
A Rat Model of Prenatal Zika Virus Infection and Associated Long-Term Outcomes

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Subjects
2.2. ZIKV Growth Conditions
2.3. ZIKV Infection
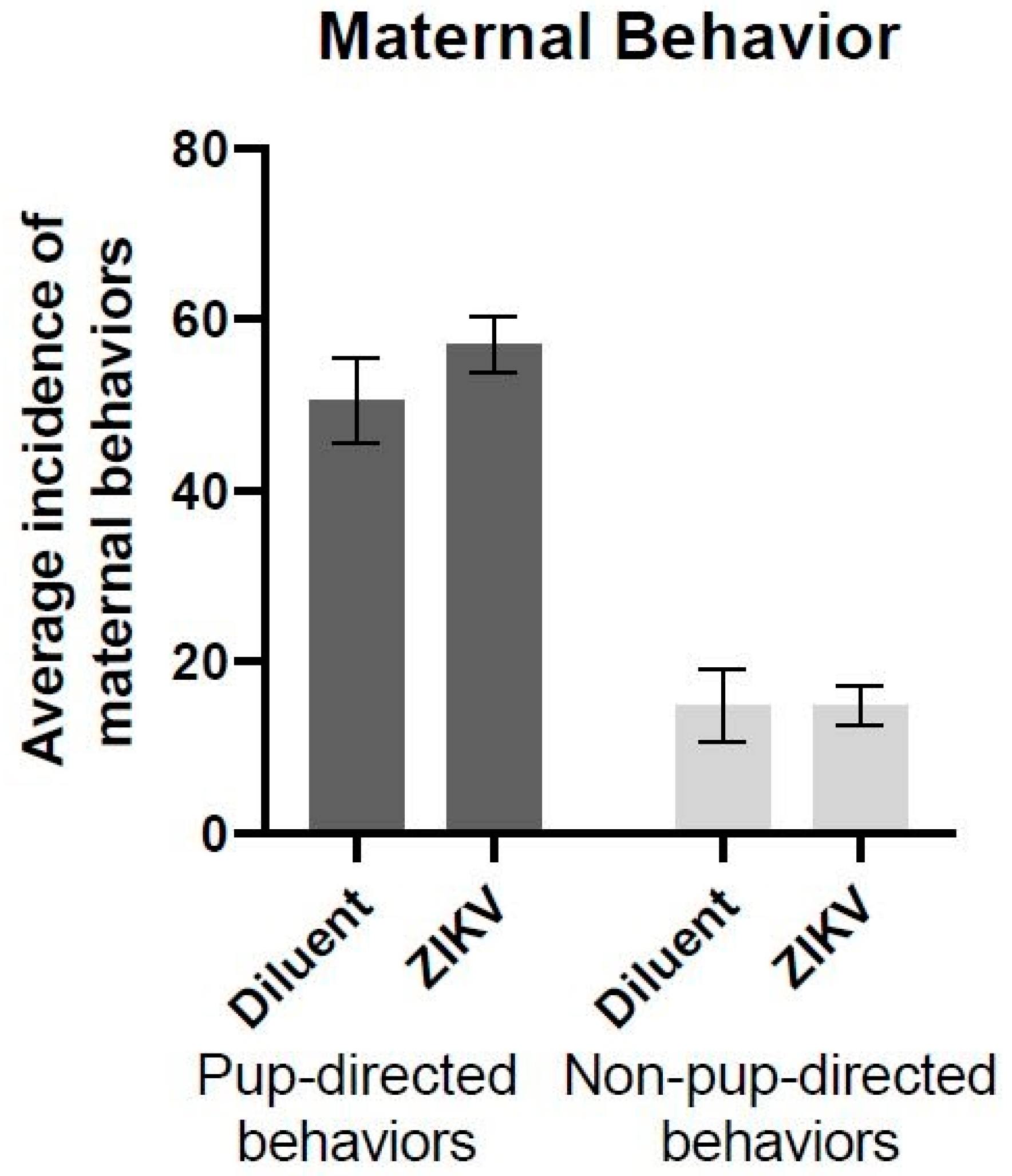
2.4. Maternal Behavior
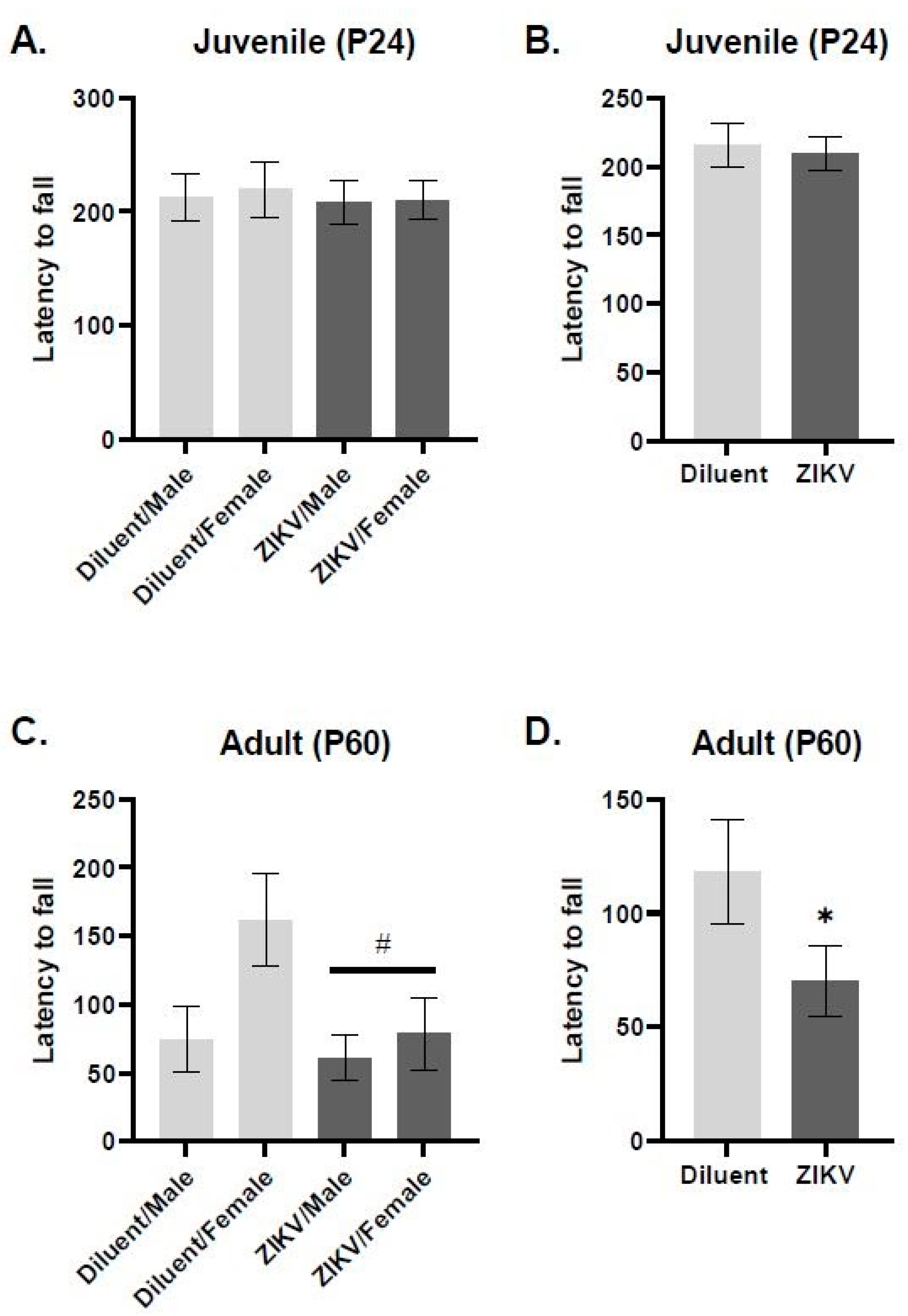
2.5. Rotarod
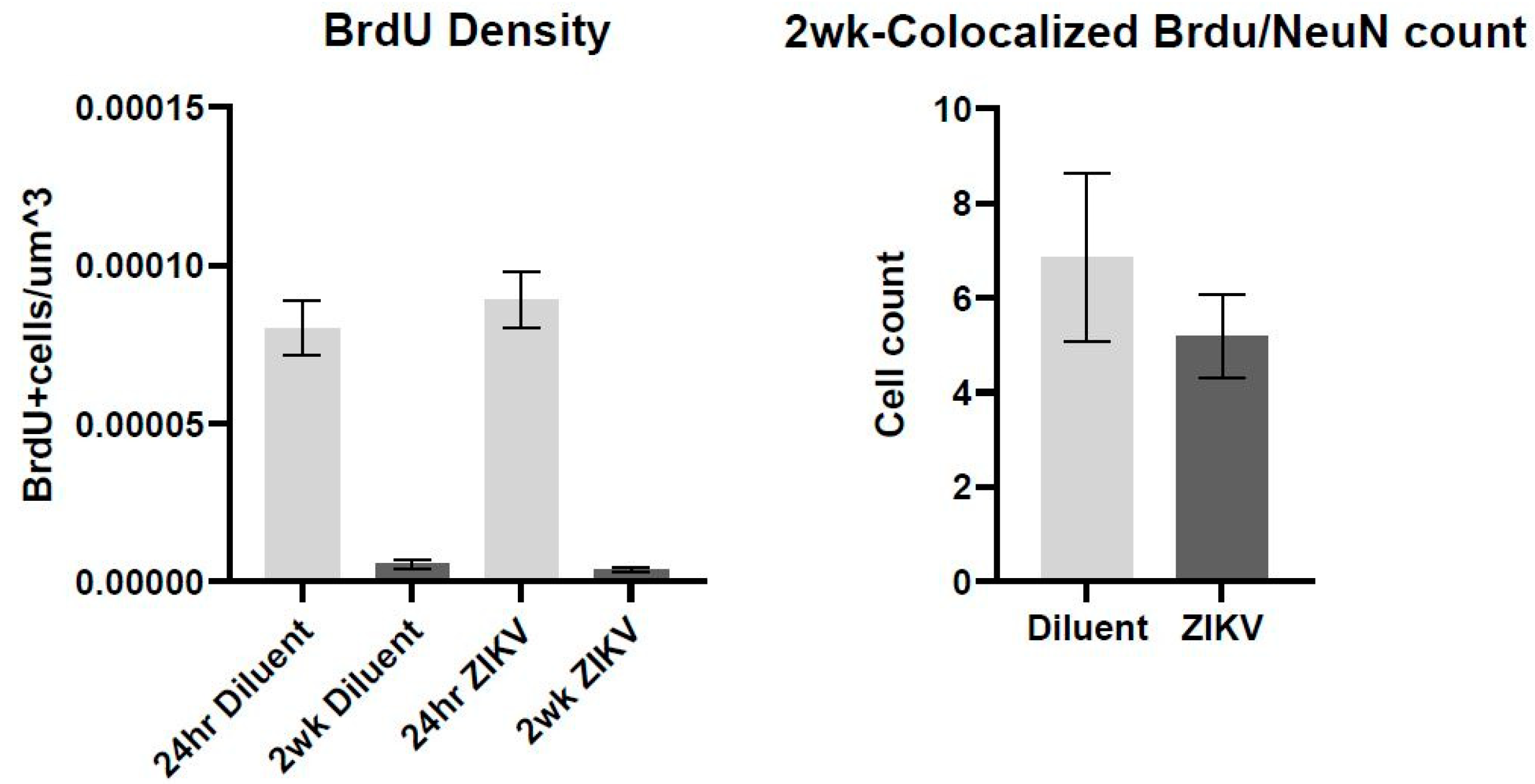
2.6. BrdU Administration
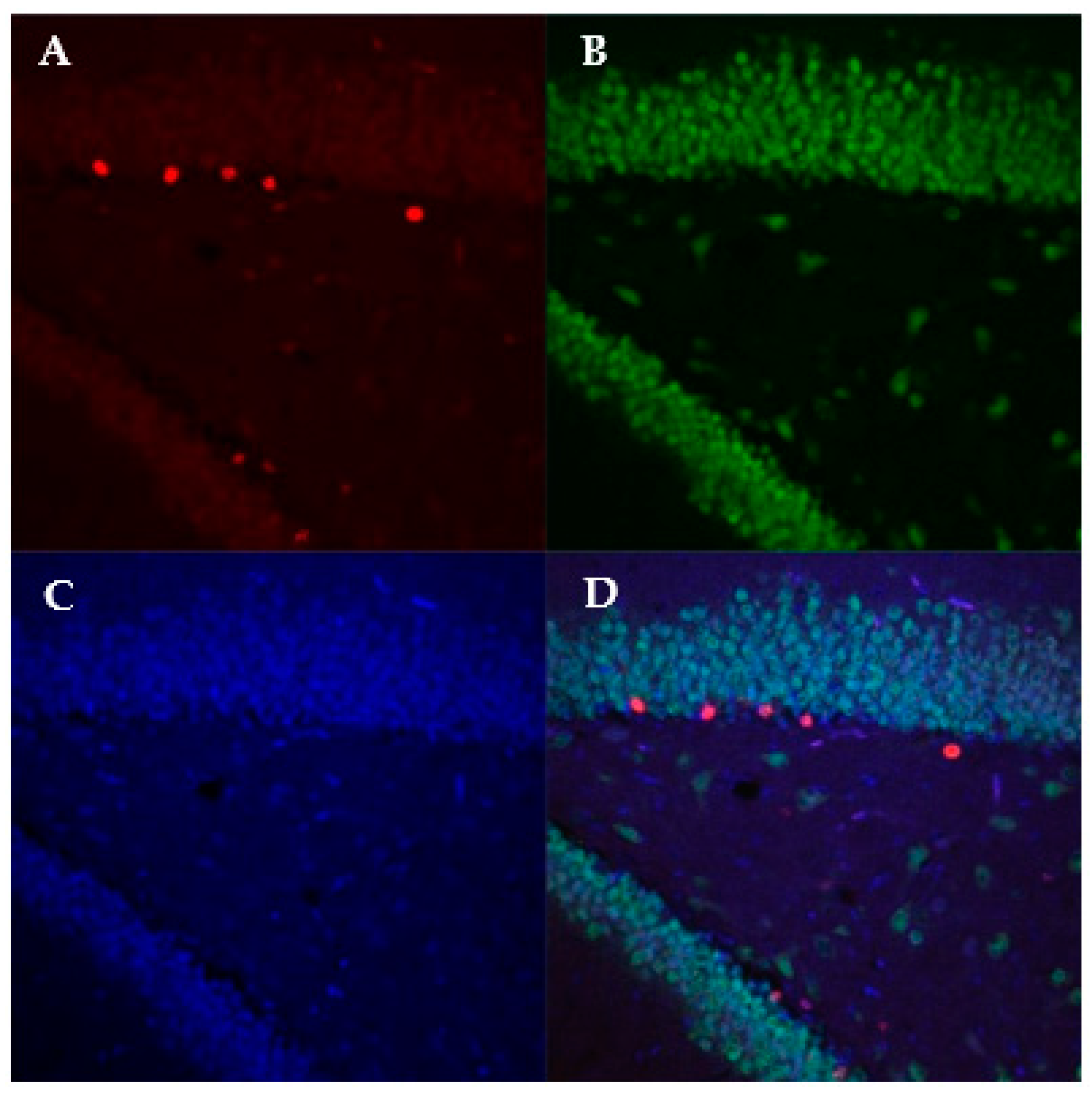
2.7. Immunohistochemistry
2.8. Confocal Imaging and Imaris Analysis
2.9. Poly(I:C) Injection
2.10. Real-Time PCR
2.11. Analysis of Gene Expression following Treatment with Poly(I:C)
3. Results
3.1. Maternal Behavior
3.2. Rotarod
3.3. Immunohistochemistry
3.4. RT-qPCR
3.4.1. ZIKV Presence and Persistence
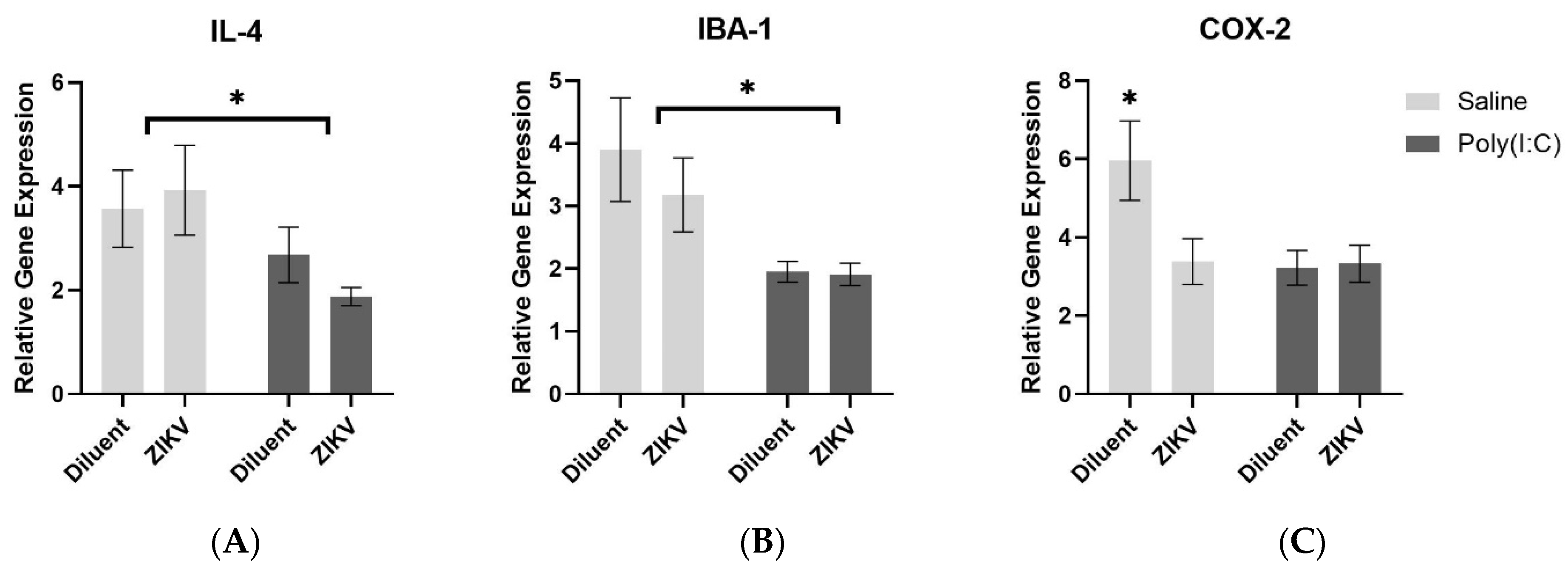
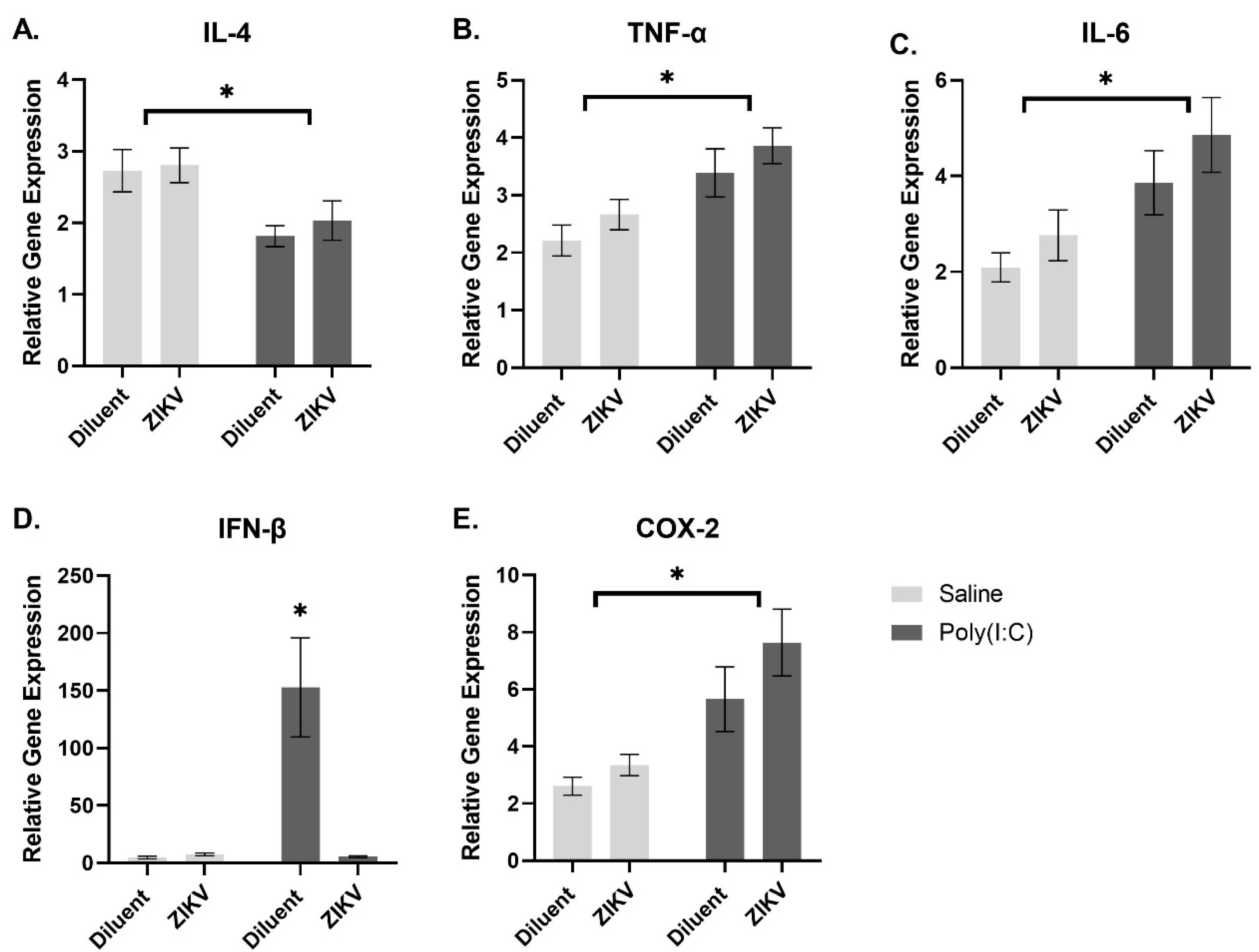
3.4.2. Analysis of Gene Expression following Treatment with Poly(I:C)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rombi, F.; Bayliss, R.; Tuplin, A.; Yeoh, S. The journey of Zika to the developing brain. Mol. Biol. Rep. 2020, 47, 3097–3115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cugola, F.R.; Fernandes, I.R.; Russo, F.B.; Freitas, B.C.; Dias, J.L.M.; Guimarães, K.P.; Benazzato, C.; Almeida, N.; Pignatari, G.C.; Romero, S.; et al. The Brazilian Zika virus strain causes birth defects in experimental models. Nature 2016, 534, 267–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Q.; Herrlinger, S.; Yang, S.-L.; Lai, F.; Moore, J.M.; Brindley, M.A.; Chen, J.-F. Zika virus infection disrupts neurovascular development and results in postnatal microcephaly with brain damage. Development 2016, 143, 4127–4136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, T.Z.; Haddad, L.B.; Williams, T.R.; Hills, S.L.; Read, J.S.; Dee, D.L.; Dziuban, E.J.; Pérez-Padilla, J.; Jamieson, D.J.; Honein, M.A.; et al. Breast milk transmission of flaviviruses in the context of Zika virus: A systematic review. Paediatr. Perinat. Epidemiol. 2018, 32, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Pomar, L.; Musso, D.; Malinger, G.; Vouga, M.; Panchaud, A.; Baud, D. Zika virus during pregnancy: From maternal exposure to congenital Zika virus syndrome. Prenat. Diagn. 2019, 39, 420–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devhare, P.; Meyer, K.; Steele, R.; Ray, R.B.; Ray, R. Zika virus infection dysregulates human neural stem cell growth and inhibits differentiation into neuroprogenitor cells. Cell Death Dis. 2017, 8, e3106. [Google Scholar] [CrossRef] [Green Version]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodušek, V.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Retallack, H.; Lullo, E.D.; Arias, C.; Knopp, K.A.; Laurie, M.T.; Sandoval-Espinosa, C.; Leon, W.R.M.; Krencik, R.; Ullian, E.M.; Spatazza, J.; et al. Zika virus cell tropism in the developing human brain and inhibition by azithromycin. Proc. Natl. Acad. Sci. USA 2016, 113, 14408–14413. [Google Scholar] [CrossRef] [Green Version]
- Gladwyn-Ng, I.; Cordón-Barris, L.; Alfano, C.; Creppe, C.; Couderc, T.; Morelli, G.; Thelen, N.; America, M.; Bessières, B.; Encha-Razavi, F.; et al. Stress-induced unfolded protein response contributes to Zika virus–associated microcephaly. Nat. Neurosci. USA 2018, 21, 63–71. [Google Scholar] [CrossRef]
- Moore, C.A.; Staples, J.E.; Dobyns, W.B.; Pessoa, A.; Ventura, C.V.; da Fonseca, E.B.; Ribeiro, E.M.; Ventura, L.O.; Neto, N.N.; Arena, J.F.; et al. Characterizing the Pattern of Anomalies in Congenital Zika Syndrome for Pediatric Clinicians. JAMA Pediatr. 2017, 171, 288–295. [Google Scholar] [CrossRef] [Green Version]
- Kapogiannis, B.G.; Chakhtoura, N.; Hazra, R.; Spong, C.Y. Bridging Knowledge Gaps to Understand How Zika Virus Exposure and Infection Affect Child Development. JAMA Pediatr. 2017, 171, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, A.C. Development of Infants With Congenital Zika Syndrome: What Do We Know and What Can We Expect? Pediatrics 2018, 141 (Suppl. S2), S154–S160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, J.F.; Pescarini, J.M.; de Souza Rodrigues, M.; de Araujo Almeida, B.; Henriques, C.M.P.; Gouveia, F.C.; Rabello, E.T.; Matta, G.C.; Barreto, M.L.; Sampaio, R.B. The global scientific research response to the public health emergency of Zika virus infection. PLoS ONE 2020, 15, e0229790. [Google Scholar] [CrossRef] [Green Version]
- Lazear, H.M.; Govero, J.; Smith, A.M.; Platt, D.J.; Fernandez, E.; Miner, J.J.; Diamond, M.S. A Mouse Model of Zika Virus Pathogenesis. Cell Host Microbe 2016, 19, 720–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Li, P.; Zhang, Y.; Liu, Y.; Tan, Z.; Sun, J.; Ke, X.; Miao, Y.; Luo, D.; Hu, Q.; et al. In vivo imaging of Zika virus reveals dynamics of viral invasion in immune-sheltered tissues and vertical propagation during pregnancy. Theranostics 2020, 10, 6430–6447. [Google Scholar] [CrossRef] [PubMed]
- Zukor, K.; Wang, H.; Siddharthan, V.; Julander, J.G.; Morrey, J.D. Zika virus-induced acute myelitis and motor deficits in adult interferon αβ/γ receptor knockout mice. J. Neurovirol. 2018, 24, 273–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, K.K.; Azouz, F.; Shin, O.S.; Kumar, M. Understanding the Pathogenesis of Zika Virus Infection Using Animal Models. Immune Netw. 2017, 17, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Morrison, T.E.; Diamond, M.S. Animal Models of Zika Virus Infection, Pathogenesis, and Immunity. J. Virol. 2017, 91, e00009-17. [Google Scholar] [CrossRef] [Green Version]
- Larocca, R.A.; Abbink, P.; Peron, J.P.S.; de A. Zanotto, P.M.; Iampietro, M.J.; Badamchi-Zadeh, A.; Boyd, M.; Ng’ang’a, D.; Kirilova, M.; Nityanandam, R.; et al. Vaccine protection against Zika virus from Brazil. Nature 2016, 536, 474–478. [Google Scholar] [CrossRef]
- Rossi, S.L.; Tesh, R.B.; Azar, S.R.; Muruato, A.E.; Hanley, K.A.; Auguste, A.J.; Langsjoen, R.M.; Paessler, S.; Vasilakis, N.; Weaver, S.C. Characterization of a Novel Murine Model to Study Zika Virus. Am. J. Trop. Med. Hyg. 2016, 94, 1362–1369. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.P.; Klein, S.L. Pregnancy and pregnancy-associated hormones alter immune responses and disease pathogenesis. Horm. Behav. 2012, 62, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Sherer, M.L.; Posillico, C.K.; Schwarz, J.M. The psychoneuroimmunology of pregnancy. Front. Neuroendocrinol. 2018, 51, 25–35. [Google Scholar] [CrossRef]
- Bilbo, S.D.; Schwarz, J.M. Early-life programming of later-life brain and behavior: A critical role for the immune system. Front. Behav. Neurosci. 2009, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, A.C.; Zaverucha-do-Valle, C.; Reis, P.A.; Barbosa-Lima, G.; Vieira, Y.R.; Mattos, M.; de Paiva Silva, P.; Sacramento, C.; de Castro Faria Neto, H.C.; Campanati, L.; et al. Sofosbuvir protects Zika virus-infected mice from mortality, preventing short- and long-term sequelae. Sci. Rep. 2017, 7, 9409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raper, J.; Kovacs-Balint, Z.; Mavigner, M.; Gumber, S.; Burke, M.W.; Habib, J.; Mattingly, C.; Fair, D.; Earl, E.; Feczko, E.; et al. Long-term alterations in brain and behavior after postnatal Zika virus infection in infant macaques. Nat. Commun. 2020, 11, 2534. [Google Scholar] [CrossRef]
- Zhao, Z.; Shang, Z.; Vasconcelos, Z.; Li, C.; Jiang, Y.; Zu, S.; Zhang, J.; Wang, F.; Yao, L.; Jung, J.U.; et al. Zika Virus Infection Leads to Variable Defects in Multiple Neurological Functions and Behaviors in Mice and Children. Adv. Sci. 2020, 7, 1901996. [Google Scholar] [CrossRef]
- Mavigner, M.; Raper, J.; Kovacs-Balint, Z.; Gumber, S.; O’Neal, J.T.; Bhaumik, S.K.; Zhang, X.; Habib, J.; Mattingly, C.; McDonald, C.E.; et al. Postnatal Zika virus infection is associated with persistent abnormalities in brain structure, function, and behavior in infant macaques. Sci. Transl. Med. 2018, 10, eaao6975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos, A.C.; Siqueira, M.; Alvarez-Rosa, L.; Cascabulho, C.M.; Waghabi, M.C.; Barbosa, H.S.; Adesse, D.; Stipursky, J. Toxoplasma gondii infection impairs radial glia differentiation and its potential to modulate brain microvascular endothelial cell function in the cerebral cortex. Microvasc. Res. 2020, 131, 104024. [Google Scholar] [CrossRef]
- Mutnal, M.B.; Cheeran, M.C.-J.; Hu, S.; Lokensgard, J.R. Murine Cytomegalovirus Infection of Neural Stem Cells Alters Neurogenesis in the Developing Brain. PLoS ONE 2011, 6, e16211. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Xu, D.; Ye, Q.; Hong, S.; Jiang, Y.; Liu, X.; Zhang, N.; Shi, L.; Qin, C.-F.; Xu, Z. Zika Virus Disrupts Neural Progenitor Development and Leads to Microcephaly in Mice. Cell Stem Cell 2016, 19, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Adams Waldorf, K.M.; Olson, E.M.; Nelson, B.R.; Little, M.-T.E.; Rajagopal, L. The Aftermath of Zika: Need for Long-Term Monitoring of Exposed Children. Trends Microbiol. 2018, 26, 729–732. [Google Scholar] [CrossRef]
- Christie, B.R.; Cameron, H.A. Neurogenesis in the adult hippocampus. Hippocampus 2006, 16, 199–207. [Google Scholar] [CrossRef]
- Emsley, J.G.; Mitchell, B.D.; Kempermann, G.; Macklis, J.D. Adult neurogenesis and repair of the adult CNS with neural progenitors, precursors, and stem cells. Prog. Neurobiol. 2005, 75, 321–341. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Saucedo-Cuevas, L.; Regla-Nava, J.A.; Chai, G.; Sheets, N.; Tang, W.; Terskikh, A.V.; Shresta, S.; Gleeson, J.G. Zika Virus Infects Neural Progenitors in the Adult Mouse Brain and Alters Proliferation. Cell Stem Cell 2016, 19, 593–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherer, M.L.; Khanal, P.; Talham, G.; Brannick, E.M.; Parcells, M.S.; Schwarz, J.M. Zika virus infection of pregnant rats and associated neurological consequences in the offspring. PLoS ONE 2019, 14, e0218539. [Google Scholar] [CrossRef] [PubMed]
- Hoeijmakers, L.; Lucassen, P.J.; Korosi, A. The interplay of early-life stress, nutrition, and immune activation programs adult hippocampal structure and function. Front. Mol. Neurosci. 2015, 7, 103. [Google Scholar] [CrossRef]
- Filiano, A.J.; Xu, Y.; Tustison, N.J.; Marsh, R.L.; Baker, W.; Smirnov, I.; Overall, C.C.; Gadani, S.P.; Turner, S.D.; Weng, Z.; et al. Unexpected role of interferon-γ in regulating neuronal connectivity and social behaviour. Nature 2016, 535, 425–429. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-L.; Wang, S. Prenatal lipopolysaccharide exposure increases depression-like behaviors and reduces hippocampal neurogenesis in adult rats. Behav. Brain Res. 2014, 259, 24–34. [Google Scholar] [CrossRef]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Careaga, M.; Taylor, S.L.; Chang, C.; Chiang, A.; Ku, K.M.; Berman, R.F.; Van de Water, J.A.; Bauman, M.D. Variability in PolyIC induced immune response: Implications for preclinical maternal immune activation models. J. Neuroimmunol. 2018, 323, 87–93. [Google Scholar] [CrossRef]
- Cunningham, C.; Campion, S.; Teeling, J.; Felton, L.; Perry, V.H. The sickness behaviour and CNS inflammatory mediator profile induced by systemic challenge of mice with synthetic double-stranded RNA (poly I:C). Brain. Behav. Immun. 2007, 21, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Huang, S.; Zhao, X.; Chen, M.; Lin, Y.; Xia, Y.; Sun, C.; Yang, X.; Wang, J.; Guo, Y.; et al. Poly(I:C) Treatment Leads to Interferon-Dependent Clearance of Hepatitis B Virus in a Hydrodynamic Injection Mouse Model. J. Virol. 2014, 88, 10421–10431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibney, S.M.; McGuinness, B.; Prendergast, C.; Harkin, A.; Connor, T.J. Poly I:C-induced activation of the immune response is accompanied by depression and anxiety-like behaviours, kynurenine pathway activation and reduced BDNF expression. Brain. Behav. Immun. 2013, 28, 170–181. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and Serologic Properties of Zika Virus Associated with an Epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.R.; Roby, J.A.; Dobyns, W.B.; Rajagopal, L.; Gale, M.; Adams Waldorf, K.M. Immune Evasion Strategies Used by Zika Virus to Infect the Fetal Eye and Brain. Viral Immunol. 2020, 33, 22–37. [Google Scholar] [CrossRef]
- Musso, D.; Ko, A.I.; Baud, D. Zika Virus Infection—After the Pandemic. N. Engl. J. Med. 2019, 381, 1444–1457. [Google Scholar] [CrossRef]
- Labouesse, M.A.; Langhans, W.; Meyer, U. Long-term pathological consequences of prenatal infection: Beyond brain disorders. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R1–R12. [Google Scholar] [CrossRef] [Green Version]
- Hagberg, H.; Mallard, C. Effect of inflammation on central nervous system development and vulnerability: Review. Curr. Opin. Neurol. 2005, 18, 117–123. [Google Scholar] [CrossRef]
- Snyder-Keller, A.; Kramer, L.D.; Zink, S.; Bolivar, V.J. Mouse Strain and Sex-Dependent Differences in Long-term Behavioral Abnormalities and Neuropathologies after Developmental Zika Infection. J. Neurosci. 2019, 39, 5393–5403. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Zou, P.; Chen, E.; Yao, H.; Zheng, H.; Wang, Q.; Zhu, J.-N.; Jiang, S.; Lu, L.; Zhang, J. Visual and Motor Deficits in Grown-up Mice with Congenital Zika Virus Infection. EBioMedicine 2017, 20, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Zeng, Y.; Lerner, A.; Gao, B.; Law, M. Nervous System Injury and Neuroimaging of Zika Virus Infection. Front. Neurol. 2018, 9, 227. [Google Scholar] [CrossRef] [PubMed]
- Klintsova, A.Y.; Helfer, J.L.; Calizo, L.H.; Dong, W.K.; Goodlett, C.R.; Greenough, W.T. Persistent Impairment of Hippocampal Neurogenesis in Young Adult Rats Following Early Postnatal Alcohol Exposure. Alcohol. Clin. Exp. Res. 2007, 31, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Fernandes, L.; Cugola, F.R.; Russo, F.B.; Kawahara, R.; de Melo Freire, C.C.; Leite, P.E.C.; Bassi Stern, A.C.; Angeli, C.B.; de Oliveira, D.B.L.; Melo, S.R.; et al. Zika Virus Impairs Neurogenesis and Synaptogenesis Pathways in Human Neural Stem Cells and Neurons. Front. Cell. Neurosci. 2019, 13, 64. [Google Scholar] [CrossRef] [PubMed]
- Meyer, U. Neurodevelopmental Resilience and Susceptibility to Maternal Immune Activation. Trends Neurosci. 2019, 42, 793–806. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Maternal immune activation: Implications for neuropsychiatric disorders. Science 2016, 353, 772–777. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (F) and Reverse (R) Primers |
|---|---|
| IFN-β | F: ATGGCCAACACGTGGACCCT R: TCAGTTCTGGAAGTTTCTAT |
| IL-4 | F: AAGGAACACCACGGAGAACG R: CAGACCGCTGACACCTCTAC |
| Iba1 | F: GAATGATGCTGGGCAAGAGA R: CAGTTGGCTTCTGGTGTTC |
| COX-2 | F: CTTCGCCTCTTTCAATGTGC R: GGTCAGTAGACTCTTACAGC |
| TNF-α | F: CTTCAAGGGACAAGGCTG R: GAGGCTGACTTTCTCCTG |
| ZIKV | F: CCGCTGCCCAACACAAG R: CCACTAACGTTCTTTTGCAGACAT Probe: FAM/AGCCTACCT/ZEN/TGACAAGCAATCAGACACTCAA/3IABkFQ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sherer, M.L.; Lemanski, E.A.; Patel, R.T.; Wheeler, S.R.; Parcells, M.S.; Schwarz, J.M. A Rat Model of Prenatal Zika Virus Infection and Associated Long-Term Outcomes. Viruses 2021, 13, 2298. https://doi.org/10.3390/v13112298
Sherer ML, Lemanski EA, Patel RT, Wheeler SR, Parcells MS, Schwarz JM. A Rat Model of Prenatal Zika Virus Infection and Associated Long-Term Outcomes. Viruses. 2021; 13(11):2298. https://doi.org/10.3390/v13112298
Chicago/Turabian StyleSherer, Morgan L., Elise A. Lemanski, Rita T. Patel, Shannon R. Wheeler, Mark S. Parcells, and Jaclyn M. Schwarz. 2021. "A Rat Model of Prenatal Zika Virus Infection and Associated Long-Term Outcomes" Viruses 13, no. 11: 2298. https://doi.org/10.3390/v13112298
APA StyleSherer, M. L., Lemanski, E. A., Patel, R. T., Wheeler, S. R., Parcells, M. S., & Schwarz, J. M. (2021). A Rat Model of Prenatal Zika Virus Infection and Associated Long-Term Outcomes. Viruses, 13(11), 2298. https://doi.org/10.3390/v13112298