGACGGATAAAGCTG | XM424811 | |
| Reverse | CGTCTCTGAGCTTCACGTTG | |||
| β-actin | Forward | GAGAAATTGTGCGTGACATCA | NM205518 | Our group’s designs (unpublished) |
| Reverse | CCTGAACCTCTCATTGCCA |
| Group | Heart | Liver | Spleen | Lung | Pancreas | Caecum | Morbidity | Mortality | Tumor Rate | PI |
|---|---|---|---|---|---|---|---|---|---|---|
| I | 22.5% (9/40) | 12.5% (5/40) | 7.5% (3/40) | 5% (2/40) | 20% (8/40) | 17.5% (7/40) | 45% (18/40) | 25% (10/40) | 42.5% (17/40) | 39.9% |
| II | 15% (6/40) | 7.5% (3/40) | 5% (2/40) | 0 | 7.5% (3/40) | 10% (4/40) | 27.5% (11/40) | 15% (6/40) | 27.5% (11/40) | 61.1% |
| III | 34.5% (20/58) | 32.8% (19/58) | 17.2% (10/58) | 13.8% (8/58) | 27.6% (16/58) | 17.2% (10/58) | 75.9% (44/58) | 43.1% (25/58) | 70.7% (41/58) | -- |
| IV | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | -- |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, M.-y.; Li, M.; Wang, W.-w.; Deng, Q.-m.; Li, Q.-h.; Gao, Y.-l.; Wang, P.-k.; Huang, T.; Wei, P. The Emergence of a vv + MDV Can Break through the Protections Provided by the Current Vaccines. Viruses 2020, 12, 1048. https://doi.org/10.3390/v12091048
Shi M-y, Li M, Wang W-w, Deng Q-m, Li Q-h, Gao Y-l, Wang P-k, Huang T, Wei P. The Emergence of a vv + MDV Can Break through the Protections Provided by the Current Vaccines. Viruses. 2020; 12(9):1048. https://doi.org/10.3390/v12091048
Chicago/Turabian StyleShi, Meng-ya, Min Li, Wei-wei Wang, Qiao-mu Deng, Qiu-hong Li, Yan-li Gao, Pei-kun Wang, Teng Huang, and Ping Wei. 2020. "The Emergence of a vv + MDV Can Break through the Protections Provided by the Current Vaccines" Viruses 12, no. 9: 1048. https://doi.org/10.3390/v12091048
APA StyleShi, M. -y., Li, M., Wang, W. -w., Deng, Q. -m., Li, Q. -h., Gao, Y. -l., Wang, P. -k., Huang, T., & Wei, P. (2020). The Emergence of a vv + MDV Can Break through the Protections Provided by the Current Vaccines. Viruses, 12(9), 1048. https://doi.org/10.3390/v12091048