
Genomic Epidemiology and Heterogeneity of SRLV in Italy from 1998 to 2019

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
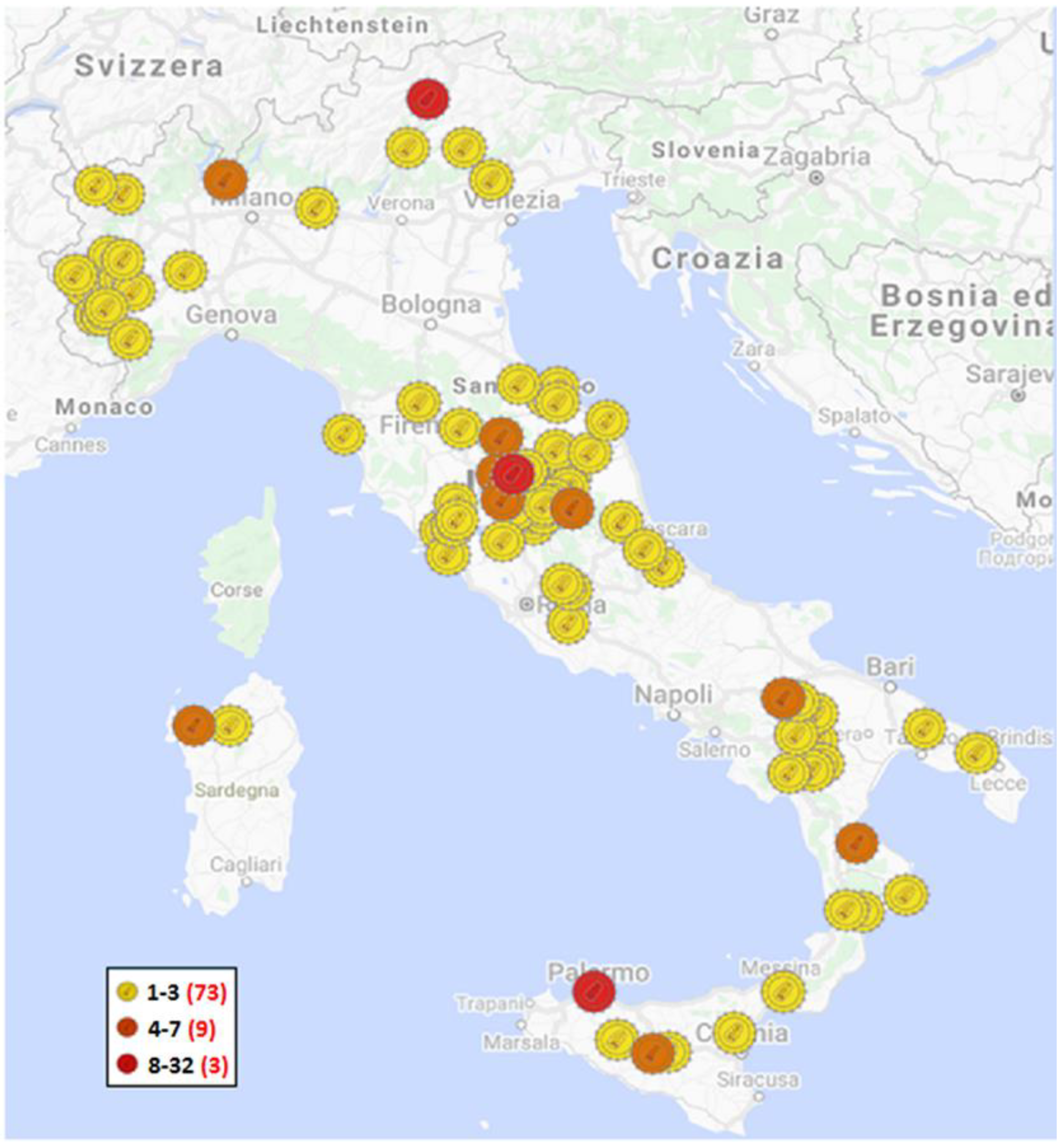
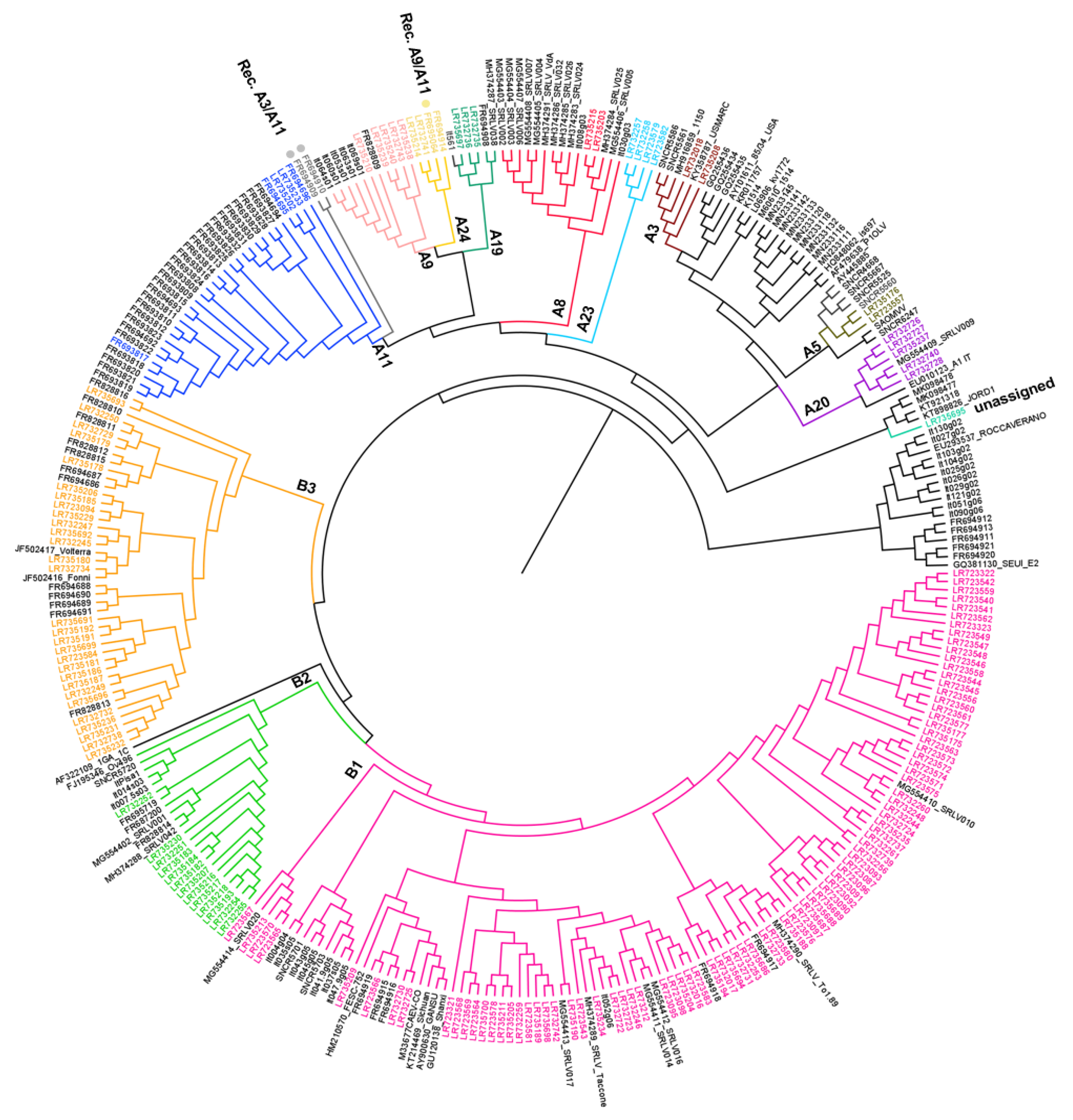
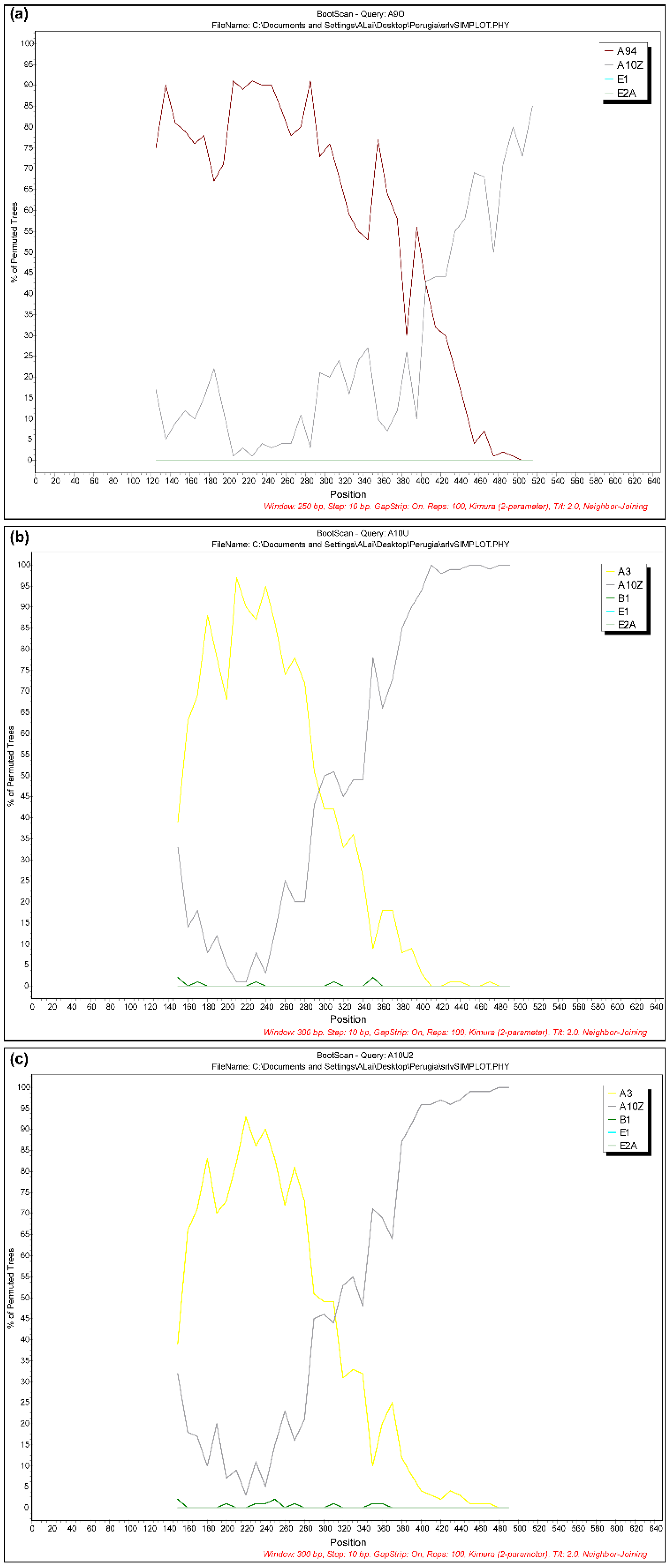
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blacklaws, B.A. Small Ruminant Lentiviruses: Immunopathogenesis of Visna-Maedi and Caprine Arthritis and Encephalitis Virus. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Leroux, C.; Minardi, C.J.; Mornex, J.-F. SRLVs: A Genetic Continuum of Lentiviral Species in Sheep and Goats with Cumulative Evidence of Cross Species Transmission. CHR 2010, 8, 94–100. [Google Scholar] [CrossRef]
- Ramírez, H.; Reina, R.; Amorena, B.; Andrés, D.; Martínez, H. Small Ruminant Lentiviruses: Genetic Variability, Tropism and Diagnosis. Viruses 2013, 5, 1175–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colitti, B.; Coradduzza, E.; Puggioni, G.; Capucchio, M.T.; Reina, R.; Bertolotti, L.; Rosati, S. A New Approach for Small Ruminant Lentivirus Full Genome Characterization Revealed the Circulation of Divergent Strains. PLoS ONE 2019, 14, e0212585. [Google Scholar] [CrossRef]
- Pritchard, G.C.; McConnel, I.; Aitken, I.D. Maedi -Visna. In Diseases of Sheep; Blackwell Publishing: Oxford, UK, 2007; pp. 217–223. [Google Scholar]
- Minguijón, E.; Reina, R.; Pérez, M.; Polledo, L.; Villoria, M.; Ramírez, H.; Leginagoikoa, I.; Badiola, J.J.; García-Marín, J.F.; de Andrés, D.; et al. Small Ruminant Lentivirus Infections and Diseases. Vet. Microbiol. 2015, 181, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Narayan, O.; Griffin, D.E.; Chase, J. Antigenic Shift of Visna Virus in Persistently Infected Sheep. Science 1977, 197, 376–378. [Google Scholar] [CrossRef]
- Narayan, O.; Griffin, D.E.; Silverstein, A.M. Slow Virus Infection: Replication and Mechanisms of Persistence of Visna Virus in Sheep. J. Infect. Dis. 1977, 135, 800–806. [Google Scholar] [CrossRef]
- Murphy, F.A.; Gibbs, E.P.J.; Horzinek, M.C.; Studdert, M.J. Veterinary Virology, III ed.; Academic Press: San Diego, CA, USA, 1999. [Google Scholar]
- Shah, C.; Böni, J.; Huder, J.B.; Vogt, H.-R.; Mühlherr, J.; Zanoni, R.; Miserez, R.; Lutz, H.; Schüpbach, J. Phylogenetic Analysis and Reclassification of Caprine and Ovine Lentiviruses Based on 104 New Isolates: Evidence for Regular Sheep-to-Goat Transmission and Worldwide Propagation through Livestock Trade. Virology 2004, 319, 12–26. [Google Scholar] [CrossRef] [Green Version]
- Cardinaux, L.; Zahno, M.-L.; Deubelbeiss, M.; Zanoni, R.; Vogt, H.-R.; Bertoni, G. Virological and Phylogenetic Characterization of Attenuated Small Ruminant Lentivirus Isolates Eluding Efficient Serological Detection. Vet. Microbiol. 2013, 162, 572–581. [Google Scholar] [CrossRef]
- de Andrés, X.; Ramírez, H.; Bertolotti, L.; San Román, B.; Glaria, I.; Crespo, H.; Jáuregui, P.; Minguijón, E.; Juste, R.; Leginagoikoa, I.; et al. An Insight into a Combination of ELISA Strategies to Diagnose Small Ruminant Lentivirus Infections. Vet. Immunol. Immunopathol. 2013, 152, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Molaee, V.; Bazzucchi, M.; De Mia, G.M.; Otarod, V.; Abdollahi, D.; Rosati, S.; Lühken, G. Phylogenetic Analysis of Small Ruminant Lentiviruses in Germany and Iran Suggests Their Expansion with Domestic Sheep. Sci. Rep. 2020, 10, 2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olech, M.; Valas, S.; Kuźmak, J. Epidemiological Survey in Single-Species Flocks from Poland Reveals Expanded Genetic and Antigenic Diversity of Small Ruminant Lentiviruses. PLoS ONE 2018, 13, e0193892. [Google Scholar] [CrossRef] [Green Version]
- Olech, M.; Murawski, M.; Kuźmak, J. Molecular Analysis of Small-Ruminant Lentiviruses in Polish Flocks Reveals the Existence of a Novel Subtype in Sheep. Arch. Virol. 2019, 164, 1193–1198. [Google Scholar] [CrossRef] [Green Version]
- Bertolotti, L.; Mazzei, M.; Puggioni, G.; Carrozza, M.L.; Dei Giudici, S.; Muz, D.; Juganaru, M.; Patta, C.; Tolari, F.; Rosati, S. Characterization of New Small Ruminant Lentivirus Subtype B3 Suggests Animal Trade within the Mediterranean Basin. J. Gen. Virol. 2011, 92, 1923–1929. [Google Scholar] [CrossRef] [PubMed]
- Giammarioli, M.; Bazzucchi, M.; Puggioni, G.; Brajon, G.; Dei Giudici, S.; Taccori, F.; Feliziani, F.; De Mia, G.M. Phylogenetic Analysis of Small Ruminant Lentivirus (SRLV) in Italian Flocks Reveals the Existence of Novel Genetic Subtypes. Virus Genes. 2011, 43, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Grego, E.; Bertolotti, L.; Quasso, A.; Profiti, M.; Lacerenza, D.; Muz, D.; Rosati, S. Genetic Characterization of Small Ruminant Lentivirus in Italian Mixed Flocks: Evidence for a Novel Genotype Circulating in a Local Goat Population. J. Gen. Virol. 2007, 88, 3423–3427. [Google Scholar] [CrossRef] [PubMed]
- Pisoni, G.; Quasso, A.; Moroni, P. Phylogenetic Analysis of Small-Ruminant Lentivirus Subtype B1 in Mixed Flocks: Evidence for Natural Transmission from Goats to Sheep. Virology 2005, 339, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Reina, R.; Bertolotti, L.; Dei Giudici, S.; Puggioni, G.; Ponti, N.; Profiti, M.; Patta, C.; Rosati, S. Small Ruminant Lentivirus Genotype E Is Widespread in Sarda Goat. Vet. Microbiol. 2010, 144, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Win-Dows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Lemey, P.; Lott, M.; Moulton, V.; Posada, D.; Lefeuvre, P. RDP3: A Flexible and Fast Computer Program for Analyzing Recombination. Bioinformatics 2010, 26, 2462–2463. [Google Scholar] [CrossRef] [PubMed]
- Peterhans, E.; Greenland, T.; Badiola, J.; Harkiss, G.; Bertoni, G.; Amorena, B.; Eliaszewicz, M.; Juste, R.A.; Kraßnig, R.; Lafont, J.-P.; et al. Routes of Transmission and Consequences of Small Ruminant Lentiviruses (SRLVs) Infection and Eradication Schemes. Vet. Res. 2004, 35, 257–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Martin, E.; Golomingi, A.; Zahno, M.; Cachim, J.; Di Labio, E.; Perler, L.; Abril, C.; Zanoni, R.; Bertoni, G. Diagnostic Response to a Cross-Border Challenge for the Swiss Caprine Arthritis Encephalitis Virus Eradication Program. SAT 2019, 161, 93–104. [Google Scholar] [CrossRef]
- Adjadj, N.R.; Vicca, J.; Michiels, R.; De Regge, N. (Non-)Sense of Milk Testing in Small Ruminant Lentivirus Control Programs in Goats. Comparative Analysis of Antibody Detection and Molecular Diagnosis in Blood and Milk. Viruses 2019, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Kampen, A.H.; Åkerstedt, J.; Klevar, S. The Surveillance Programme for Small Ruminant Lentivirus Infections in Sheep and Goats in Norway 2020; Norwegian Veterinary Institute: Oslo, Norway, 2021; ISSN 1890-3290. [Google Scholar]
- Turin, L.; Pisoni, G.; Giannino, M.L.; Antonini, M.; Rosati, S.; Ruffo, G.; Moroni, P. Correlation between Milk Parameters in CAEV Seropositive and Negative Primiparous Goats during an Eradication Program in Italian Farm. Small Rumin. Res. 2005, 57, 73–79. [Google Scholar] [CrossRef]
- Cirone, F.; Maggiolino, A.; Cirilli, M.; Sposato, A.; De Palo, P.; Ciappetta, G.; Pratelli, A. Small Ruminant Lentiviruses in Goats in Southern Italy: Serological Evidence, Risk Factors and Implementation of Control Programs. Vet. Microbiol. 2019, 228, 143–146. [Google Scholar] [CrossRef]
- Nardelli, S.; Bettini, A.; Capello, K.; Bertoni, G.; Tavella, A. Eradication of Caprine Arthritis Encephalitis Virus in the Goat Population of South Tyrol, Italy: Analysis of the Tailing Phenomenon during the 2016–2017 Campaign. J. VET Diagn. Investig. 2020, 32, 589–593. [Google Scholar] [CrossRef]
- Tavella, A.; Bettini, A.; Ceol, M.; Zambotto, P.; Stifter, E.; Kusstatscher, N.; Lombardi, R.; Nardeli, S.; Beato, M.S.; Capello, K.; et al. Achievements of an Eradication Programme against Caprine Arthritis Encephalitis Virus in South Tyrol, Italy. Vet. Rec. 2018, 182, 51. [Google Scholar] [CrossRef] [Green Version]
- de Andrés, D.; Klein, D.; Watt, N.J.; Berriatua, E.; Torsteinsdottir, S.; Blacklaws, B.A.; Harkiss, G.D. Diagnostic Tests for Small Ruminant Lentiviruses. Vet. Microbiol. 2005, 107, 49–62. [Google Scholar] [CrossRef]
- Pisoni, G.; Bertoni, G.; Manarolla, G.; Vogt, H.-R.; Scaccabarozzi, L.; Locatelli, C.; Moroni, P. Genetic Analysis of Small Ruminant Lentiviruses Following Lactogenic Transmission. Virology 2010, 407, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhar, U.; Barlič-Maganja, D.; Grom, J. Phylogenetic Analysis of Small Ruminant Lentiviruses Detected in Slovenia. Vet. Microbiol. 2013, 162, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Santry, L.A.; de Jong, J.; Gold, A.C.; Walsh, S.R.; Menzies, P.I.; Wootton, S.K. Genetic Characterization of Small Ruminant Lentiviruses Circulating in Naturally Infected Sheep and Goats in Ontario, Canada. Virus Res. 2013, 175, 30–44. [Google Scholar] [CrossRef]
- L’Homme, Y.; Leboeuf, A.; Arsenault, J.; Fras, M. Identification and Characterization of an Emerging Small Ruminant Lentivirus Circulating Recombinant Form (CRF). Virology 2015, 475, 159–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michiels, R.; Adjadj, N.R.; De Regge, N. Phylogenetic Analysis of Belgian Small Ruminant Lentiviruses Supports Cross Species Virus Transmission and Identifies New Subtype B5 Strains. Pathogens 2020, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Gjerset, B.; Jonassen, C.M.; Rimstad, E. Natural Transmission and Comparative Analysis of Small Ruminant Lentiviruses in the Norwegian Sheep and Goat Populations. Virus Res. 2007, 125, 153–161. [Google Scholar] [CrossRef]
- Olech, M.; Kuźmak, J. Molecular Characterization of Small Ruminant Lentiviruses of Subtype A5 Detected in Naturally Infected but Clinically Healthy Goats of Carpathian Breed. Pathogens 2020, 9, 992. [Google Scholar] [CrossRef]
- Hu, W.-S.; Temin, H.M. Retroviral Recombination and Reverse Transcription. Science 1990, 250, 1227–1233. [Google Scholar] [CrossRef]
- Fras, M.; Leboeuf, A.; Labrie, F.-M.; Laurin, M.-A.; Singh Sohal, J.; L’Homme, Y. Phylogenetic Analysis of Small Ruminant Lentiviruses in Mixed Flocks: Multiple Evidence of Dual Infection and Natural Transmission of Types A2 and B1 between Sheep and Goats. Infect. Genet. Evol. 2013, 19, 97–104. [Google Scholar] [CrossRef]
- Jolly, P.E.; Narayan, O. Evidence for Interference, Coinfections and Intertypic Virus Enhancement of Infection by Ovine-Caprine Lentiviruses. J. Virol. 1989, 63, 4682–4688. [Google Scholar] [CrossRef] [Green Version]
- Pisoni, G.; Bertoni, G.; Puricelli, M.; Maccalli, M.; Moroni, P. Demonstration of Coinfection with and Recombination by Caprine Arthritis-Encephalitis Virus and Maedi-Visna Virus in Naturally Infected Goats. J. Virol. 2007, 81, 4948–4955. [Google Scholar] [CrossRef] [Green Version]
- Pisoni, G.; Moroni, P.; Turin, L.; Bertoni, G. Compartmentalization of Small Ruminant Lentivirus between Blood and Colostrum in Infected Goats. Virology 2007, 369, 119–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, H.; Glaria, I.; de Andrés, X.; Martínez, H.A.; Hernández, M.M.; Reina, R.; Iráizoz, E.; Crespo, H.; Berriatua, E.; Vázquez, J.; et al. Recombinant Small Ruminant Lentivirus Subtype B1 in Goats and Sheep of Imported Breeds in Mexico. Vet. J. 2011, 190, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, H.; Reina, R.; Bertolotti, L.; Cenoz, A.; Hernández, M.-M.; San Román, B.; Glaria, I.; de Andrés, X.; Crespo, H.; Jáuregui, P.; et al. Study of Compartmentalization in the Visna Clinical Form of Small Ruminant Lentivirus Infection in Sheep. BMC Vet. Res. 2012, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Blatti-Cardinaux, L.; Pisoni, G.; Stoffel, M.H.; Zanoni, R.; Zahno, M.-L.; Bertoni, G. Generation of a Molecular Clone of an Attenuated Lentivirus, a First Step in Understanding Cytopathogenicity and Virulence. Virology 2016, 487, 50–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazzucchi, M.; Pierini, I.; Gobbi, P.; Pirani, S.; Torresi, C.; Iscaro, C.; Feliziani, F.; Giammarioli, M. Genomic Epidemiology and Heterogeneity of SRLV in Italy from 1998 to 2019. Viruses 2021, 13, 2338. https://doi.org/10.3390/v13122338
Bazzucchi M, Pierini I, Gobbi P, Pirani S, Torresi C, Iscaro C, Feliziani F, Giammarioli M. Genomic Epidemiology and Heterogeneity of SRLV in Italy from 1998 to 2019. Viruses. 2021; 13(12):2338. https://doi.org/10.3390/v13122338
Chicago/Turabian StyleBazzucchi, Moira, Ilaria Pierini, Paola Gobbi, Silvia Pirani, Claudia Torresi, Carmen Iscaro, Francesco Feliziani, and Monica Giammarioli. 2021. "Genomic Epidemiology and Heterogeneity of SRLV in Italy from 1998 to 2019" Viruses 13, no. 12: 2338. https://doi.org/10.3390/v13122338
APA StyleBazzucchi, M., Pierini, I., Gobbi, P., Pirani, S., Torresi, C., Iscaro, C., Feliziani, F., & Giammarioli, M. (2021). Genomic Epidemiology and Heterogeneity of SRLV in Italy from 1998 to 2019. Viruses, 13(12), 2338. https://doi.org/10.3390/v13122338