
Genome Organizations and Functional Analyses of a Novel Gammapartitivirus from Rhizoctonia solani AG-1 IA Strain D122


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Cultural Conditions
2.2. Extraction and Detection of dsRNA
2.3. cDNA Cloning, Sequencing and Sequence Analysis
2.4. Northern Hybridization
2.5. Purification of Viral Particles and Electron Microscopy
2.6. Virus-Elimination
2.7. RNA-Seq and Data Analysis
3. Results
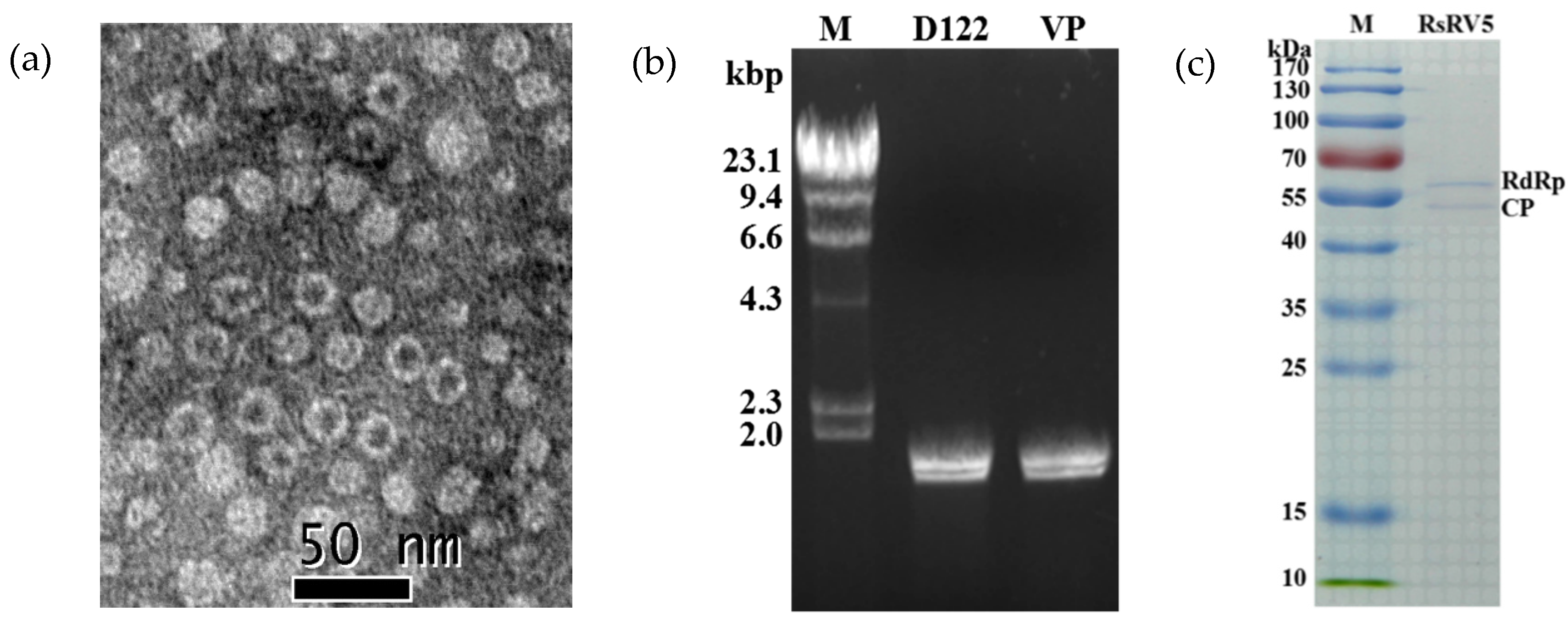
3.1. Detection of dsRNAs in R. solani Strains
3.2. Nucleotide Sequence and Amino Acid Sequence of RsRV5
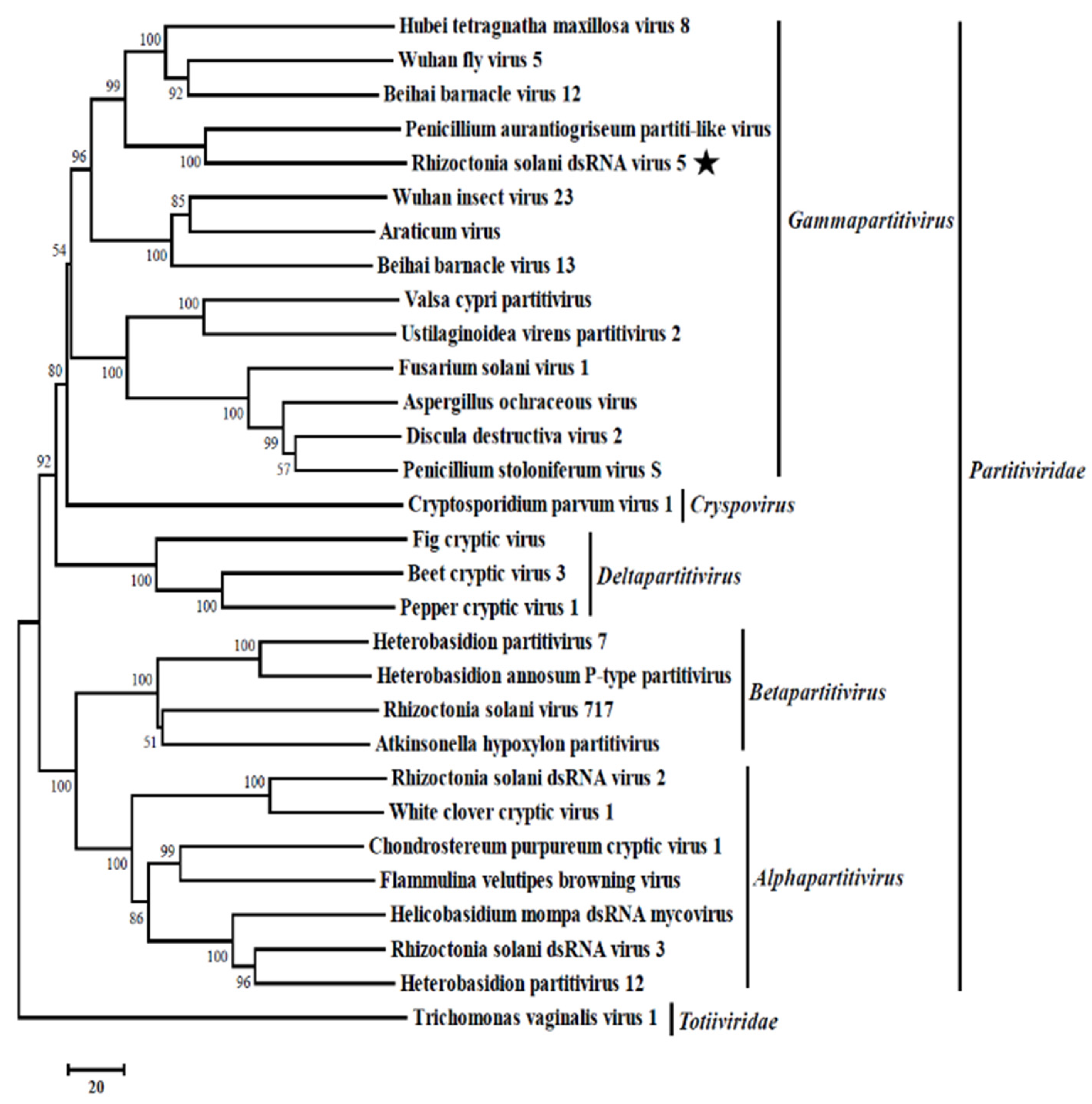
3.3. Phylogenetic Analysis of the Partitivirus RsRV5
3.4. Viral Particles
3.5. The Mycovirus Affects the Fungal Host Phenotypes in Strain D122
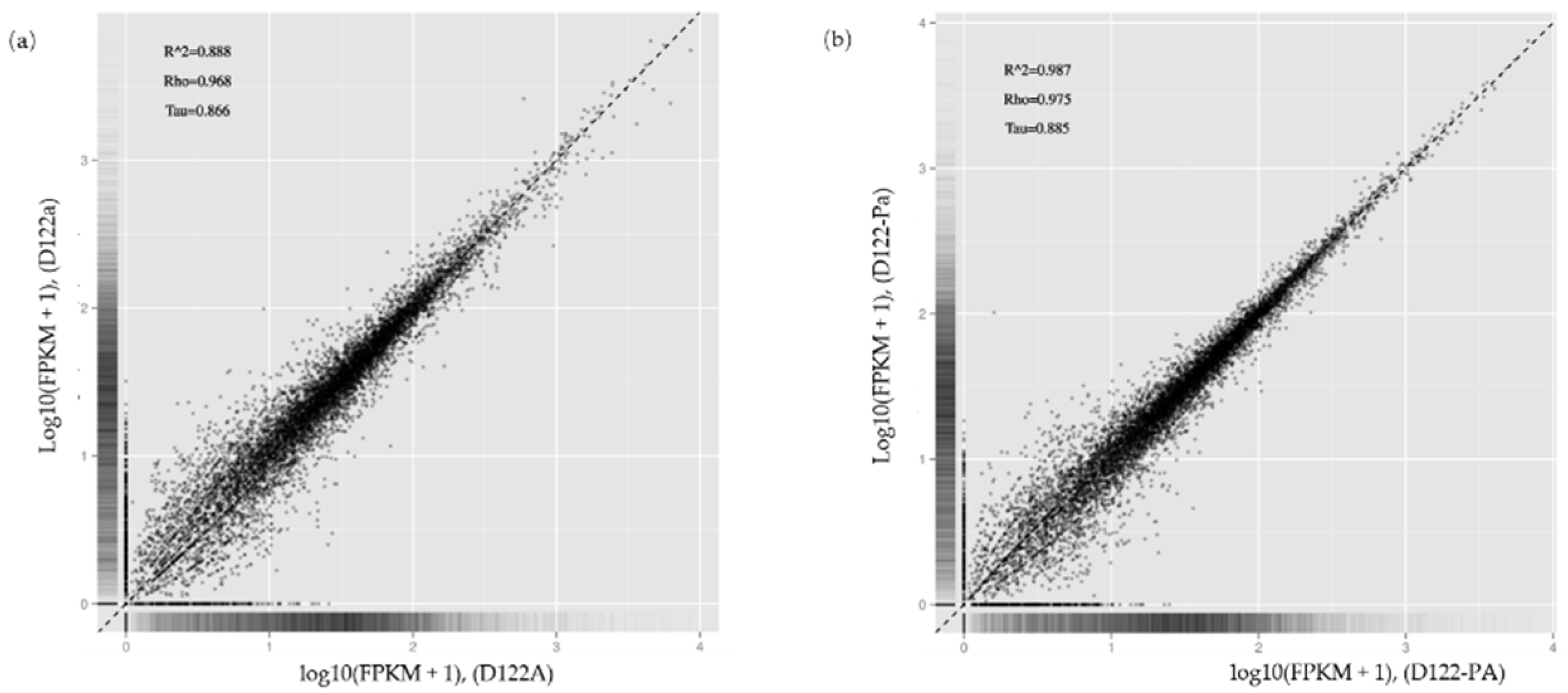
3.6. RNA-seq Analysis of Rhizoctonia solani AG-1 IA Response to RsRV5 Infection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant. Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Pedrajas, M.D.; Canizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G. Mycoviruses in biological control: From basic research to field implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Coutts, R.H.A. Mycoviruses in Aspergilli: A comprehensive review. Front. Microbiol. 2017, 8, 1699. [Google Scholar] [CrossRef] [Green Version]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef] [Green Version]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.H. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. ICTV virus taxonomy profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in plain sight: New virus genomes discovered via a systematic analysis of fungal public transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef] [Green Version]
- Milgroom, M.G.; Cortesi, P. Biological control of chestnut blight with hypovirulence: A critical analysis. Annu. Rev. Phytopathol. 2004, 42, 311–338. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.X.; Nuss, D.L. Engineering super mycovirus donor strains of chestnut blight fungus by systematic disruption of multilocus vic genes. Proc. Natl. Acad. Sci. USA 2016, 113, 2062–2067. [Google Scholar] [CrossRef] [Green Version]
- Ihrmark, K.; Zheng, J.; Stenström, E.; Stenlid, J. Transmission of double-stranded RNA in Heterobasidion annosum. Fungal Genet. Biol. 2002, 36, 147–154. [Google Scholar] [CrossRef]
- Valverde, R.A.; Gutierrez, D.L. Molecular and biological properties of a putative partitivirus from jalapeño pepper (Capsicum annuum L.). Rev. Mex. Fitopatol. 2008, 26, 1–6. [Google Scholar]
- Liu, C.; Zeng, M.; Zhang, M.; Shu, C.; Zhou, E. Complete nucleotide sequence of a partitivirus from Rhizoctonia solani AG-1 IA strain C24. Viruses 2018, 10, 703. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Shu, C.; Zhang, M.; Yang, M.; Zhou, E. Molecular characterization of a novel endornavirus conferring hypovirulence in rice sheath blight fungus Rhizoctonia solani AG-1 IA strain GD-2. Viruses 2019, 11, 178. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Zhang, M.; Chen, Q.; Zhu, M.; Zhou, E. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani. Virology 2014, 456–457, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.L.; Zheng, L.; Liu, C.; Shu, C.W.; Zhou, E.X. Characterization of a novel dsRNA mycovirus isolated from strain A105 of Rhizoctonia solani AG-1 IA. Arch. Virol. 2018, 163, 427–430. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, H.; Zhang, M.; Cao, X.; Zhou, E. The complete genomic sequence of a novel mycovirus from Rhizoctonia solani AG-1 IA strain B275. Arch. Virol. 2013, 158, 1609–1612. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Yang, M.; Li, M.H.; Zhou, E.X. Cloning and functional analysis of an endo-PG-encoding gene Rrspg1 of Rhizoctonia solani, the causal agent of rice sheath blight. Can. J. Plant Pathol. 2012, 34, 1–12. [Google Scholar] [CrossRef]
- Morris, T.J.; Dodds, J.A. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854–858. [Google Scholar] [CrossRef] [Green Version]
- Darissa, O.; Willingmann, P.; Adam, G. Optimized approaches for the sequence determination of double-stranded RNA templates. J. Virol. Methods 2010, 169, 397–403. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Sanderlin, R.S.; Ghabrial, S.A. Physiochemical properties of two distinct types of virus-like particles from Helminthosporium victoriae. Virology 1978, 87, 142–151. [Google Scholar] [CrossRef]
- Sasaki, A.; Kanematsu, S.; Onoue, M.; Oikawa, Y.; Nakamura, H.; Yoshida, K. Artificial infection of Rosellinia necatrix with purified viral particles of a member of the genus Mycoreovirus reveals its uneven distribution in single colonies. Phytopathology 2007, 97, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Hillman, B.I.; Supyani, S.; Kondo, H.; Suzuki, N. A reovirus of the fungus Cryphonectria parasitica that is infectious as particles and related to the Coltivirus genus of animal pathogens. J. Virol. 2004, 78, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Lim, W.S.; Jeong, J.H.; Jeong, R.D.; Yoo, Y.B.; Yie, S.W.; Kim, K.H. Complete nucleotide sequence and genome organization of a dsRNA partitivirus infecting Pleurotus ostreatus. Virus Res. 2005, 108, 111–119. [Google Scholar] [CrossRef]
- Osaki, H.; Nomura, K.; Iwanami, T.; Kanematsu, S.; Okabe, I.; Matsumoto, N.; Sasaki, A.; Ohtsu, Y. Detection of a double-stranded RNA virus from a strain of the violet root rot fungus Helicobasidium mompa Tanaka. Virus Genes 2002, 25, 139–145. [Google Scholar] [CrossRef]
- Strauss, E.E.; Lakshman, D.K.; Tavantzis, S.M. Molecular characterization of the genome of a partitivirus from the basidiomycete Rhizoctonia solani. J. Gen. Virol. 2000, 81, 549–555. [Google Scholar] [CrossRef]
- Nerva, L.; Ciuffo, M.; Vallino, M.; Margaria, P.; Varese, G.C.; Gnavi, G.; Turina, M. Multiple approaches for the detection and characterization of viral and plasmid symbionts from a collection of marine fungi. Virus Res. 2016, 219, 22–38. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Chen, Y.; Gai, X.T.; Chen, R.X.; Li, C.X.; Zhong, J. Characterization of three novel betapartitiviruses co-infecting the phytopathogenic fungus Rhizoctonia solani. Virus Res. 2019, 270, 197649. [Google Scholar] [CrossRef]
- Lyu, R.; Zhang, Y.; Tang, Q.; Li, Y.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Xie, J. Two alphapartitiviruses co-infecting a single isolate of the plant pathogenic fungus Rhizoctonia solani. Arch. Virol. 2017, 163, 515–520. [Google Scholar] [CrossRef]
- Ribas, J.C.; Fujimura, T. Essential RNA binding and packaging domains of the Gag-Pol fusion protein of the L-A double-stranded RNA virus of Saccharomyces cerevisiae. J. Biol. Chem. 1994, 269, 28420–28428. [Google Scholar] [CrossRef]
- MacDonald, W.L.; Fulbright, D.W. Biological control of chestnut blight: Use and limitations of transmissible hypovirulence. Plant Dis. 1991, 75, 656–661. [Google Scholar] [CrossRef]
- Zhong, J.; Chen, D.; Lei, X.H.; Zhu, H.J.; Zhu, J.Z.; Gao, B.D. Detection and characterization of a novel Gammapartitivirus in the phytopathogenic fungus Colletotrichum acutatum strain HNZJ001. Virus Res. 2014, 190, 104–109. [Google Scholar] [CrossRef]
- Qu, Z.; Fu, Y.; Lin, Y.; Zhao, Z.; Jiang, D. Transcriptional responses of Sclerotinia sclerotiorum to the infection by SsHADV-1. J. Fungi 2021, 7, 493. [Google Scholar] [CrossRef]
- Wang, B.; Guo, G.; Wang, C.; Lin, Y.; Wang, X.; Zhao, M.; Guo, Y.; He, M.; Zhang, Y.; Pan, L. Survey of the transcriptome of Aspergillus oryzae via massively parallel mRNA sequencing. Nucleic Acids Res. 2010, 38, 5075–5087. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alternative Splicing Types | Alternative Splicing Numbers | Numbers of Genes Involved |
|---|---|---|
| exon skipping | 7792 | 2662 |
| intron retention | 13,377 | 3965 |
| alternative 5′ss splice or alternative 3′ss splice | 18,396 | 5088 |
| alternative transcription start site | 13,466 | 10,687 |
| alternative transcription terminal site | 12,504 | 10,687 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; He, Z.; Huang, X.; Shu, C.; Zhou, E. Genome Organizations and Functional Analyses of a Novel Gammapartitivirus from Rhizoctonia solani AG-1 IA Strain D122. Viruses 2021, 13, 2254. https://doi.org/10.3390/v13112254
Zhang M, He Z, Huang X, Shu C, Zhou E. Genome Organizations and Functional Analyses of a Novel Gammapartitivirus from Rhizoctonia solani AG-1 IA Strain D122. Viruses. 2021; 13(11):2254. https://doi.org/10.3390/v13112254
Chicago/Turabian StyleZhang, Meiling, Zhenrui He, Xiaotong Huang, Canwei Shu, and Erxun Zhou. 2021. "Genome Organizations and Functional Analyses of a Novel Gammapartitivirus from Rhizoctonia solani AG-1 IA Strain D122" Viruses 13, no. 11: 2254. https://doi.org/10.3390/v13112254
APA StyleZhang, M., He, Z., Huang, X., Shu, C., & Zhou, E. (2021). Genome Organizations and Functional Analyses of a Novel Gammapartitivirus from Rhizoctonia solani AG-1 IA Strain D122. Viruses, 13(11), 2254. https://doi.org/10.3390/v13112254