
Isolation and Characterization of Salmonella Jumbo-Phage pSal-SNUABM-04


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains Used in This Study
2.2. Isolation and Characterization of the Phage pSal-SNUABM-04
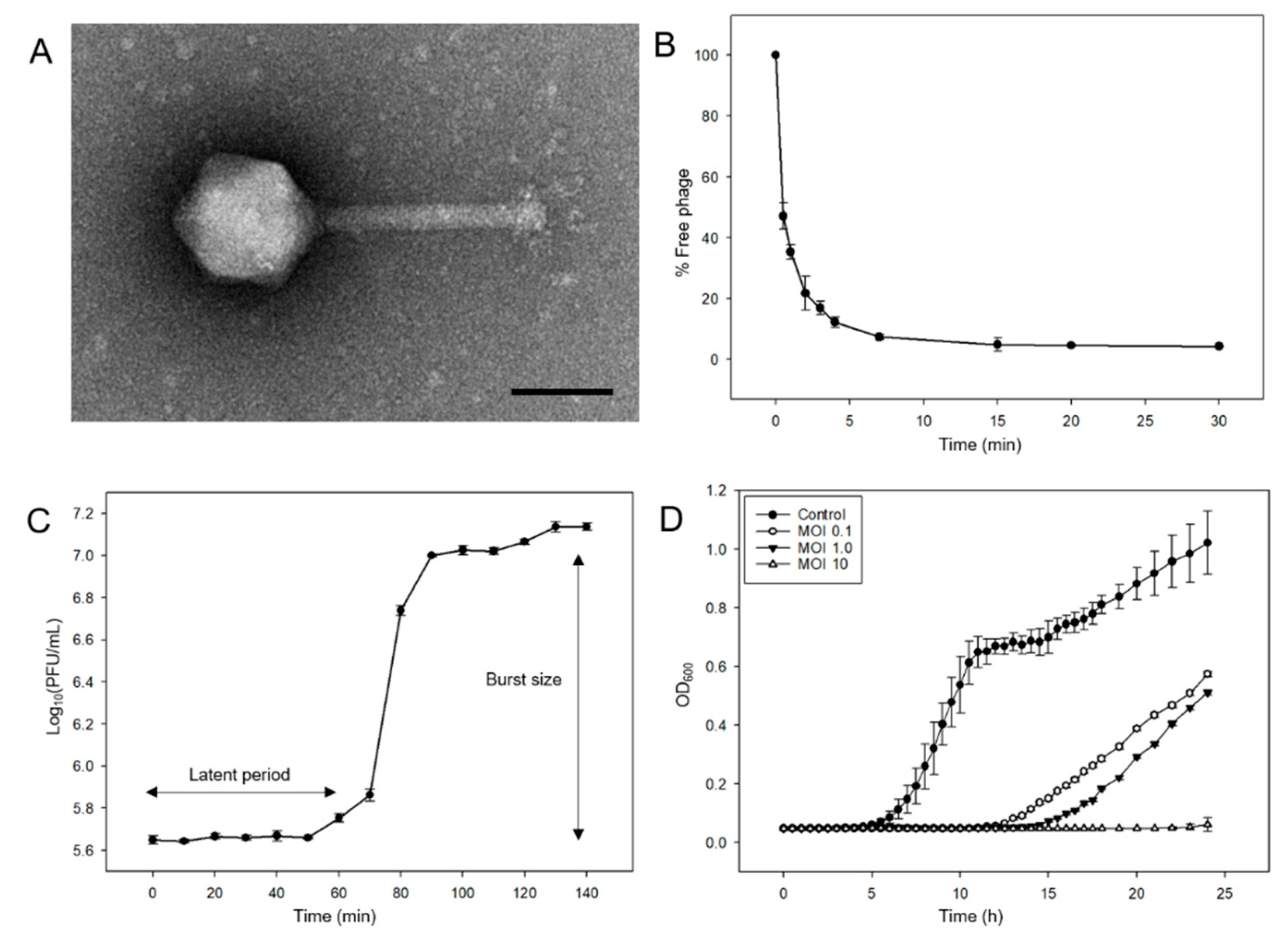
2.3. Electron Microscopy of the Phage pSal-SNUABM-04
2.4. In Vitro Planktonic Bacterial Lysis Assay
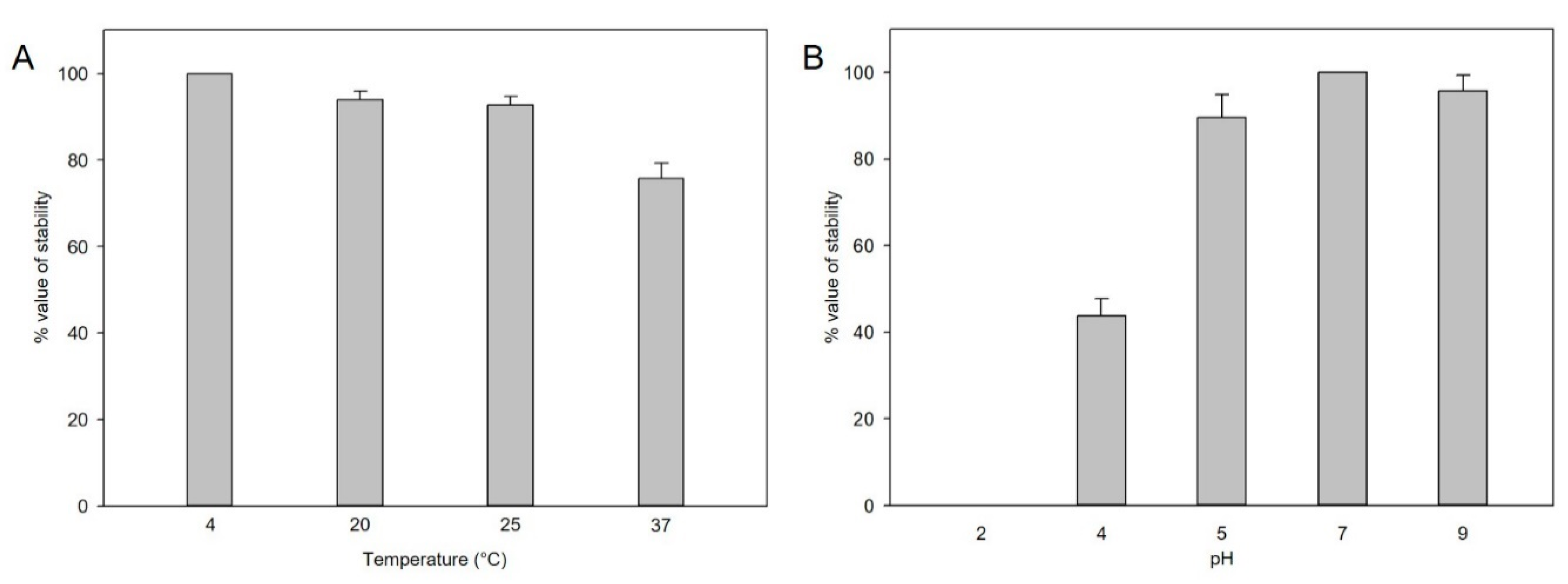
2.5. Evaluation of Phage Stability under Different Thermal and pH Conditions
2.6. Phage Sequencing and Genome Analysis
2.7. Comparative Genome Analysis
3. Results and discussions
3.1. Bacterial Strains Used in This Study
3.2. Biological Features of the Phage pSal-SNUABM-04
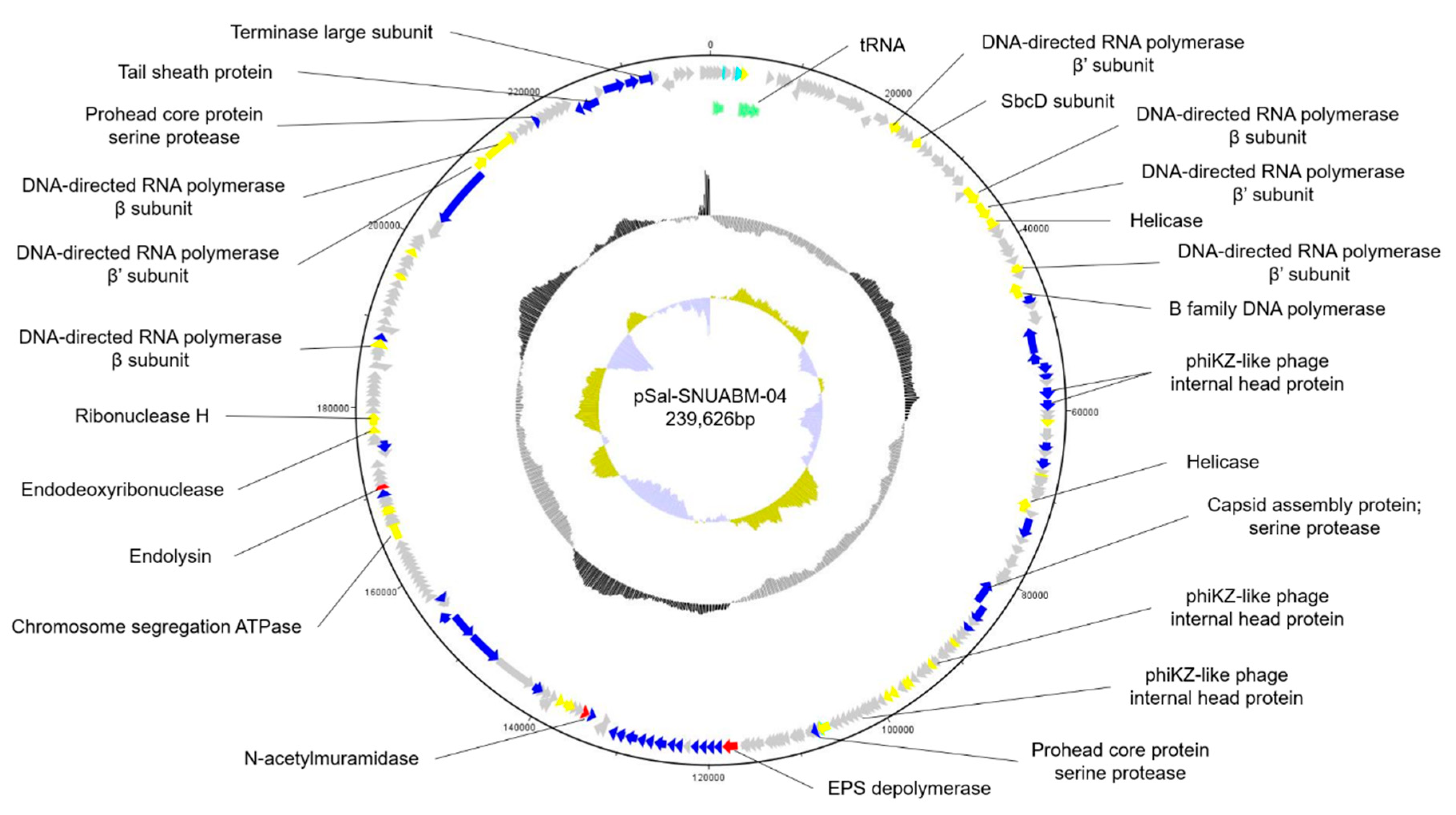
3.3. General Features of the Phage pSal-SNUABM-04 Genome
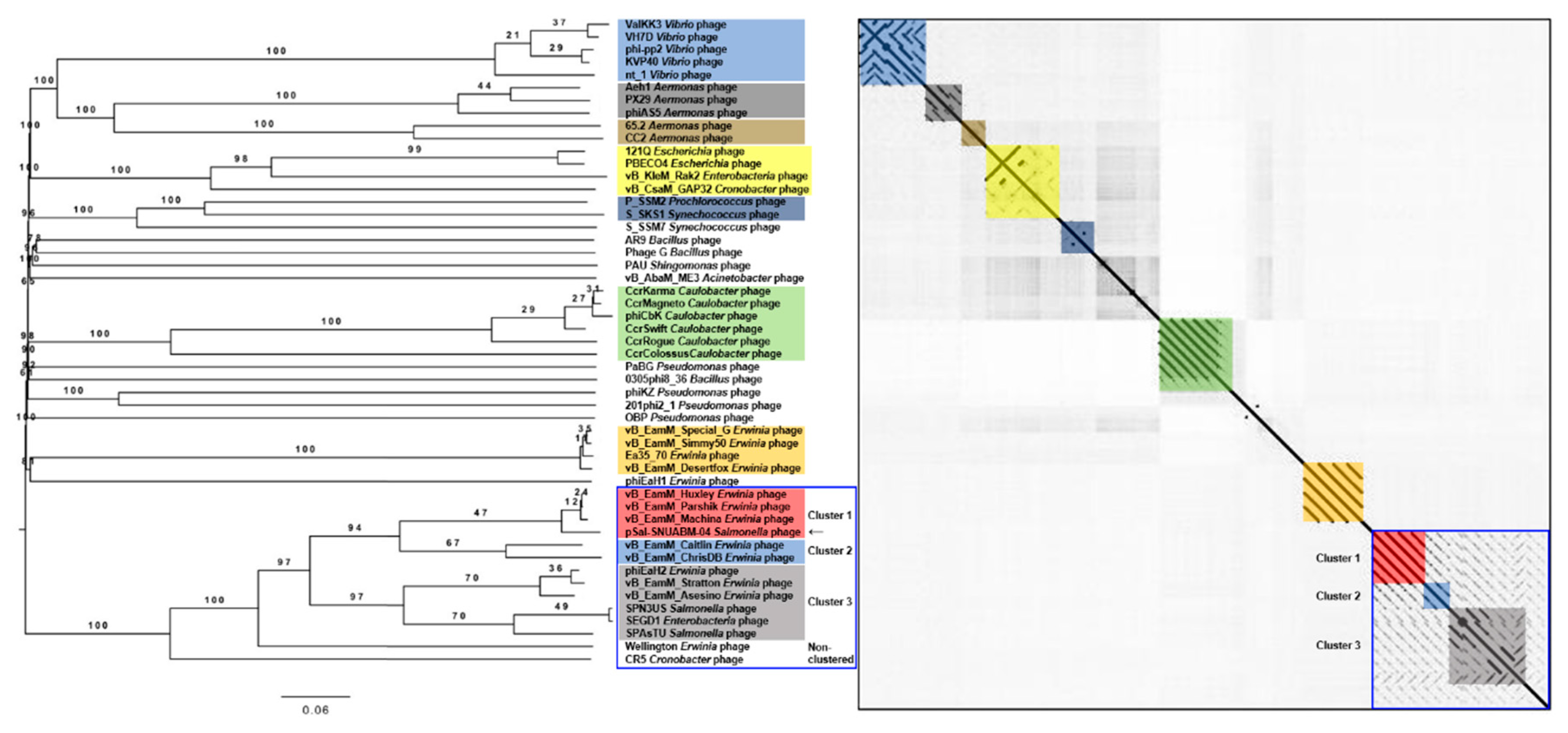
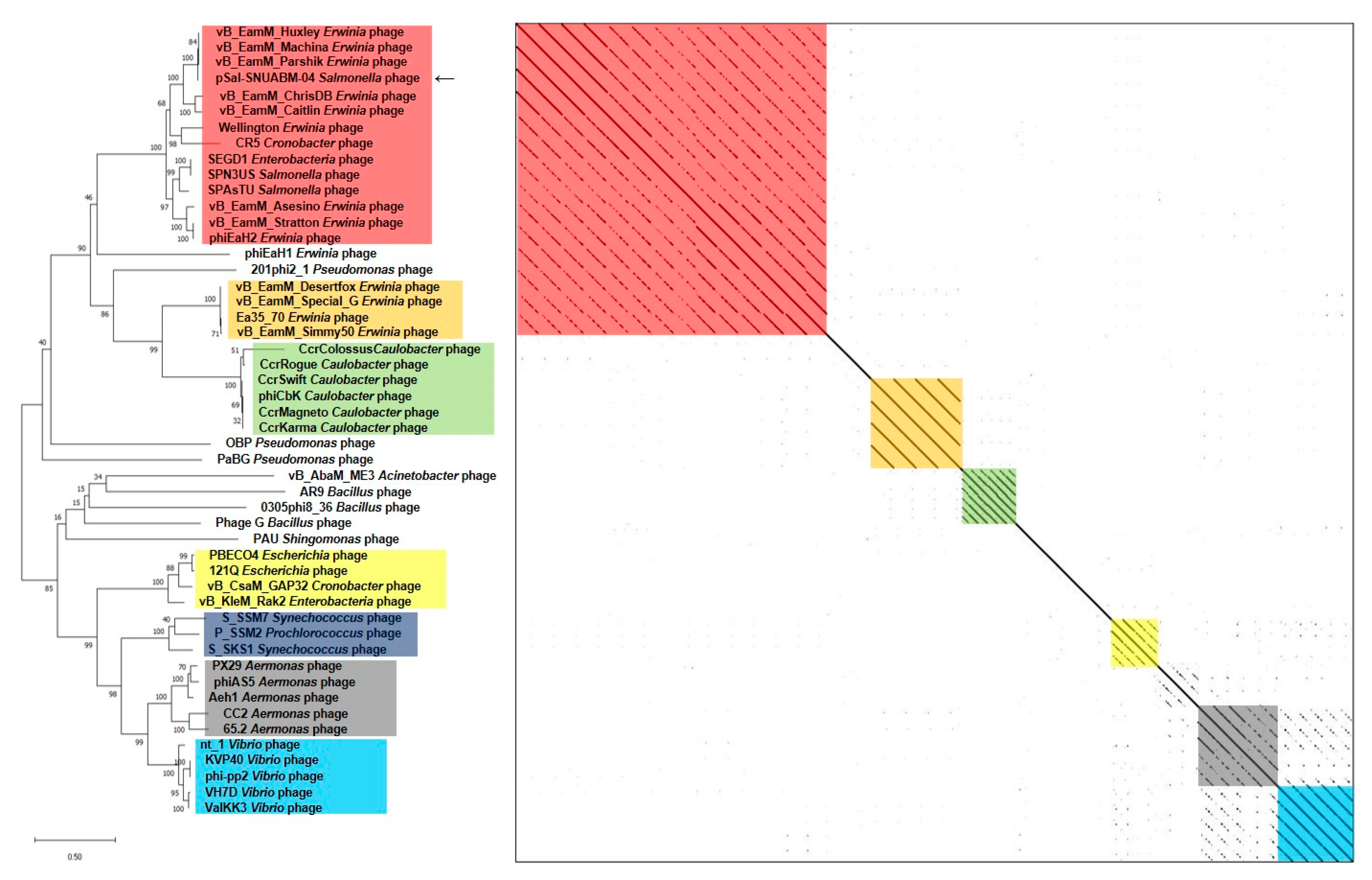
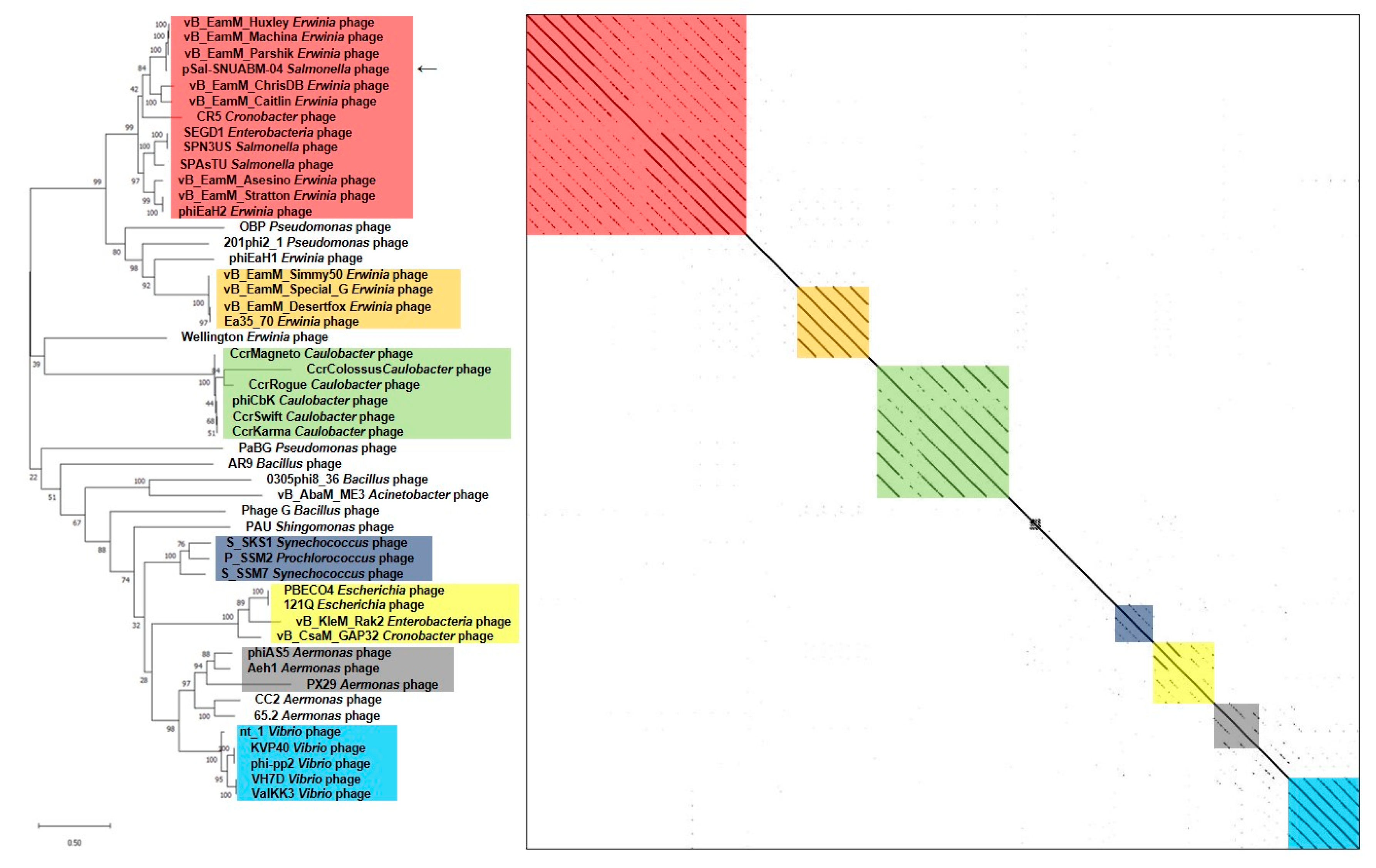
3.4. Comparative Genomics of the pSal-SNUABM-04 Genome
3.5. Specific Features of the pSal-SNUABM-04 Genome
3.5.1. Nucleotide Metabolism-Related Genes
3.5.2. Structural and Packaging Related Genes
3.5.3. Lysis-Related Genes
3.5.4. Additional Function Genes
3.5.5. Horizontal Gene Transfer
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, S.G.; Jun, J.W.; Giri, S.S.; Yun, S.; Kim, H.J.; Kang, J.W.; Han, S.J.; Jeong, D.; Park, S.C. Isolation and characterisation of pVa-21, a giant bacteriophage with anti-biofilm potential against Vibrio alginolyticus. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Kutateladze, M.; Adamia, R. Bacteriophages as potential new therapeutics to replace or supplement antibiotics. Trends Biotechnol. 2010, 28, 591–595. [Google Scholar] [CrossRef]
- Yuan, Y.; Gao, M. Jumbo bacteriophages: An overview. Front. Microbiol. 2017, 8, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Pielstick, B.A.; Bell, K.A.; Nieman, T.B.; Stubbs, O.A.; Yeates, E.L.; Baltrus, D.A.; Grose, J.H. A Novel, Highly Related Jumbo Family of Bacteriophages That Were Isolated Against Erwinia. Front. Microbiol. 2019, 10, 1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olszak, T.; Danis-Wlodarczyk, K.; Arabski, M.; Guła, G.; Maciejewska, B.; Wasik, S.; Lood, C.; Higgins, G.; Harvey, B.J.; Lavigne, R.; et al. Pseudomonas aeruginosa PA5oct Jumbo Phage Impacts Planktonic and Biofilm Population and Reduces Its Host Virulence. Viruses 2019, 11, 1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farris, J.S. Estimating Phylogenetic Trees from Distance Matrices. Am. Nat. 1972, 106, 645–668. [Google Scholar] [CrossRef]
- FigTree 1.4.3—A graphical viewer of phylogenetic trees and a program for producing publication-ready figures. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 11 November 2020).
- Krumsiek, J.; Arnold, R.; Rattei, T. Gepard: A rapid and sensitive tool for creating dot plots on genome scale. Bioinformatics 2007, 23, 1026–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.-H.; Ha, S.-M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple Alignment of Conserved Genomic Sequence with Rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, H.-W. 5500 Phages examined in the electron microscope. Arch. Virol. 2006, 152, 227–243. [Google Scholar] [CrossRef]
- Hyman, P.; Abedon, S.T. Practical Methods for Determining Phage Growth Parameters. In Bioinformatics in MicroRNA Research; Springer Science and Business Media LLC: New York, NY, USA, 2009; Volume 501, pp. 175–202. [Google Scholar]
- Thomas, J.A.; Quintana, A.D.B.; Bosch, M.A.; De Peña, A.C.; Aguilera, E.; Coulibaly, A.; Wu, W.; Osier, M.V.; Hudson, A.O.; Weintraub, S.T.; et al. Identification of Essential Genes in the Salmonella Phage SPN3US Reveals Novel Insights into Giant Phage Head Structure and Assembly. J. Virol. 2016, 90, 10284–10298. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.; Clooney, A.G.; Stockdale, S.R.; Buttimer, C.; Draper, L.A.; Ross, R.P.; Hill, C. Isolation of a Novel Jumbo Bacteriophage Effective Against Klebsiella aerogenes. Front. Med. 2020, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Ceyssens, P.J.; Minakhin, L.; Van den Bossche, A.; Yakunina, M.; Klimuk, E.; Blasdel, B.; De Smet, J.; Noben, J.P.; Bläsi, U.; Severinov, K.; et al. Development of giant bacteriophage ϕKZ is independent of the host transcription apparatus. J. Virol. 2014, 88, 10501–10510. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.S.; Kutter, E.; Mosig, G.; Arisaka, F.; Kunisawa, T.; Rüger, W. Bacteriophage T4 Genome. Microbiol. Mol. Biol. Rev. 2003, 67, 86–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storvik, K.A.M.; Foster, P.L. The SMC-Like Protein Complex SbcCD Enhances DNA Polymerase IV-Dependent Spontaneous Mutation in Escherichia coli. J. Bacteriol. 2010, 193, 660–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, J.A.; Weintraub, S.T.; Wu, W.; Winkler, D.C.; Cheng, N.; Steven, A.C.; Black, L.W. Extensive proteolysis of head and inner body proteins by a morphogenetic protease in the giant Pseudomonas aeruginosa phage φKZ. Mol. Microbiol. 2012, 84, 324–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Thomas, J.A.; Cheng, N.; Black, L.W.; Steven, A.C. Bubblegrams reveal the inner body structure of ϕKZ. Science 2012, 335, 10–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, L.W.; Thomas, J.A. Condensed Genome Structure. In Advances in Experimental Medicine and Biology; Springer Science and Business Media LLC: New York, NY, USA, 2011; Volume 726, pp. 469–487. [Google Scholar]
- Krylov, V.N.; Smirnova, T.A.; Minenkova, I.B.; Plotnikova, T.G.; Zhazikov, I.Z.; Khrenova, E.A. Pseudomonas bacteriophage contains an inner body in its capsid. Can. J. Microbiol. 1984, 30, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Desmond, M.I.; Mallory, S.A.; Benítez, A.D.; Buckley, L.J.; Weintraub, S.T.; Osier, M.V.; Black, L.W.; Thomas, J.A. To Be or Not to Be T4: Evidence of a Complex Evolutionary Pathway of Head Structure and Assembly in Giant Salmonella Virus SPN3US. Front. Microbiol. 2017, 8, 2251. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.A.; Black, L.W. Mutational analysis of the Pseudomonas aeruginosa myovirus ϕKZ morphogenetic protease gp175. J. Virol. 2013, 87, 8713–8725. [Google Scholar] [CrossRef] [Green Version]
- Young, R. Bacteriophage lysis: Mechanism and regulation. Microbiol. Mol. Biol. Rev. 1992, 56, 430–481. [Google Scholar] [CrossRef]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Futur. Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [Green Version]
- Matamp, N.; Bhat, S.G. Phage Endolysins as Potential Antimicrobials against Multidrug Resistant Vibrio alginolyticus and Vibrio parahaemolyticus: Current Status of Research and Challenges Ahead. Microorganisms 2019, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Tišáková, L.; Godány, A. Bacteriophage endolysins and their use in biotechnological processes. J. Microbiol. Biotechnol. Food Sci. 2020, 9, 164–170. [Google Scholar]
- Latka, A.; Maciejewska, B.; Majkowska-Skrobek, G.; Briers, Y.; Drulis-Kawa, Z. Bacteriophage-encoded virion-associated enzymes to overcome the carbohydrate barriers during the infection process. Appl. Microbiol. Biotechnol. 2017, 101, 3103–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, D.; Briers, Y.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; Lavigne, R.; García, P. Role of the pre-neck appendage protein (Dpo7) from phage vB_SepiS-phiIPLA7 as an anti-biofilm agent in Staphylococcal species. Front. Microbiol. 2015, 6, 1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, I.W.; Hughes, K.A.; Skillman, L.C.; Tait, K. The interaction of phage and biofilms. FEMS Microbiol. Lett. 2004, 232, 1–6. [Google Scholar] [CrossRef]
- Knecht, L.E.; Veljkovic, M.; Fieseler, L. Diversity and Function of Phage Encoded Depolymerases. Front. Microbiol. 2020, 10, 2949. [Google Scholar] [CrossRef]
- Sousa, M.C.; Trame, C.B.; Tsuruta, H.; Wilbanks, S.M.; Reddy, V.S.; McKay, D.B. Crystal and Solution Structures of an HslUV Protease–Chaperone Complex. Cell 2000, 103, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Rohrwild, M.; Coux, O.; Huang, H.C.; Moerschell, R.P.; Yoo, S.J.; Seol, J.H.; Chung, C.H.; Goldberg, A.L. HslV-HslU: A novel ATP-dependent protease complex in Escherichia coli related to the eukaryotic proteasome. Proc. Natl. Acad. Sci. USA 1996, 93, 5808–5813. [Google Scholar] [CrossRef] [Green Version]
- Taylor, D.E. Bacterial tellurite resistance. Trends Microbiol. 1999, 7, 111–115. [Google Scholar] [CrossRef]
- Hendrix, R.W. Bacteriophages: Evolution of the Majority. Theor. Popul. Biol. 2002, 61, 471–480. [Google Scholar] [CrossRef]
- Haggård-Ljungquist, E.; Halling, C.; Calendar, R. DNA sequences of the tail fiber genes of bacteriophage P2: Evidence for horizontal transfer of tail fiber genes among unrelated bacteriophages. J. Bacteriol. 1992, 174, 1462–1477. [Google Scholar] [CrossRef] [Green Version]
- Rokyta, D.R.; Burch, C.L.; Caudle, S.B.; Wichman, H.A. Horizontal Gene Transfer and the Evolution of Microvirid Coliphage Genomes. J. Bacteriol. 2006, 188, 1134–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, A.; Witney, A.A.; Lindsay, J. Staphylococcus aureus Temperate Bacteriophage: Carriage and Horizontal Gene Transfer is Lineage Associated. Front. Cell. Infect. Microbiol. 2012, 2, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakasis, A.; Panitsa, G. Bacteriophage therapy as an alternative treatment for human infections. A comprehensive review. Int. J. Antimicrob. Agents 2019, 53, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G. Bacteriophage Therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Sulakvelidze, A.; Kutter, E. Bacteriophage Therapy in Humans. In Bacteriophages; CRC Press: Washington, DC, USA, 2004; pp. 381–426. [Google Scholar]
- Eaton, M.D.; Bayne-Jones, S. Bacteriophage therapy: Review of the principles and results of the use of bacteriophage in the treatment of infections. JAMA 1934, 103, 1769–1776. [Google Scholar] [CrossRef]
- Berghman, L.R.; Abi-Ghanem, D.; Waghela, S.D.; Ricke, S.C. Antibodies: an alternative for antibiotics? Poult. Sci. 2005, 84, 660–666. [Google Scholar] [CrossRef]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: an alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Kim, S.G.; Giri, S.S.; Yun, S.; Kim, H.J.; Kim, S.W.; Kang, J.W.; Han, S.J.; Kwon, J.; Oh, O.T.; Jun, J.W.; et al. Synergistic Phage-surfactant Combination Clears IgE-promoted Staphylococcus aureus Aggregation in Vitro and Enhances the Effect in Vivo. J. Antimicrob. Agents 2020, 105997. [Google Scholar] [CrossRef]
- Singla, S.; Harjai, K.; Katare, O.P.; Chhibber, S. Encapsulation of Bacteriophage in Liposome Accentuates Its Entry in to Macrophage and Shields It from Neutralizing Antibodies. PLoS ONE 2016, 11, e0153777. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Sampled Species | pSal-SNUABM-04 Susceptibility | |
|---|---|---|---|
| Common Name | Nomenclature | ||
| Sal-SNUABM-svn1 | Savannah monitor | Varanus exanthematicus | +++ |
| Sal-SNUABM-svn2 | Savannah monitor | Varanus exanthematicus | - |
| Sal-SNUABM-svn3 | Savannah monitor | Varanus exanthematicus | + |
| Sal-SNUABM-svn4 | Savannah monitor | Varanus exanthematicus | + |
| Sal-SNUABM-lp1 | Leopard gecko | Eublepharis macularius | +++ |
| Sal-SNUABM-bts1 | Common blue tongue skink | Tiliqua scincoides | - |
| Sal-SNUABM-mk1 | Milk snake | Lampropeltis triangulum | +++ |
| Sal-SNUABM-bks1 | Black king snake | Lampropeltis getula nigrita | - |
| Sal-SNUABM-gg1 | Tokay gecko | gecko gekko | - |
| Sal-SNUABM-bd1 | Bearded dragon | Pogona vitticeps | +++ |
| Sal-SNUABM-bd2 | Bearded dragon | Pogona vitticeps | + |
| Sal-SNUABM-bd3 | Bearded dragon | Pogona vitticeps | ++ |
| Sal-SNUABM-bs1 | Black rat snake | Pantherophis obsoletus | + |
| Sal-SNUABM-bp1 | Ball python | Python regius | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, J.; Kim, S.G.; Kim, H.J.; Giri, S.S.; Kim, S.W.; Lee, S.B.; Park, S.C. Isolation and Characterization of Salmonella Jumbo-Phage pSal-SNUABM-04. Viruses 2021, 13, 27. https://doi.org/10.3390/v13010027
Kwon J, Kim SG, Kim HJ, Giri SS, Kim SW, Lee SB, Park SC. Isolation and Characterization of Salmonella Jumbo-Phage pSal-SNUABM-04. Viruses. 2021; 13(1):27. https://doi.org/10.3390/v13010027
Chicago/Turabian StyleKwon, Jun, Sang Guen Kim, Hyoun Joong Kim, Sib Sankar Giri, Sang Wha Kim, Sung Bin Lee, and Se Chang Park. 2021. "Isolation and Characterization of Salmonella Jumbo-Phage pSal-SNUABM-04" Viruses 13, no. 1: 27. https://doi.org/10.3390/v13010027
APA StyleKwon, J., Kim, S. G., Kim, H. J., Giri, S. S., Kim, S. W., Lee, S. B., & Park, S. C. (2021). Isolation and Characterization of Salmonella Jumbo-Phage pSal-SNUABM-04. Viruses, 13(1), 27. https://doi.org/10.3390/v13010027