
Evaluation of the Effect of Inactivated Transmissible Gastroenteritis Virus Vaccine with Nano Silicon on the Phenotype and Function of Porcine Dendritic Cells

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fluorescent Labeling of MSNs
2.2. Preparation of Inactivated TGEV Vaccine with MSNs
2.3. Animal Experiments
2.4. Peripheral Blood Mononuclear Cells Isolation
2.5. Monocyte Derived DC Culture
2.6. MSN Toxicity Measure on DCs
2.7. Study of DC Uptake of MSNs
2.8. Intracellular Localization of MSNs
2.9. Identification of DC Phenotype
2.10. Quantifications of TLRs, Cytokines, and Cytokine Receptor Expressions in DCs
2.11. Statistical Analysis
3. Results
3.1. Characterization of the Inactivated TGEV Vaccine with MSNs
3.2. Toxicity of MSNs on im-DCs
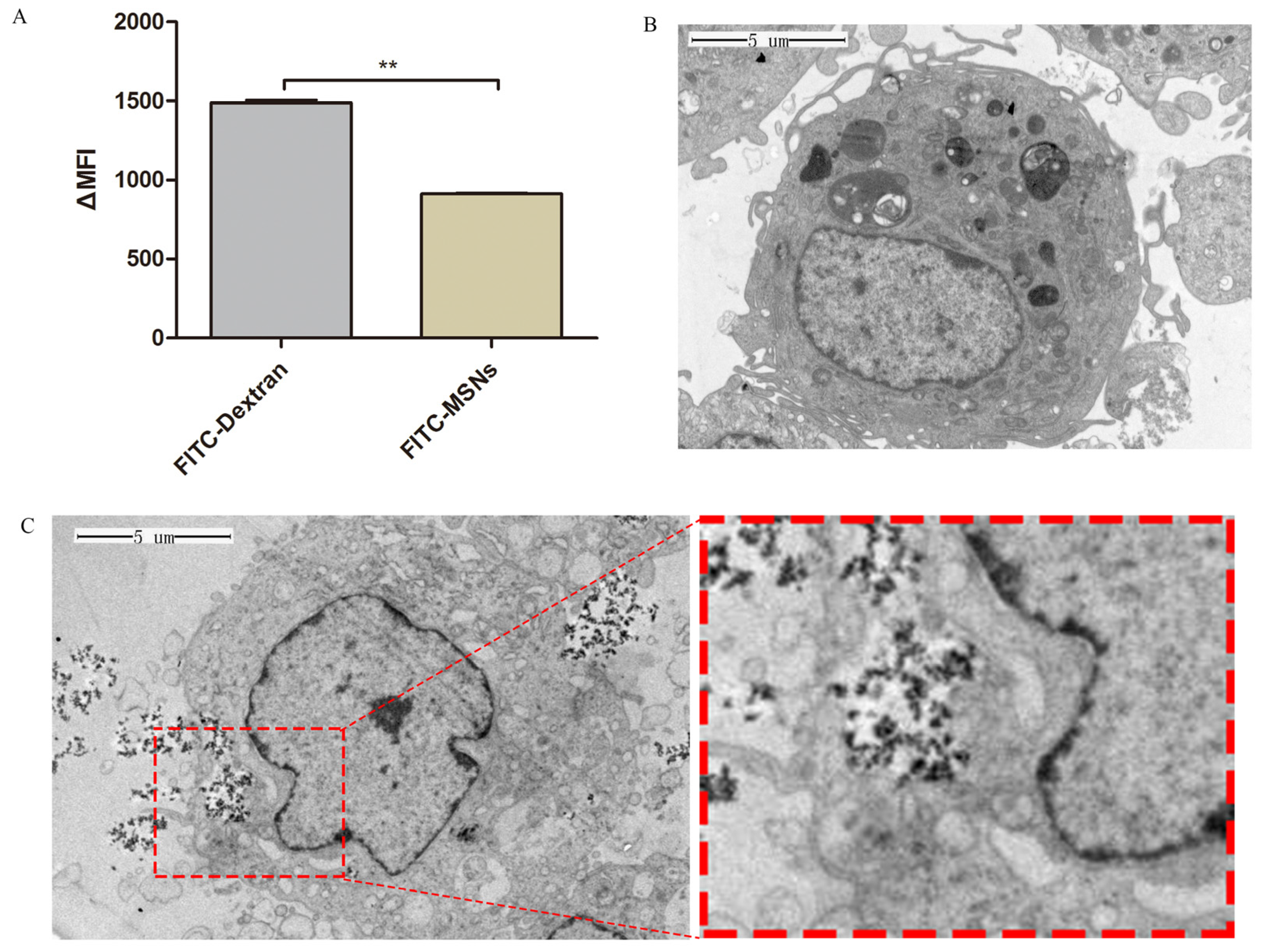
3.3. Uptake and Intracellular Localization of MSNs in im-DCs
3.4. Inactivated TGEV Vaccine with MSNs Promoting DC Maturation
3.5. mRNA Expression of TLRs
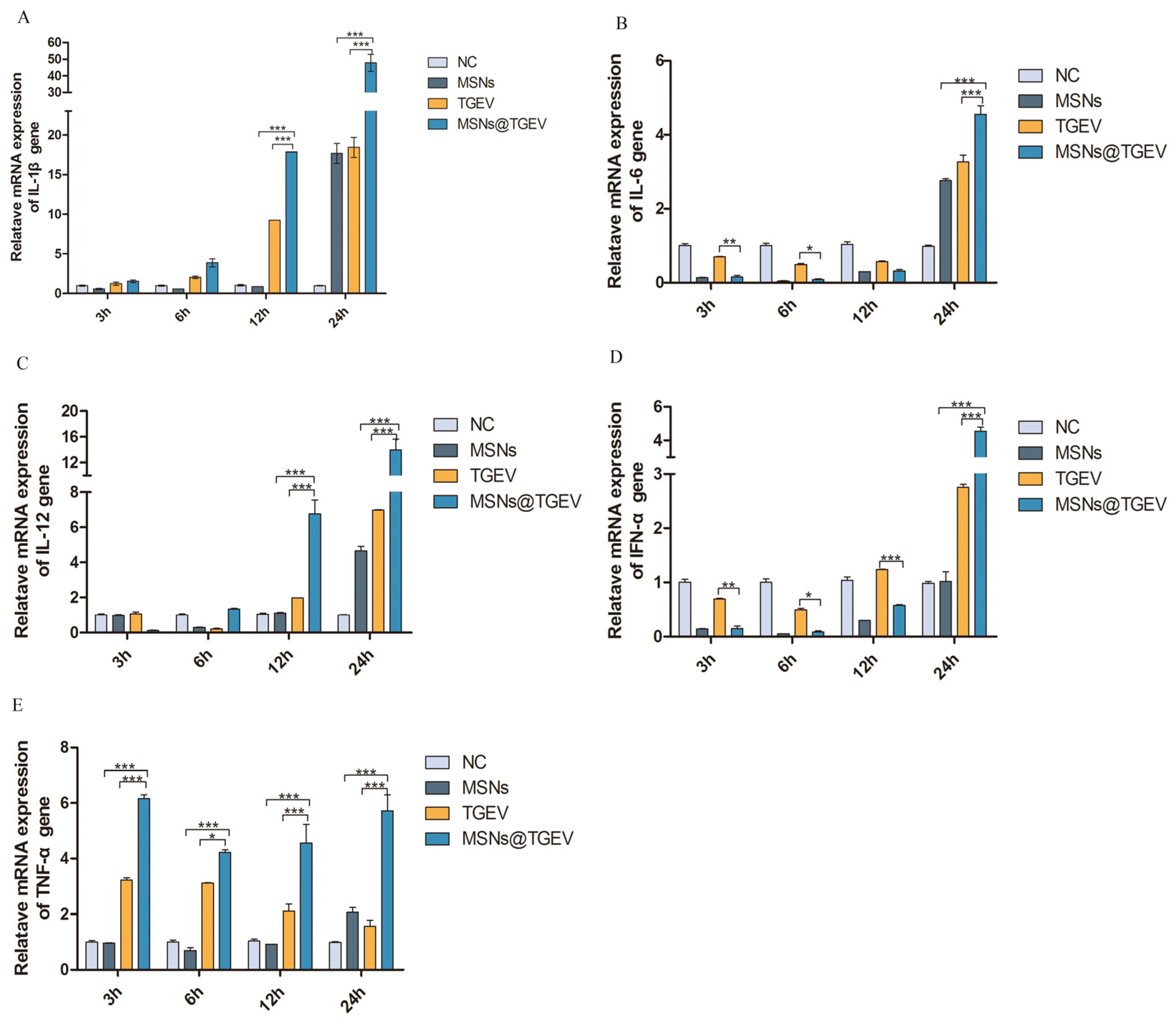
3.6. mRNA Expression of Cytokines and Cytokine Receptor
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Zhu, Y.; Zhu, X.; Chen, J.; Shi, H.; Shi, D.; Dong, H.; Feng, L. ORF3a deletion in field strains of porcine-transmissible gastroenteritis virus in China: A hint of association with porcine respiratory coronavirus. Transbound. Emerg. Dis. 2017, 64, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Yang, Y.H.; Wang, J.L.; Jing, Y.C.; Yang, Q. Impact of TGEV infection on the pig small intestine. Virol. J. 2018, 15, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, G.C.; Paton, D.J.; Wibberley, G.; Ibata, G. Transmissible gastroenteritis and porcine epidemic diarrhoea in Britain. Vet. Rec. 1999, 144, 616–618. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.L.; Fan, B.C.; Chang, X.J.; Zhou, J.Z.; Zhao, Y.X.; Shi, D.Y.; Yu, Z.Y.; He, K.W. Characterization and evaluation of the pathogenicity of a natural recombinant transmissible gastroenteritis virus in China. Virology 2020, 545, 24–32. [Google Scholar] [CrossRef]
- Valko, A.; Ba’lint, A.; Bozsa, A.; Csa’gola, A. Prevalence of antibodies against transmissible gastroenteritis virus (TGEV) in Hungary. Vet. Anim. Sci. 2019, 7, 100042. [Google Scholar] [CrossRef]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef]
- Mody, K.T.; Popat, A.; Mahony, D.; Cavallaro, A.S.; Yu, C.; Mitter, N. Mesoporous silica nanoparticles as antigen carriers and adjuvants for vaccine delivery. Nanoscale 2013, 5, 5167–5179. [Google Scholar] [CrossRef]
- Neun, B.W.; Dobrovolskaia, M.A. Analysis of nanoparticle adjuvant properties in vivo. Methods. Mol. Biol. 2018, 1682, 189–195. [Google Scholar]
- Jin, X.H.; Zheng, L.L.; Song, M.R.; Xu, W.S.; Kou, Y.N.; Zhou, Y.; Zhang, L.W.; Zhu, Y.N.; Wan, B.; Wei, Z.Y.; et al. A nano silicon adjuvant enhances inactivated transmissible gastroenteritis vaccine through activeation the Toll-like receptors and promotes humoral and cellular immune responses. Nanomedicine 2018, 14, 1201–1212. [Google Scholar] [CrossRef]
- Hubbell, J.A.; Thomas, S.N.; Swartz, M.A. Materials engineering for immunomodulation. Nature 2009, 462, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Gayo, M.E.; Yu, X.G. Role of Dendritic Cells in Natural Immune Control of HIV-1 Infection. Front. Immunol. 2019, 10, 1306. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Sathe, P.; Helft, J.; Miller, J.; Mortha, A. The dendritic cell lineage: Ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef] [Green Version]
- Hochweller, K.; Wabnitz, G.H.; Samstag, Y.; Suffner, J.; Hämmerling, G.J.; Mitchison, N.G. Dendritic cells control T cell tonic signaling required for responsiveness to foreign antigen. Proc. Natl. Acad. Sci. USA 2010, 107, 5931–5936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.J.; Ru, J.X.; Sun, S.Q.; Teng, Z.D.; Dong, H.; Song, P.; Yang, Y.; Guo, H.C. Uniform dendrimer-like mesoporous silica nanoparticles as a nano-adjuvant for foot-and-mouth disease virus-like particle vaccine. J. Mater. Chem. B. 2019, 7, 3446–3454. [Google Scholar] [CrossRef]
- Zhao, F.J.; Liu, L.T.; Xu, M.L.; Shu, X.L.; Zheng, L.L.; Wei, Z.Y. Assessments of different inactivating reagents in formulating transmissible gastroenteritis virus vaccine. Virol. J. 2020, 17, 163. [Google Scholar] [CrossRef]
- Lin, C.M.; Jeng, C.R.; Chang, H.W.; Guo, I.C.; Huang, Y.L.; Tsai, Y.C.; Chia, M.Y.; Pang, V.F. Characterization of porcine circovirus type 2 (PCV2) infection in swine lymphocytes using mitogen-stimulated peripheral blood lymphocytes from healthy PCV2-carrier pigs. Vet. Immunol. Immunopathol. 2008, 124, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.C.; Jeng, C.R.; Hsiao, S.H.; Chang, H.W.; Liu, J.J.; Chang, C.C.; Lin, C.M.; Chia, M.Y.; Pang, V.F. Porcine circovirus type 2 (PCV2) induces cell proliferation, fusion, and chemokine expression in swine monocytic cells in vitro. Vet. Res. 2010, 41, 60. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Messerle, M.; Sapinoro, R.; Santos, K.; Hocknell, P.K.; Jin, X.; Dewhurst, S. Murine cytomegalovirus abortively infects human dendritic cells, leading to expression and presentation of virally vectored genes. J. Virol. 2003, 77, 7182–7192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Zhao, S.; Qin, T.; Yin, Y.; Yang, Q. Effects of porcine epidemic diarrhea virus on porcine monocyte-derived dendritic cells and intestinal dendritic cells. Vet. Microbiol. 2015, 179, 131–141. [Google Scholar] [CrossRef]
- Ferreira, R.S., Jr.; Anderlini, R.P.; Pimenta, D.C.; Oliveira orsi, R.D.; Barraviera, B.; Sant’anna, O.A. New nanostructured silica adjuvant (SBA-15) employed to produce antivenom in young sheep using Crotalus durissus terrificus and Apis mellifera venoms detoxified by cobalt-60. J. Toxicol. Environ. Health A 2010, 73, 926–933. [Google Scholar] [CrossRef]
- Möller, K.; Bein, T. Degradable Drug Carriers: Vanishing Mesoporous Silica Nanoparticles. Chem. Mater. 2019, 31, 4364–4378. [Google Scholar] [CrossRef]
- Vallet-Regi, M.; Colilla, M.; Izquierdo-Barba, I.; Manzano, M. Mesoporous Silica Nanoparticles for Drug Delivery: Current Insights. Molecules 2018, 23, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meka, A.K.; Abbaraju, P.L.; Song, H.; Xu, C.; Zhang, J.; Zhang, H.; Yu, M.; Yu, C. A Vesicle Supra-Assembly Approach to Synthesize Amine-Functionalized Hollow Dendritic Mesoporous Silica Nanospheres for Protein Delivery. Small 2016, 12, 5169–5177. [Google Scholar] [CrossRef] [PubMed]
- Croissant, J.G.; Fatieiev, Y.; Khashab, N.M. Degradability and Clearance of Silicon, Organosilica, Silsesquioxane, Silica Mixed Oxide, and Mesoporous Silica Nanoparticles. Adv. Mater. 2017, 29, 1604634. [Google Scholar] [CrossRef]
- Bai, M.; Dong, H.; Su, X.; Jin, Y.; Sun, S.; Zhang, Y.; Yang, Y.; Guo, H. Hollow mesoporous silica nanoparticles as delivery vehicle of foot-and-mouth disease virus-like particles induce persistent immune responses in guinea pigs. J. Med. Virol. 2019, 91, 941–948. [Google Scholar] [CrossRef] [Green Version]
- Reffitt, D.M.; Jugdaohsingh, R.; Thompson, R.P.; Powell, J.J. Silicic acid: Its gastrointestinal uptake and urinary excretion in man and effects on aluminium excretion. J. Inorg. Biochem. 1999, 76, 141–147. [Google Scholar] [CrossRef]
- Mohammadpour, R.; Cheney, D.L.; Grunberger, J.W.; Yazdimamaghani, M.; Jedrzkiewicz, J.; Isaacson, K.J.; Dobrovolskaia, M.A.; Ghandehari, H. One-year chronic toxicity evaluation of single dose intravenously administered silica nanoparticles in mice and their Ex vivo human hemocompatibility. J. Control Release 2020, 324, 471–481. [Google Scholar] [CrossRef]
- Langel, S.N.; Paim, F.C.; Lager, K.M.; Vlasova, A.N.; Saif, L.J. Lactogenic immunity and vaccines for porcine epidemic diarrhea virus (PEDV): Historical and current concepts. Virus. Res. 2016, 226, 93–107. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.C.; Everett, H.E.; Pedrera, M.; Mokhtar, H.; Marchi, E.; Soldevila, F.; Kaveh, D.A.; Hogarth, P.J.; Johns, H.L.; Nunez-Garcia, J. CD1− and CD1+ porcine blood dendritic cells are enriched for the orthologues of the two major mammalian conventional subsets. Sci. Rep. 2017, 7, 40942. [Google Scholar] [CrossRef] [Green Version]
- Boraschi, D.; Italiani, P.; Palomba, R.; Decuzzi, P.; Duschl, A.; Fadeel, B.; Moghimi, S.M. Nanoparticles and innate immunity: New perspectives on host defence. Semin. Immunol. 2017, 34, 33–51. [Google Scholar] [CrossRef]
- Franzoni, G.; Graham, S.P.; Dei Giudici, S.; Oggiano, A. Porcine Dendritic Cells and Viruses: An Update. Viruses 2019, 11, 445. [Google Scholar] [CrossRef] [Green Version]
- Facci, M.R.; Auray, G.; Buchanan, R.; Kessel, J.; Thompson, D.R.; Mackenzie-Dyck, S.; Babiuk, L.A.; Gerdts, V. A comparison between isolated blood dendritic cells and monocyte-derived dendritic cells in pigs. Immunology 2009, 129, 396–405. [Google Scholar] [CrossRef]
- Blank, F.; Stumbles, P.A.; Seydoux, E.; Holt, P.G.; Fink, A.; Rothen-Rutishauser, B.; Strickland, D.H.; Von Garnier, C. Size-dependent Uptake of Particles by Pulmonary APC Populations and Trafficking to Regional Lymph Nodes. Am. J. Respir. Cell. Mol. Biol. 2013, 49, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Seydoux, E.; Rothen-Ruthishauser, B.; Nita, I.; Gazdhar, A.; Petri-Fink, A.; Blank, F.; Von Garnier, C.; Balog, S.; Stumbles, P. Size-dependent accumulation of particles in lysosomes modulates dendritic cell function through impaired antigen degradation. Int. J. Nanomed. 2014, 9, 3885–3902. [Google Scholar] [CrossRef] [Green Version]
- Santos, T.D.; Varela, J.; Lynch, I.; Salvati, A.; Dawson, K.A. Quantitative assessment of the comparative nanoparticle-uptake efficiency of a range of cell lines. Small 2011, 7, 3341–3349. [Google Scholar] [CrossRef] [Green Version]
- Pathak, S.K.; Sköld, A.E.; Mohanram, V.; Persson, C.; Johansson, U.; Spetz, A.L. Activated apoptotic cells induce dendritic cell maturation via engagement of Toll-like receptor 4 (TLR4), dendritic cell-specific intercellular adhesion molecule 3 (ICAM-3)-grabbing nonintegrin (DC-SIGN), and β2 integrins. J. Biol. Chem. 2012, 287, 13731–13742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auray, G.; Keller, I.; Python, S.; Gerber, M.; Bruggmann, R.; Ruggli, N.A. Summerfield, Characterization and transcriptomic analysis of porcine blood conventional and plasmacytoid dendritic cells reveals striking species-specific differences. J. Immunol. 2016, 197, 4791–4806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, M.; Bigley, V. Human dendritic cell subsets: An update. Immunology 2018, 154, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Vreman, S.; Auray, G.; Savelkoul, H.F.J.; Rebel, A.; Summerfield, A.; Stockhofe-Zurwieden, N. Neonatal porcine blood derived dendritic cell subsets show activation after TLR2 or TLR9 stimulation. Dev. Comp. Immunol. 2018, 84, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Summerfield, A.; McCullough, K.C. The porcine dendritic cell family. Dev. Comp. Immunol. 2009, 33, 299–309. [Google Scholar] [CrossRef]
- Warner, N.; Nunez, G. MyD88: A critical adaptor protein in innate immunity signal transduction. J. Immunol. 2013, 190, 3–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summerfield, A.; Guzylack-Piriou, L.; Schaub, A.; Carrasco, C.P.; Tche, V.; Charley, B.; Mccullough, K.C. Porcine peripheral blood dendritic cells and natural interferon-producing cells. Immunology 2003, 110, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 2003, 3, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, S.; Renukaradhya, G.J. Nanoparticle-based vaccine development and evaluation against viral infections in pigs. Vet. Res. 2019, 50, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession Numbers in Genbank | Primers (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|
| TLR-1 | NM001031775.1 | F: CCAGAGCTGCCAGAAGATTAG R:TCTACCACGTCACTCGATACT | 114 |
| TLR-2 | NM213761.1 | F: GGAGCCTTAGAAGTAGAGTTTGA R: AAGGGAACAGGGAACCAG | 234 |
| TLR-3 | NM001097444.1 | F: ATGCTCCGAAGGGTGG R: GGGTTTGCGTGTTTCC | 111 |
| TLR-4 | NM001293316.1 | F: CCACCTGTCAGATAAGCG R: CCTCACCCAGTCTTCGTC | 102 |
| TLR-5 | NM001348771.1 | F: CCACCAGGAGTCTTTCGC R: CGGCACTTAGTGAGGTGAAT | 111 |
| TLR-6 | NM213760.2 | F: ATCACCAGCCTCAAGCATTT R: TAGCCAGTTGTAAACACCCT | 114 |
| TLR-7 | NM001097434.1 | F: CCTTTCTGTCTCTCTGTGTCTTC R: CACCCTTCTCCCAACAGTATTT | 81 |
| TLR-8 | NM214187.1 | F: CTTTGATGATGACGCTGCTTTC R: GGTGTGTCACTCCTGCTATTC | 99 |
| TLR-9 | NM213958.1 | F: TTACTAGGGAGGTGGATGGTAG R: CCTTGCAGTTTGGCATGAAG | 108 |
| TLR-10 | NM001030534.1 | F: CACCACCACCTCTTCCATAAA R: AGAGCTTTCAGTGCAGGATAC | 104 |
| IL-1β | NM214055.1 | F: ACCTGGACCTTGGTTCTC R: GGATTCTTCATCGGCTTC | 124 |
| IL-6 | NM001252429.1 | F: TTCAGTCCAGTCGCCTTCT R: GTGGCATCACCTTTGGCATCTTCTT | 99 |
| IL-12 | NM214097.2 | F: AGTGCCCTCAGTAAGAGTAAGA R: CTTGTGTTCCCACCCATCAA | 86 |
| TNF-α | NM214022.1 | F: CCCTTGAGCATCAACCCT R: GCATTGGCATACCCACTCT | 131 |
| IFN-α | JQ839262.1 | F: CATCCTGGCTGTGAGGAAATAC R: CAGGTTTGTGGAGGAAGAGAAG | 118 |
| CCR-7 | NM_001001532.3 | F: CGGTCATGTACTCCATCATCTG R: ATGGTCTTGAGCCTCTTGAAATA | 91 |
| β-actin | DQ452569.1 | F: TTCCAGCCCTCCTTCCTG R: AGGTCCTTGCGGATGTCG | 94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, L.; Zhao, F.; Ru, J.; Liu, L.; Wang, Z.; Wang, N.; Shu, X.; Wei, Z.; Guo, H. Evaluation of the Effect of Inactivated Transmissible Gastroenteritis Virus Vaccine with Nano Silicon on the Phenotype and Function of Porcine Dendritic Cells. Viruses 2021, 13, 2158. https://doi.org/10.3390/v13112158
Zheng L, Zhao F, Ru J, Liu L, Wang Z, Wang N, Shu X, Wei Z, Guo H. Evaluation of the Effect of Inactivated Transmissible Gastroenteritis Virus Vaccine with Nano Silicon on the Phenotype and Function of Porcine Dendritic Cells. Viruses. 2021; 13(11):2158. https://doi.org/10.3390/v13112158
Chicago/Turabian StyleZheng, Lanlan, Fujie Zhao, Jiaxi Ru, Lintao Liu, Zi Wang, Nianxiang Wang, Xiangli Shu, Zhanyong Wei, and Huichen Guo. 2021. "Evaluation of the Effect of Inactivated Transmissible Gastroenteritis Virus Vaccine with Nano Silicon on the Phenotype and Function of Porcine Dendritic Cells" Viruses 13, no. 11: 2158. https://doi.org/10.3390/v13112158
APA StyleZheng, L., Zhao, F., Ru, J., Liu, L., Wang, Z., Wang, N., Shu, X., Wei, Z., & Guo, H. (2021). Evaluation of the Effect of Inactivated Transmissible Gastroenteritis Virus Vaccine with Nano Silicon on the Phenotype and Function of Porcine Dendritic Cells. Viruses, 13(11), 2158. https://doi.org/10.3390/v13112158