
Nudivirus Sequences Identified from the Southern and Western Corn Rootworms (Coleoptera: Chrysomelidae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Collection, DNA Isolation, Library Preparation and Illumina Sequencing
2.2. Sequence Assembly
2.3. Identification of Contigs and Viral Sequence Discovery
2.4. Viral Sequence Analysis and Annotation
3. Results
3.1. Assembly of Genomic Sequence Data for Nudiviral Sequence Identification
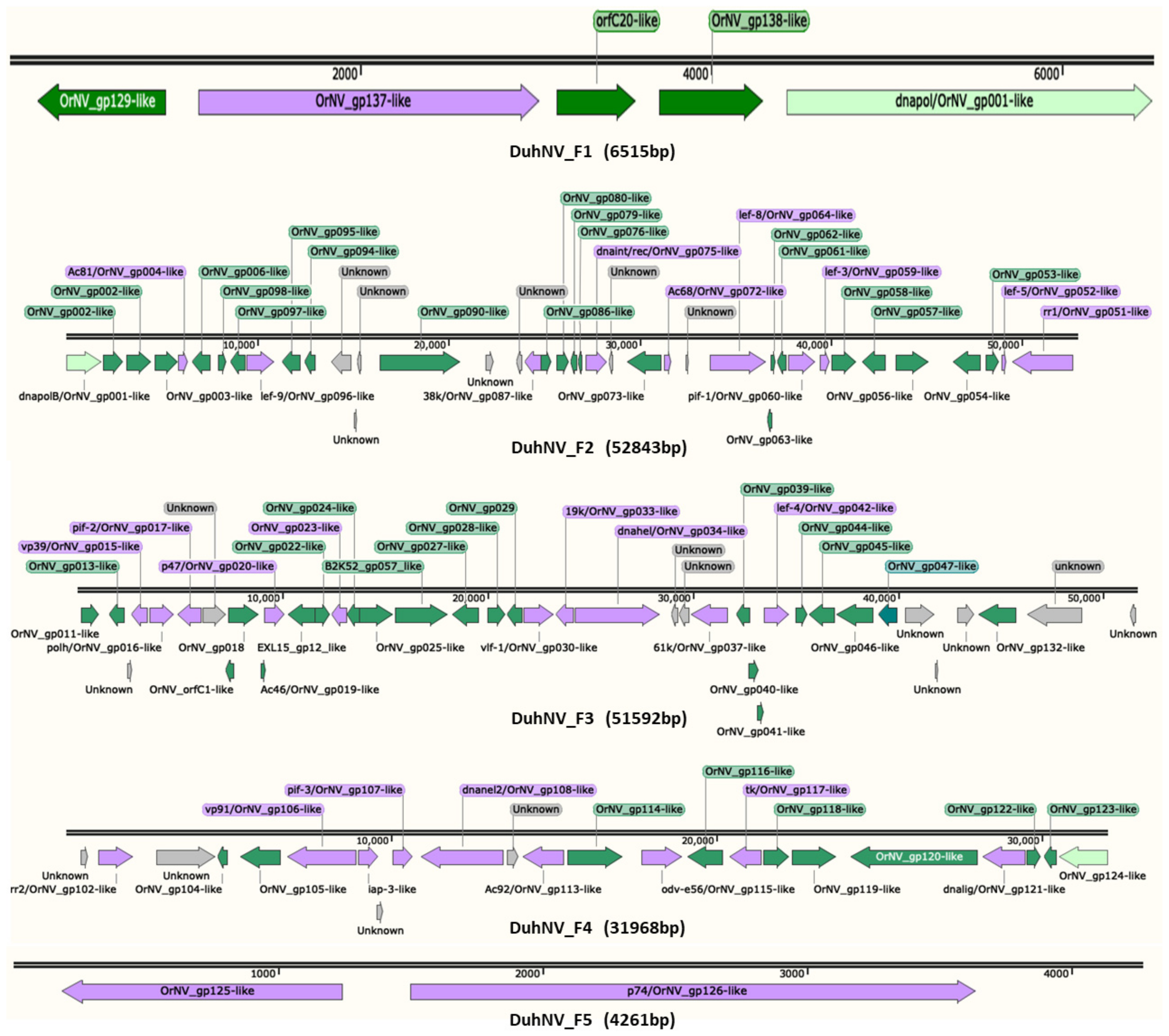
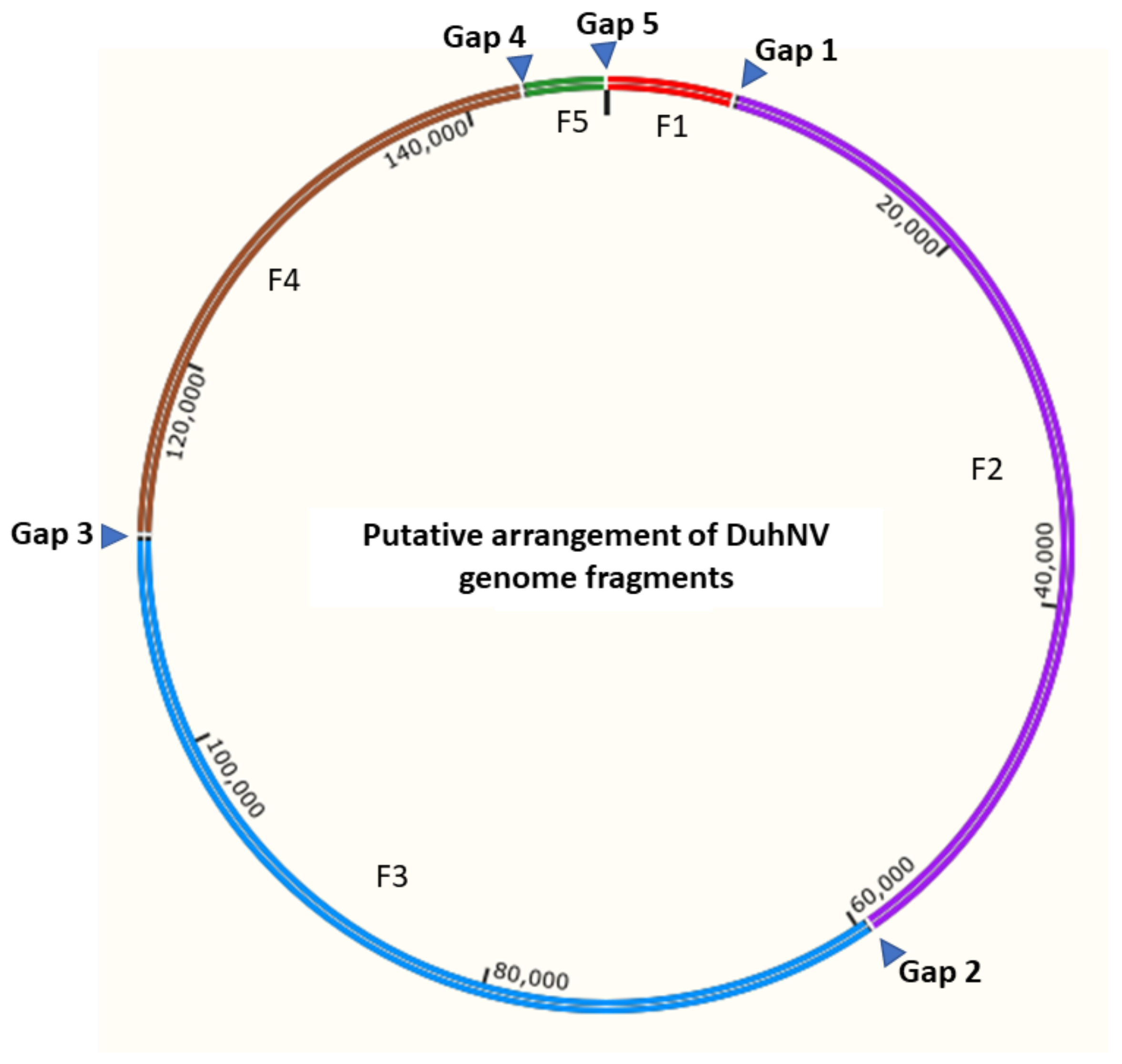
3.2. Sequence Analysis of Diabrotica undecimpunctata Howardi Nudivirus (DuhNV)
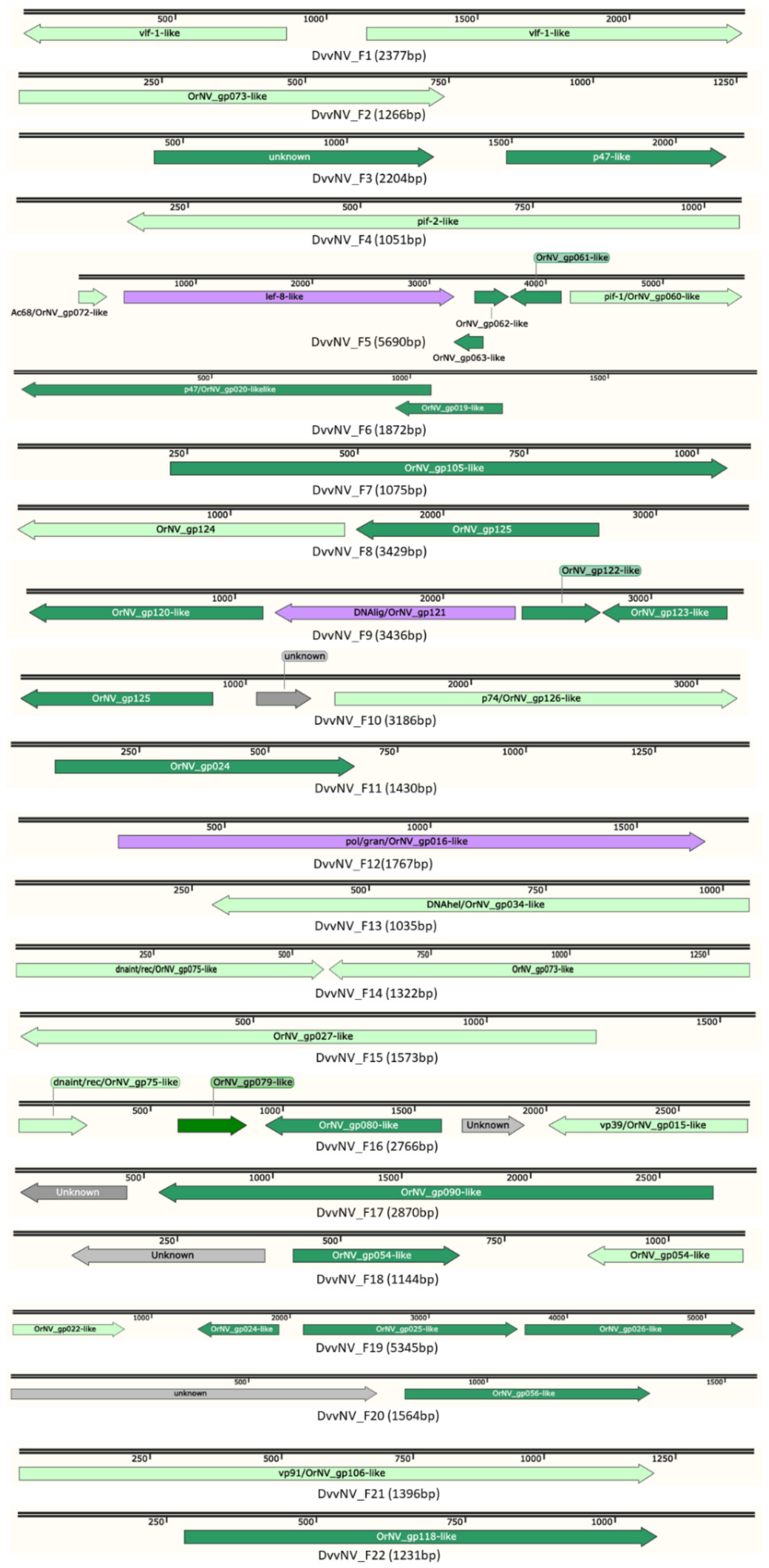
3.3. Analysis of Diabrotica virgifera virgifera Nudivirus Sequences
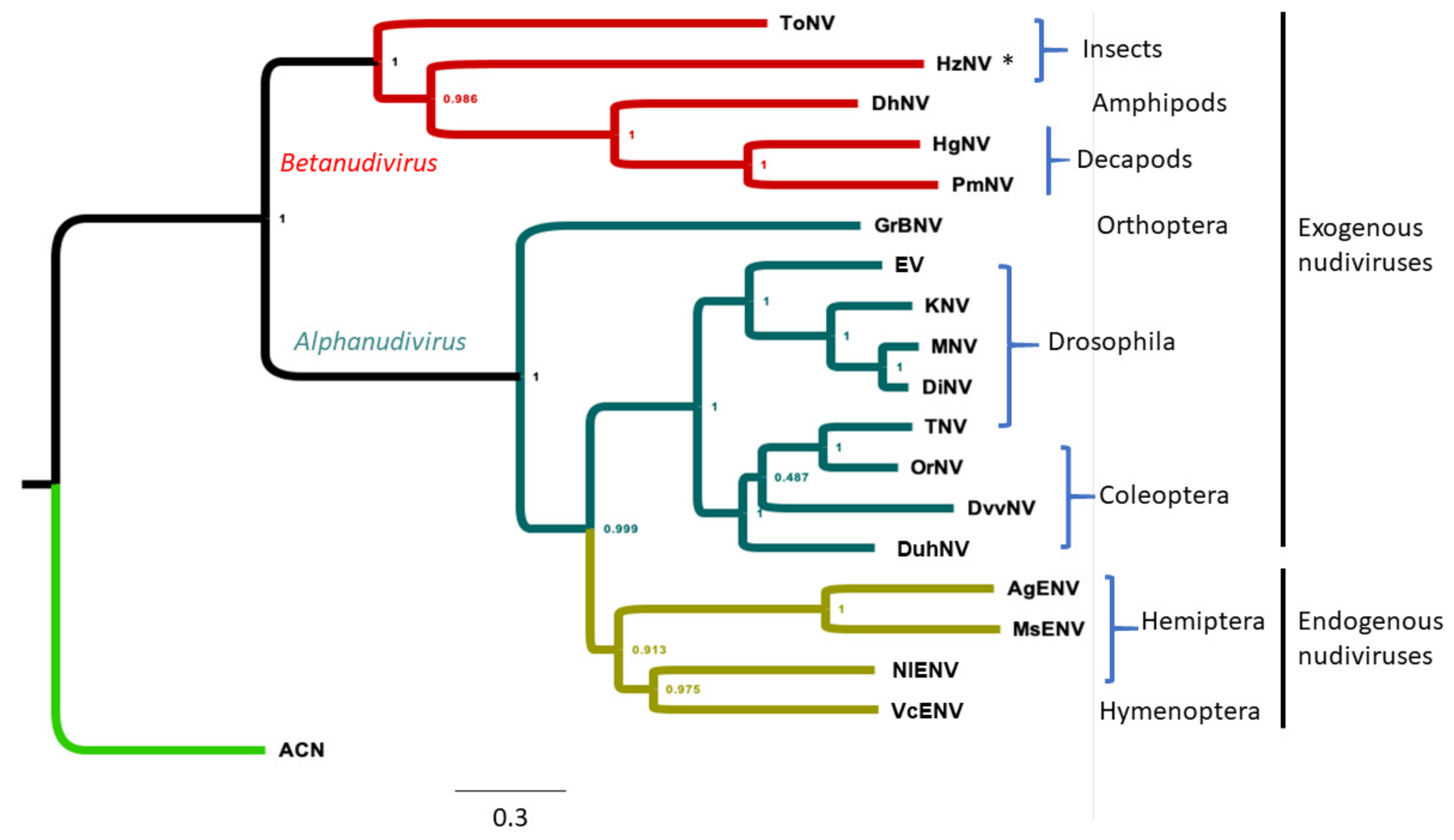
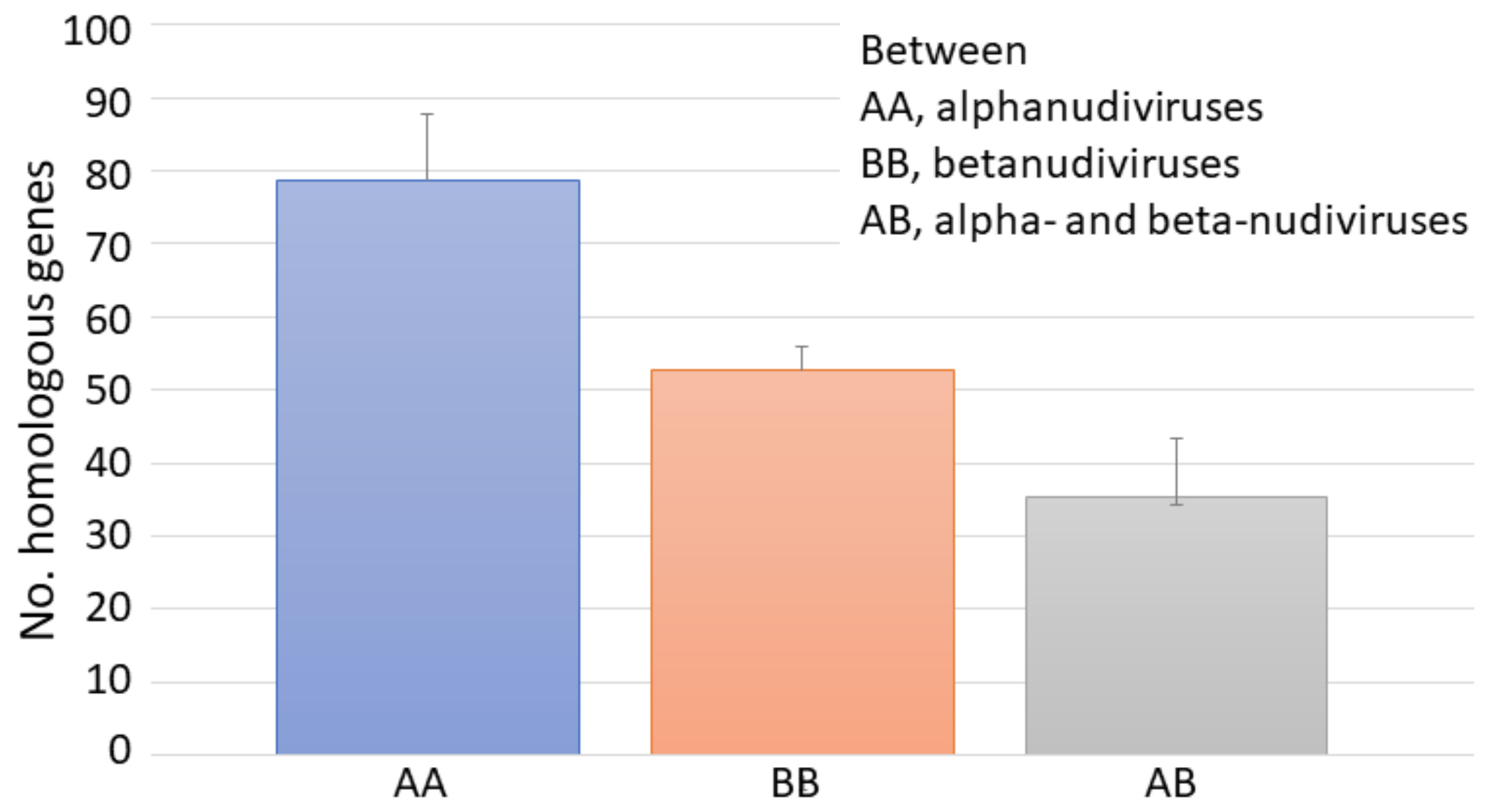
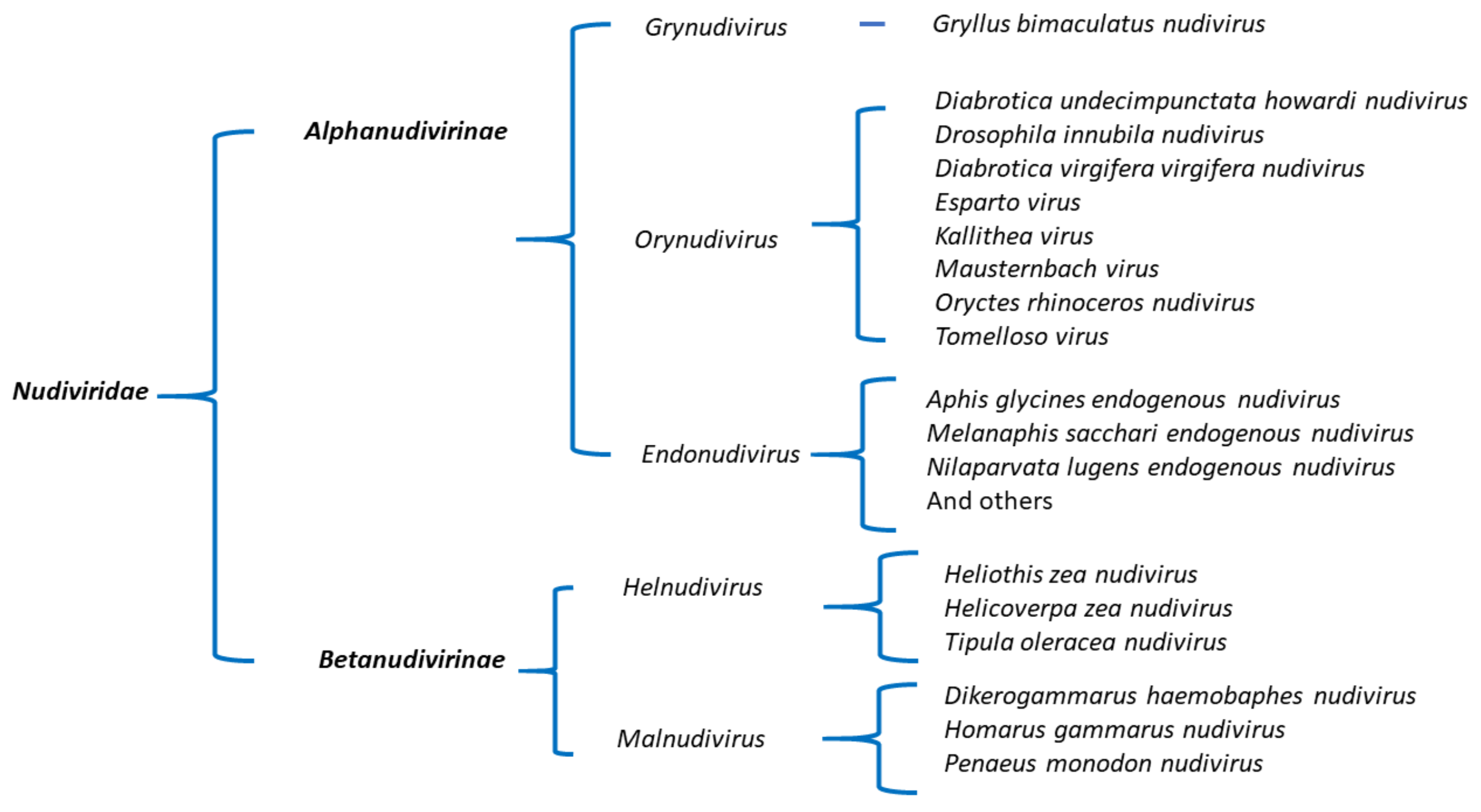
3.4. Phylogenetic Analysis of WCR Nudiviruses and Other Nudiviruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Author Notes
References
- Cabrera Walsh, G.; Ávila, C.J.; Cabrera, N.; Nava, D.E.; de Sene Pinto, A.; Weber, D.C. Biology and management of pest Diabrotica species in South America. Insects 2020, 11, 421. [Google Scholar] [CrossRef]
- Krysan, J.L.; Smith, R.F. Systematics of the virgifera species group of Diabrotica (Coleoptera: Chrysomelidae: Galerucinae). Entomography 1987, 5, 375–484. [Google Scholar]
- Wechsler, S.; Smith, D. Has resistance taken root in U.S. corn fields? Demand for insect control. Am. J. Agric. Econ. 2018, 100, 1136–1150. [Google Scholar] [CrossRef]
- Souza, D.; Peterson, J.A.; Wright, R.J.; Meinke, L.J. Field efficacy of soil insecticides on pyrethroid-resistant western corn rootworms (Diabrotica virgifera virgifera LeConte). Pest Manag. Sci. 2020, 76, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, A.J.; Petzold-Maxwell, J.L.; Keweshan, R.S.; Dunbar, M.W. Field-evolved resistance to Bt maize by western corn rootworm. PLoS ONE 2011, 6, e22629. [Google Scholar]
- Gassmann, A.J. Resistance to Bt maize by western corn rootworm: Insights from the laboratory and the field. Curr. Opin. Insect Sci. 2016, 15, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, A.J.; Shrestha, R.B.; Kropf, A.L.; St Clair, C.R.; Brenizer, B.D. Field-evolved resistance by western corn rootworm to Cry34/35Ab1 and other Bacillus thuringiensis traits in transgenic maize. Pest Manag. Sci. 2020, 76, 268–276. [Google Scholar] [CrossRef]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Price, D.R.; Gatehouse, J.A. RNAi-mediated crop protection against insects. Trends Biotechnol. 2008, 26, 393–400. [Google Scholar] [CrossRef]
- Fishilevich, E.; Vélez, A.M.; Storer, N.P.; Li, H.; Bowling, A.J.; Rangasamy, M.; Worden, S.E.; Narva, K.E.; Siegfried, B.D. RNAi as a management tool for the western corn rootworm, Diabrotica virgifera virgifera. Pest Manag. Sci. 2016, 72, 1652–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khajuria, C.; Ivashuta, S.; Wiggins, E.; Flagel, L.; Moar, W.; Pleau, M.; Miller, K.; Zhang, Y.; Ramaseshadri, P.; Jiang, C.; et al. Development and characterization of the first dsRNA-resistant insect population from western corn rootworm, Diabrotica virgifera virgifera LeConte. PLoS ONE 2018, 13, e0197059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velez, A.M.; Fishilevich, E.; Rangasamy, M.; Khajuria, C.; McCaskill, D.G.; Pereira, A.E.; Gandra, P.; Frey, M.L.; Worden, S.E.; Whitlock, S.L.; et al. Control of western corn rootworm via RNAi traits in maize: Lethal and sublethal effects of Sec23 dsRNA. Pest Manag. Sci. 2020, 76, 1500–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedford, G.O. Biology and management of palm dynastid beetles: Recent advances. Annu. Rev. Entomol. 2013, 58, 353–372. [Google Scholar] [CrossRef]
- Jackson, T.A.; Crawford, A.M.; Glare, T.R. Oryctes virus--time for a new look at a useful biocontrol agent. J. Invertebr. Pathol. 2005, 89, 91–94. [Google Scholar] [CrossRef]
- Degrugillier, M.E.; Degrugillier, S.S.; Jackson, J.J. Nonoccluded, Cytoplasmic Virus-Particles and Rickettsia-Like Organisms in Testes and Spermathecae of Diabrotica-virgifera. J. Invertebr. Pathol. 1991, 57, 50–58. [Google Scholar] [CrossRef]
- Kim, K.S.; Kitajima, E.W. Nonoccluded Baculovirus-Like and Filamentous Virus-Like Particles in the Spotted Cucumber Beetle, Diabrotica-Undecimpunctata (Coleoptera, Chrysomelid). J. Invertebr. Pathol. 1984, 43, 234–241. [Google Scholar] [CrossRef]
- Kim, K.S. Cytopathology of spotted cucumber beetle hemocytes containing virus-like particles. J. Invertebr. Pathol. 1980, 36, 292–301. [Google Scholar] [CrossRef]
- Kim, K.S.; Scott, H.A.; Fulton, J.P. Viruses discovered in beetle pests. Ark. Farm Res. 1981, 30. [Google Scholar]
- Harrison, R.L.; Herniou, E.A.; Bézier, A.; Jehle, J.A.; Burand, J.P.; Theilmann, D.A.; Krell, P.J.; van Oers, M.M.; Nakai, M. and ICTV Report ConsortiumICTV Virus Taxonomy Profile: Nudiviridae. J. Gen. Virol. 2020, 101, 3–4. [Google Scholar] [CrossRef]
- Huger, A.M.; Krieg, A. Baculoviridae. Nonoccluded Baculoviruses. In Atlas of Invertebrate Viruses; Adams, J.R., Bonami, J.R., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1991; pp. 287–319. [Google Scholar]
- Cheng, R.L.; Li, X.F.; Zhang, C.X. Nudivirus Remnants in the Genomes of Arthropods. Genome Biol. Evol. 2020, 12, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Drezen, J.M.; Leobold, M.; Bézier, A.; Huguet, E.; Volkoff, A.N.; Herniou, E.A. Endogenous viruses of parasitic wasps: Variations on a common theme. Curr. Opin. Virol. 2017, 25, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Leobold, M.; Bézier, A.; Pichon, A.; Herniou, E.A.; Volkoff, A.N.; Drezen, J.M. The Domestication of a Large DNA Virus by the Wasp Venturia canescens Involves Targeted Genome Reduction through Pseudogenization. Genome Biol. Evol. 2018, 10, 1745–1764. [Google Scholar] [CrossRef] [PubMed]
- Pichon, A.; Bézier, A.; Urbach, S.; Aury, J.M.; Jouan, V.; Ravallec, M.; Guy, J.; Cousserans, F.; Thézé, J.; Gauthier, J. Recurrent DNA virus domestication leading to different parasite virulence strategies. Sci. Adv. 2015, 1, e1501150. [Google Scholar] [CrossRef] [Green Version]
- Bézier, A.; Annaheim, M.; Herbinière, J.; Wetterwald, C.; Gyapay, G.; Bernard-Samain, S.; Wincker, P.; Roditi, I.; Heller, M.; Belghazi, M.; et al. Polydnaviruses of braconid wasps derive from an ancestral nudivirus. Science 2009, 323, 926–930. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Coates, B.S.; Bonning, B.C. Endogenous viral elements integrated into the genome of the soybean aphid, Aphis glycines. Insect Biochem. Mol. Biol. 2020, 123, 103405. [Google Scholar]
- Cheng, R.L.; Xi, Y.; Lou, Y.H.; Wang, Z.; Xu, J.Y.; Xu, H.J.; Zhang, C.X. Brown Planthopper Nudivirus DNA Integrated in Its Host Genome. J. Virol. 2014, 88, 5310–5318. [Google Scholar] [CrossRef] [Green Version]
- Huger, A.M. A new virus-disease of crickets (Orthoptera, Gryllidae) causing macronucleosis of fat-body. J. Invertebr. Pathol. 1985, 45, 108–111. [Google Scholar] [CrossRef]
- Wang, Y.; Kleespies, R.G.; Huger, A.M.; Jehle, J.A. The genome of Gryllus bimaculatus nudivirus indicates an ancient diversification of baculovirus-related nonoccluded nudiviruses of insects. J. Virol. 2007, 81, 5395–5406. [Google Scholar] [CrossRef] [Green Version]
- Palmer, W.H.; Medd, N.C.; Beard, P.M.; Obbard, D.J. Isolation of a natural DNA virus of Drosophila melanogaster, and characterisation of host resistance and immune responses. PLoS Pathog. 2018, 14. [Google Scholar] [CrossRef] [PubMed]
- Unckless, R.L. A DNA Virus of Drosophila. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huger, A.M. A virus disease of Indian rhinoceros beetle Oryctes rhinoceros (Linnaeus) caused by a new type of insect virus Rhabdionvirus oryctes Gen N sp. N. J. Invertebr. Pathol. 1966, 8, 38–51. [Google Scholar] [CrossRef]
- Wang, Y.; Bininda-Emonds, O.R.; van Oers, M.M.; Vlak, J.M.; Jehle, J.A. The genome of Oryctes rhinoceros nudivirus provides novel insight into the evolution of nuclear arthropod-specific large circular double-stranded DNA viruses. Virus Genes 2011, 42, 444–456. [Google Scholar] [CrossRef]
- Granados, R.R. Early events in the infection of Heliothis zea midgut cells by a baculovirus. Virology 1978, 90, 170–174. [Google Scholar] [CrossRef]
- Cheng, C.H.; Liu, S.M.; Chow, T.Y.; Hsiao, Y.Y.; Wang, D.P.; Huang, J.J.; Chen, H.H. Analysis of the complete genome sequence of the Hz-1 virus suggests that it is related to members of the Baculoviridae. J. Virol. 2002, 76, 9024–9034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raina, A.K.; Adams, J.R. Gonad-specific virus of corn-earworm. Nature 1995, 374, 770. [Google Scholar] [CrossRef]
- Burand, J.P.; Kim, W.; Afonso, C.L.; Tulman, E.R.; Kutish, G.F.; Lu, Z.; Rock, D.L. Analysis of the genome of the sexually transmitted insect virus Helicoverpa zea nudivirus 2. Viruses 2012, 4, 28–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, C.C.; Stone, M.; Bass, D.; Bateman, K.S.; van Aerle, R.; Daniels, C.L.; van der Giezen, M.; Ross, S.H.; Hooper, C.; Stentiford, G.D. The first clawed lobster virus Homarus gammarus nudivirus (HgNV n. sp.) expands the diversity of the Nudiviridae. Sci. Rep. 2019, 9, 10086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.T.; Lee, D.Y.; Wang, Y.; Hu, J.M.; Li, W.H.; Leu, J.H.; Chang, G.D.; Ke, H.M.; Kang, S.T.; Lin, S.S.; et al. The genome and occlusion bodies of marine Penaeus monodon nudivirus (PmNV, also known as MBV and PemoNPV) suggest that it should be assigned to a new nudivirus genus that is distinct from the terrestrial nudiviruses. BMC Genomics 2014, 15, 628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bézier, A.; Thézé, J.; Gavory, F.; Gaillard, J.; Poulain, J.; Drezen, J.M.; Herniou, E.A. The genome of the nucleopolyhedrosis-causing virus from Tipula oleracea sheds new light on the Nudiviridae family. J. Virol. 2015, 89, 3008–3025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allain, T.W.; Stentiford, G.D.; Bass, D.; Behringer, D.C.; Bojko, J. A novel nudivirus infecting the invasive demon shrimp Dikerogammarus haemobaphes (Amphipoda). Sci. Rep. 2020, 10, 14816. [Google Scholar] [CrossRef] [PubMed]
- Bonning, B.C. The Insect Virome: Opportunities and Challenges. Curr. Issues Mol. Biol. 2020, 34, 1–12. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Y.; Sappington, T.W.; Bonning, B.C. Genome Sequence of a Novel Positive-Sense, Single-Stranded RNA Virus Isolated from Western Corn Rootworm, Diabrotica virgifera virgifera LeConte. Genome Announc. 2017, 5, e00366-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Chen, Y.; Sappington, T.W.; Bonning, B.C. Genome Sequence of Diabrotica virgifera virgifera virus2, a Novel Small RNA Virus of the Western Corn Rootworm, Diabrotica virgifera virgifera LeConte. Genome Announc. 2017, 5, e00365-17. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Chen, Y.; Sappington, T.W.; Bonning, B.C. Genome Sequence of the First Coleopteran Iflavirus Isolated from Western Corn Rootworm, Diabrotica virgifera virgifera LeConte. Genome Announc. 2017, 5, e01530-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Valencia-Jiménez, A.; Darlington, M.; Vélez, A.M.; Bonning, B.C. Diabrotica undecimpunctata virus 2, a Novel Small RNA Virus Discovered from Southern Corn Rootworm, Diabrotica undecimpunctata howardi Barber (Coleoptera: Chrysomelidae). Microbiol. Resour Announc. 2020, 9, e00380-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Valencia-Jiménez, A.; Darlington, M.; Vélez, A.M.; Bonning, B.C. Genome Sequence of a Small RNA Virus of the Southern Corn Rootworm, Diabrotica undecimpunctata howardi Barber (Coleoptera: Chrysomelidae). Microbiol. Resour Announc. 2020, 9, e00379-20. [Google Scholar] [CrossRef]
- Coates, B.S. Assembly and annotation of full mitochondrial genomes for the corn rootworm species, Diabrotica virgifera virgifera and Diabroticabarberi (Insecta: Coleoptera: Chrysomelidae), using Next Generation Sequence data. Gene 2014, 542, 190–197. [Google Scholar] [CrossRef]
- Flagel, L.E.; Bansal, R.; Kerstetter, R.A.; Chen, M.; Carroll, M.; Flannagan, R.; Clark, T.; Goldman, B.S.; Michel, A.P. Western corn rootworm (Diabrotica virgifera virgifera) transcriptome assembly and genomic analysis of population structure. BMC Genomics 2014, 15, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program. for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Geer, L.Y.; Domrachev, M.; Lipman, D.J.; Bryant, S.H. CDART: Protein homology by domain architecture. Genome Res. 2002, 12, 1619–1623. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.J.; Jehle, J.A. Nudiviruses and other large, double-stranded circular DNA viruses of invertebrates: New insights on an old topic. J. Invertebr. Pathol. 2009, 101, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Cepas, J.; Serra, F.; Bork, P. ETE 3: Reconstruction, Analysis, and Visualization of Phylogenomic Data. Molec. Biol. Evol. 2016, 33, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Webster, C.L.; Waldron, F.M.; Robertson, S.; Crowson, D.; Ferrari, G.; Quintana, J.F.; Brouqui, J.M.; Bayne, E.H.; Longdon, B.; Buck, A.H.; et al. The Discovery, Distribution, and Evolution of Viruses Associated with Drosophila melanogaster. PLoS Biol. 2015, 13, e1002210. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, E.L.; Jaszczyszyn, Y.; Naquin, D.; Thermes, C. The Third Revolution in Sequencing Technology. Trends Genet. 2018, 34, 666–681. [Google Scholar] [CrossRef]
- Drees, B.M. Southern corn rootworm (spotted cucumber beetle). In Handbook of Corn Insects; Steffey, K., Rice, M., All, J., Andow, D., Gray, M., Van Duyn, J.., Eds.; Entomological Society of America: Lanham, MD, USA, 1999; p. 63. [Google Scholar]
- Sappington, T.W.; Hesler, L.S.; Allen, K.C.; Luttrell, R.G.; Papiernik, S.K. Prevalence of sporadic insect pests of seedling corn, and factors affecting risk of infestation. J. Integr. Pest Manag. 2018, 9, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Gray, M.E.; Sappington, T.W.; Miller, N.J.; Moeser, J.; Bohn, M.O. Adaptation and invasiveness of western corn rootworm: Intensifying research on a worsening pest. Annu. Rev. Entomol. 2009, 54, 303–321. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Abbr. | Length (bp) | Accession | No. ORFs | %G + C | Host | Isolates | Ref |
|---|---|---|---|---|---|---|---|---|
| Alphanudivirus | ||||||||
| Gryllus bimaculatus nudivirus | GrBNV | 96,944 | NC_009240.1 | 98 | 27.99 | Gryllus bimaculatus | Starnberg, Germany | [28,29] |
| Kallithea virus | KNV | 152,388 | NC_033829.1 | 95 | 32.88 | Drosophila melanogaster | Kharkiv, Ukraine | [30] |
| Drosophila innubila nudivirus | DiNV | 155,555 | NC_040699.1 | 106 | 30.29 | Drosophila innubila | Arizona, USA | [31] |
| Oryctes rhinoceros nudivirus | OrNV | 127,615 | EU747721.1 | 139 | 41.63 | Oryctes rhinoceros | Malaysia | [32,33] |
| Esparto virus | EV | 183,261 | NC_040536.1 | 87 | 29.45 | Drosophila melanogaster | Esparto, CA, USA | NA |
| Mauternbach virus | MNV | 154,465 | MG969167.1 | 93 | 30.67 | Drosophila melanogaster | DrosEU50 Mauternbach 2015, Austria | NA |
| Tomelloso virus | TNV | 112,307 | NC_040789.1 | 93 | 39.63 | Drosophila melanogaster | DrosEU28 Tomelloso 2015, Spain | NA |
| Betanudivirus | ||||||||
| Heliothis zea nudivirus 1 | HzNV1 | 228,089 | AF451898.1 | 154 | 41.81 | Heliothis zea | Taiwan, PRC | [34,35] |
| Helicoverpa zea nudivirus 2 | HzNV2 | 231,621 | NC_004156.2 | 113 | 41.88 | Helicoverpa zea | Massachusetts, USA | [36,37] |
| Homarus gammarus nudivirus | HgNV | 107,063 | MK439999.1 | 97 | 35.03 | Homarus gammarus | 52S104HLG2, hepatopancreas, UK | [38] |
| Penaeus monodon nudivirus | PmNV | 119,638 | NC_024692.1 | 115 | 34.53 | Penaeus monodon | Indonesia | [39] |
| Tipula oleracea nudivirus | ToNV | 145,704 | NC_026242.1 | 131 | 25.53 | Tipula oleracea | England, UK | [40] |
| Dikerogammarus haemobaphes nudivirus | DhNV | 119,754 | MT488302 | 106 | Dikerogammarus haemobaphes | UK | [41] | |
| Unclassified | ||||||||
| Allomyrina virus | NA | AIY68660 | NA | NA | Allomyrina dichotoma | Youngdong-1, Korea | NA |
| Species | Sequencing Dataset | U.S. Collection Site | Total Reads (M) | Trimmed Reads (M) | Average Quality Score (q) | N50 (bp) | No. of Contigs (≥200) | No. of Contigs (≥1000) |
|---|---|---|---|---|---|---|---|---|
| D. undecimpunctata | SRR13364002 | Ames, IA | 9.7 | 9.7 | 37 | 420 | 33,539 | NA |
| D. virgifera virgifera | SRR1107649 | Adams County, IN | 195 | 184 | 37 | 403 | 4.4 M | 84,419 |
| D. virgifera virgifera | SRR1107695 | Seneca, KS | 194 | 186 | 38 | 414 | 4.1 M | 71,902 |
| D. virgifera virgifera | SRR1107698 | Colfax, NE | 41 | 40 | 37 | 422 | 3.8 M | 97,060 |
| Fragment | Length (bp) | G + C (%) | No. Putative CDS | Total Reads Mapped | Total Bases Mapped | Read Coverage |
|---|---|---|---|---|---|---|
| F1 | 6515 | 28.49 | 5 | 1210 | 121,000 | 18.6 |
| F2 | 52,843 | 27.93 | 40 | 10,280 | 1,028,000 | 19.5 |
| F3 | 51,592 | 27.88 | 40 | 9858 | 985,800 | 19.1 |
| F4 | 31,968 | 27.56 | 23 | 6202 | 620,200 | 19.4 |
| F5 | 4261 | 29.46 | 2 | 687 | 68,700 | 16.1 |
| Total | 147,179 | 109 | 28,237 | 2,823,700 | ||
| Average | 28.264 | 18.5 |
| ORF | Gene/Domain | Length (aa) | OrNV Homologs |
|---|---|---|---|
| Replication, Repair and Recombination of DNA | |||
| DuhNV_F1_ORF5 * | Dnapol | 695 | OrNV_gp001 |
| DuhNV_F2_ORF1 * | Dnapol | 633 | OrNV_gp001 |
| DuhNV_F3_ORF23 | Dnahel | 1390 | OrNV_gp034 |
| DuhNV_F2_ORF23 | dnaint/rec | 389 | OrNV_gp075 |
| DuhNV_F2_ORF33 | lef-3 | 177 | OrNV_gp059 |
| DuhNV_F4_ORF10 | dnahel2 | 839 | OrNV_gp108 |
| DuhNV_F4_ORF20 | Dnalig | 423 | OrNV_gp121 |
| Nucleotide Metabolism | |||
| DuhNV_F2_ORF40 | rr1 | 1045 | OrNV_gp051 |
| DuhNV_F4_ORF2 | rr2 | 358 | OrNV_gp102 |
| DuhNV_F1_ORF2 | tk/GrBNV_gp17-like | 648 | OrNV_gp137 |
| DuhNV_F3_ORF31 | tk/GrBNV-gp74_like | 197 | OrNV_gp044 |
| DuhNV_F4_ORF16 | tk(guanosine monophosphate kinase) | 321 | OrNV_gp117 |
| DuhNV_F5_ORF1 | tk/GrBNV_gp044_like | 352 | OrNV_gp125 |
| Transcription | |||
| DuhNV_F3_ORF11 | p47 | 324 | OrNV_gp020 |
| DuhNV_F3_ORF30 | lef-4 | 417 | OrNV_gp042 |
| DuhNV_F2_ORF39 | lef-5 | 82 | OrNV_gp052 |
| DuhNV_F2_ORF28 | lef-8 | 988 | OrNV_gp064 |
| DuhNV_F2_ORF9 | lef-9 | 505 | OrNV_gp096 |
| DuhNV_F3_ORF21 | vlf-1 | 504 | OrNV_gp030 |
| Oral Infectivity | |||
| DuhNV_F5_ORF2 | p74 | 712 | OrNV_gp126 |
| DuhNV_F2_ORF32 | pif-1 | 489 | OrNV_gp060 |
| DuhNV_F3_ORF6 | pif2 | 376 | OrNV_gp017 |
| DuhNV_F4_ORF9 | pif-3 | 202 | OrNV_gp107 |
| Packaging, Assembly, and Morphogenesis | |||
| DuhNV_F3_ORF5 | polh/gran | 404 | OrNV_gp016 |
| DuhNV_F3_ORF22 | 19k/pif-5 | 281 | OrNV_gp033 |
| DuhNV_F2_ORF26 | Ac68 | 136 | OrNV_gp072 |
| DuhNV_F2_ORF18 | 38k | 276 | OrNV_gp087 |
| Inhibition of Apoptosis | |||
| DuhNV_F4_ORF7 | iap-3 | 202 | OrNV_gp134 |
| Unknown Function | |||
| DuhNV_F2_ORF5 | Ac81 | 173 | OrNV_gp004 |
| DuhNV_F3_ORF4 | vp39 | 250 | OrNV_gp015 |
| DuhNV_F4_ORF6 | vp91 | 695 | OrNV_gp106 |
| DuhNV_F4_ORF12 | Ac92 | 410 | OrNV_gp113 |
| DuhNV_F4_ORF14 | odv-e56 | 420 | OrNV_gp115 |
| Other | |||
| DuhNV_F1_ORF1 | ND | 240 | OrNV_gp129/gp136 |
| DuhNV_F1_ORF3 | ND | 149 | OrNV_orfC20 |
| DuhNV_F1_ORF4 | ND | 594 | OrNV_gp138 |
| DuhNV_F2_ORF2 | trypsin-like serine protease-like protein | 339 | OrNV_gp002 |
| DuhNV_F2_ORF3 | trypsin-like serine protease-like protein | 427 | OrNV_gp002 |
| DuhNV_F2_ORF4 | ND | 395 | OrNV_gp003 |
| DuhNV_F2_ORF6 | thymidylate synthase/pyrimidine hydroxymethylase-like | 296 | OrNV_gp006 |
| DuhNV_F2_ORF7 | ND | 143 | OrNV_gp098 |
| DuhNV_F2_ORF8 | mRNA decapping enzyme 2-like | 245 | OrNV_gp097 |
| DuhNV_F2_ORF10 | ND | 314 | OrNV_gp095 |
| DuhNV_F2_ORF11 | ND | 162 | OrNV_gp094 |
| DuhNV_F2_ORF12 | cl17238, RING-Ubox | 329 | no hit |
| DuhNV_F2_ORF13 | None | 62 | no hit |
| DuhNV_F2_ORF14 | None | 63 | no hit |
| DuhNV_F2_ORF15 1 | cl25745, pfam05764 | 1412 | OrNV_gp090 |
| DuhNV_F2_ORF16 | None | 148 | no hit |
| DuhNV_F2_ORF17 | None | 78 | no hit |
| DuhNV_F2_ORF19 | ND | 189 | OrNV_gp086 |
| DuhNV_F2_ORF20 | ND | 227 | OrNV_gp080 |
| DuhNV_F2_ORF21 | ND | 89 | OrNV_gp079 |
| DuhNV_F2_ORF22 | ND | 52 | OrNV_gp076 |
| DuhNV_F2_ORF24 | None | 61 | no hit |
| DuhNV_F2_ORF25 | ND | 593 | OrNV_gp073 |
| DuhNV_F2_ORF27 | None | 56 | no hit |
| DuhNV_F2_ORF29 | ND | 74 | OrNV_gp063 |
| DuhNV_F2_ORF30 | ND | 88 | OrNV_gp062 |
| DuhNV_F2_ORF31 | ND | 140 | OrNV_gp061 |
| DuhNV_F2_ORF34 | HZV_115-like protein | 425 | OrNV_gp058 |
| DuhNV_F2_ORF35 | patatin-like phospholipase-like protein | 397 | OrNV_gp057 |
| DuhNV_F2_ORF36 | ND | 576 | OrNV_gp056 |
| DuhNV_F2_ORF37 | ND | 458 | OrNV_gp054 |
| DuhNV_F2_ORF38 | ND | 234 | OrNV_gp053 |
| DuhNV_F3_ORF1 | mitochondrial carrier protein-like protein | 293 | OrNV_gp011 |
| DuhNV_F3_ORF2 | ND | 231 | OrNV_gp013 |
| DuhNV_F3_ORF3 | None | 76 | no hit |
| DuhNV_F3_ORF7 | None | 388 | no hit |
| DuhNV_F3_ORF8 | ND | 131 | OrNV_orfC1 |
| DuhNV_F3_ORF9 | ND | 509 | OrNV_gp018 |
| DuhNV_F3_ORF10 | Ac46 | 89 | OrNV_gp019 |
| DuhNV_F3_ORF12 2 | EXL15_gp12_like/DUF4679 (pfam15728) | 432 | no hit |
| DuhNV_F3_ORF13 | ND | 241 | OrNV_gp022 |
| DuhNV_F3_ORF14 | guanylate kinase-like protein | 245 | OrNV_gp023 |
| DuhNV_F3_ORF15 | ND | 196 | OrNV_gp024 |
| DuhNV_F3_ORF16 | ND | 550 | OrNV_gp025 |
| DuhNV_F3_ORF17 3 | B2K52_gp057/Smc cl341474 | 858 | no hit |
| DuhNV_F3_ORF18 | ND | 422 | OrNV_gp027 |
| DuhNV_F3_ORF19 4 | PPK13561/cl32896, EAL domain containing protein | 297 | OrNV_gp028 |
| DuhNV_F3_ORF20 | ND | 232 | OrNV_gp029 |
| DuhNV_F3_ORF24 | None | 96 | no hit |
| DuhNV_F3_ORF25 | None | 157 | no hit |
| DuhNV_F3_ORF26 | 61k AcORF9 | 575 | OrNV_gp037 |
| DuhNV_F3_ORF27 | ND | 199 | OrNV_gp039 |
| DuhNV_F3_ORF28 | ND | 166 | OrNV_gp040 |
| DuhNV_F3_ORF29 | ND | 110 | OrNV_gp041 |
| DuhNV_F3_ORF32 | ND | 403 | OrNV_gp045 |
| DuhNV_F3_ORF33 | ND | 591 | OrNV_gp046 |
| DuhNV_F3_ORF34 | ND | 293 | OrNV_gp047 |
| DuhNV_F3_ORF35 5 | cl34121, energy-coupling factor ABC transporter | 490 | no hit |
| DuhNV_F3_ORF36 | None | 66 | no hit |
| DuhNV_F3_ORF37 | None | 278 | no hit |
| DuhNV_F3_ORF38 | ND | 592 | OrNV_gp132 |
| DuhNV_F3_ORF39 | None | 868 | no hit |
| DuhNV_F3_ORF40 | None | 85 | no hit |
| DuhNV_F4_ORF1 | None | 76 | no hit |
| DuhNV_F4_ORF3 | None | 616 | no hit |
| DuhNV_F4_ORF4 | ND | 91 | OrNV_gp104 |
| DuhNV_F4_ORF5 | ND | 408 | OrNV_gp105 |
| DuhNV_F4_ORF8 | None | 68 | no hit |
| DuhNV_F4_ORF11 | None | 120 | no hit |
| DuhNV_F4_ORF13 | ND | 566 | OrNV_gp114 |
| DuhNV_F4_ORF15 | ND | 345 | OrNV_gp116 |
| DuhNV_F4_ORF17 | ND | 266 | OrNV_gp118 |
| DuhNV_F4_ORF18 | ND | 454 | OrNV_gp119 |
| DuhNV_F4_ORF19 | ND | 1285 | OrNV_gp120 |
| DuhNV_F4_ORF21 | ND | 143 | OrNV_gp122 |
| DuhNV_F4_ORF22 | ND | 124 | OrNV_gp123 |
| DuhNV_F4_ORF23 | ND | 476 * | OrNV_gp124 |
| ORF | Length (Some Partial) | Gene | Similar OrNV ORF | Similar DuhNV ORF |
|---|---|---|---|---|
| DvvNV_F1_ORF1 | 414 | vlf-1 | OrNV_gp030 | DuhNV_F3_ORF21 |
| DvvNV_F1_ORF2 | 289 | vlf-1 | OrNV_gp030 | DuhNV_F3_ORF21 |
| DvvNV_F2_ORF1 | 246 | OrNV_gp073-like | OrNV_gp073 | DuhNV_F2_ORF25 |
| DvvNV_F3_ORF1 | 223 | p47 | OrNV_gp020 | DuhNV_F3_ORF11 |
| DvvNV_F3_ORF2 | 284 | no hit | DuhNV_F3_ORF9 | |
| DvvNV_F4_ORF1 | 293 | pif-2 | OrNV_gp017 | DuhNV_F3_ORF6 |
| DvvNV_F5_ORF1 | 82 | Ac68-like | OrNV_gp072 | no hit |
| DvvNV_F5_ORF2 | 943 | lef-8 | OrNV_gp064 | DuhNV_F2_ORF28 |
| DvvNV_F5_ORF3 | 81 | OrNV_gp063-like | OrNV_gp063 | no hit |
| DvvNV_F5_ORF4 | 98 | OrNV_gp062-like | OrNV_gp062 | DuhNV_F2_ORF30 |
| DvvNV_F5_ORF5 | 144 | GrBNV_gp51-like | OrNV_gp061 | no hit |
| DvvNV_F5_ORF6 | 493 | pif-1 | OrNV_gp060 | DuhNV_F2_ORF32 |
| DvvNV_F6_ORF1 | 343 | p47 | OrNV_gp020 | DuhNV_F3_ORF11 |
| DvvNV_F6_ORF2 | 89 | Ac146-like | OrNV_gp019 | DuhNV_F3_ORF10 |
| DvvNV_F7_ORF1 | 272 | GrBNV gp43-like | OrNV_gp105 | DuhNV_F4_ORF5 |
| DvvNV_F8_ORF1 | 379 | GrBNV_gp44-like | OrNV_gp125 | DuhNV_F5_ORF1 |
| DvvNV_F8_ORF2 | 512 | OrNV_gp124-like | OrNV_gp124 | DuhNV_F4_ORF23 |
| DvvNV_F9_ORF1 | 373 | GrBNVgp37-like | OrNV_gp120 | DuhNV_F4_ORF19 |
| DvvNV_F9_ORF2 | 384 | DNAlig | OrNV_gp121 | DuhNV_F4_ORF20 |
| DvvNV_F9_ORF3 | 125 | GrBNV_gp39-like | OrNV_gp122 | no hit |
| DvvNV_F9_ORF4 | 199 | GrBNVgp41-like | OrNV_gp123 | DuhNV_F4_ORF22 |
| DvvNV_F10_ORF1 | 284 | GrBNV_gp44-like | OrNV_gp125 | DuhNV_F5_ORF1 |
| DvvNV_F10_ORF2 | 80 | unknown | no hit | |
| DvvNV_F10_ORF3 | 596 | p74 | OrNV_gp126 | DuhNV_F5_ORF2 |
| DvvNV_F11_ORF1 | 193 | GrBNVgp75-like | OrNV_gp024 | DuhNV_F3_ORF15 |
| DvvNV_F12_ORF1 | 474 | pol/gran | OrNV_gp016 | DuhNV_F3_ORF5 |
| DvvNV_F13_ORF1 | 251 | DNAhel | _OrNV_gp034 | DuhNV_F3_ORF23 |
| DvvNV_F14_ORF1 | 184 | dnaint/rec | OrNV_gp075 | DuhNV_F2_ORF23 |
| DvvNV_F14_ORF2 | 251 | putative gene | OrNV_gp073 | DuhNV_F2_ORF25 |
| DvvNV_F15_ORF1 | 411 | GrBNV_gp78-like | OrNV_gp027 | DuhNV_F3_ORF18 |
| DvvNV_F16_ORF1 | 86 | dnaint/rec | OrNV_gp075 | DuhNV_F2_ORF23 |
| DvvNV_F16_ORF2 | 222 | GrBNVgp60-like | OrNV_gp080 | no hit |
| DvvNV_F16_ORF3 | 86 | GrBNVgp59-like | OrNV_gp079 | DuhNV_F2_ORF21 |
| DvvNV_F16_ORF4 | 79 | unknown | ||
| DvvNV_F16_ORF5 | 250 | vp39 | OrNV_gp015 | DuhNV_F3_ORF4 |
| DvvNV_F17_ORF1 | 715 | GrBNVgp28-like | OrNV_gp090 | DuhNV_F2_ORF15 |
| DvvNV_F17_ORF2 | 136 | unknown | ||
| DvvNV_F18_ORF1 | 84 | GrBNV gp83-like | OrNV_gp054 | DuhNV_F2_ORF37 |
| DvvNV_F18_ORF2 | 97 | unknown | DuhNV_F2_ORF37 | |
| DvvNV_F18_ORF3 | 78 | GrBNV gp83-like | OrNV_gp054 | no hit |
| DvvNV_F19_ORF1 | 270 | GrBNV_gp72-like | OrNV_gp022 | DuhNV_F3_ORF13 |
| DvvNV_F19_ORF2 | 516 | GrBNV_gp76-like | OrNV_gp025 | DuhNV_F3_ORF16 |
| DvvNV_F19_ORF3 | 526 | unknown | OrNV_gp026 | DuhNV_F3_ORF17 |
| DvvNV_F19_ORF4 | 193 | GrBNVgp75-like | OrNV_gp024 | DuhNV_F3_ORF15 |
| DvvNV_F20_ORF1 | 171 | OrNV_gp056 | OrNV_gp056 | DuhNV_F2_ORF36 |
| DvvNV_F20_ORF2 | 256 | unknown | ||
| DvvNV_F21_ORF1 | 402 | vp91 | OrNV_gp106 | DuhNV_F4_ORF6 |
| DvvNV_F22_ORF1 | 263 | GrBNV_gp35-like | OrNV_gp118 | DuhNV_F4_ORF17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Sappington, T.W.; Coates, B.S.; Bonning, B.C. Nudivirus Sequences Identified from the Southern and Western Corn Rootworms (Coleoptera: Chrysomelidae). Viruses 2021, 13, 269. https://doi.org/10.3390/v13020269
Liu S, Sappington TW, Coates BS, Bonning BC. Nudivirus Sequences Identified from the Southern and Western Corn Rootworms (Coleoptera: Chrysomelidae). Viruses. 2021; 13(2):269. https://doi.org/10.3390/v13020269
Chicago/Turabian StyleLiu, Sijun, Thomas W. Sappington, Brad S. Coates, and Bryony C. Bonning. 2021. "Nudivirus Sequences Identified from the Southern and Western Corn Rootworms (Coleoptera: Chrysomelidae)" Viruses 13, no. 2: 269. https://doi.org/10.3390/v13020269
APA StyleLiu, S., Sappington, T. W., Coates, B. S., & Bonning, B. C. (2021). Nudivirus Sequences Identified from the Southern and Western Corn Rootworms (Coleoptera: Chrysomelidae). Viruses, 13(2), 269. https://doi.org/10.3390/v13020269