
Characterization of the Mycovirome of the Phytopathogenic Fungus, Neofusicoccum parvum

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Neofusicoccum parvum Isolate COLB Identification and Growth Conditions
2.2. Total RNA Extraction, High Throughput Sequencing, and Data Sequence Analysis
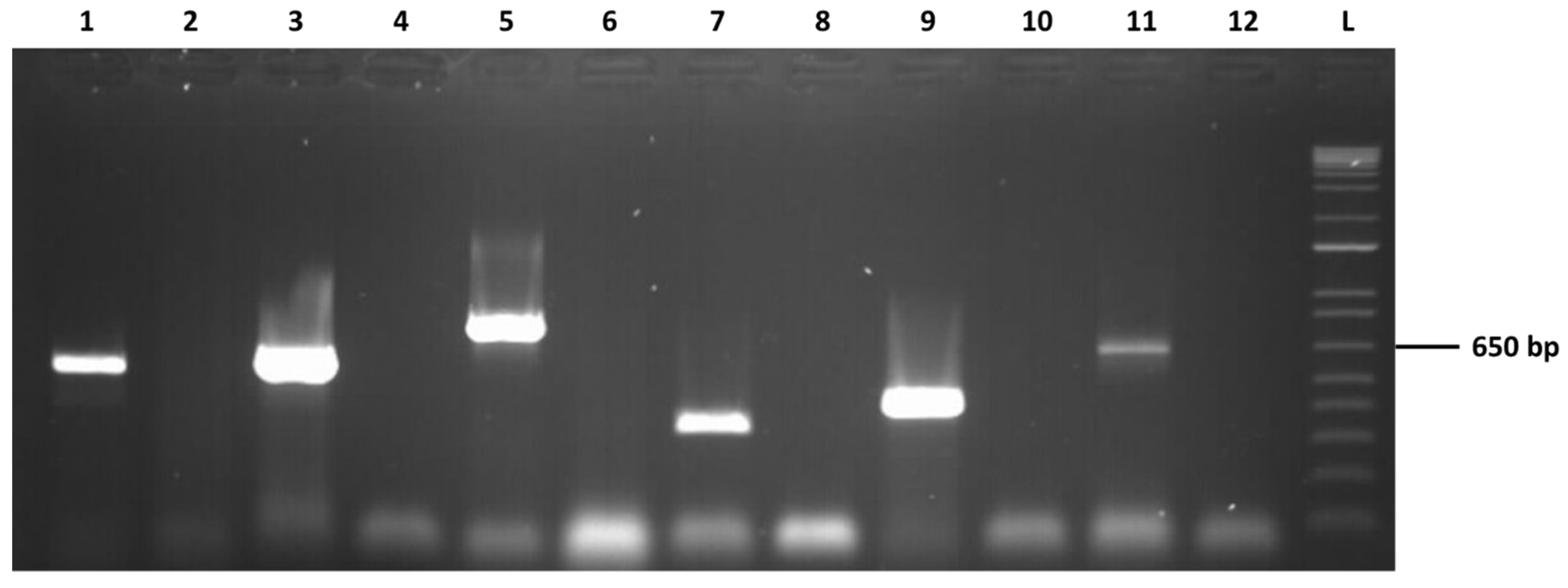
2.3. Confirmation of the Presence of the Detected Viruses
2.4. Phylogenetic Analyses
3. Results and Discussion
3.1. Mycoviruses Identification from N. parvum COLB
3.2. Molecular Features and Phylogenetic Relationships of the Identified Mycoviruses
3.2.1. Putative Novel Virus Belonging to the Family Endornaviridae
3.2.2. Putative Novel Viruses Belonging to the Families Narnaviridae and Mitoviridae
3.2.3. Putative Novel Viruses Belonging to the Family Totiviridae
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fontaine, F.; Gramaje, D.; Armengol, J.; Smardt, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.F. Grapevine Trunck Diseases: A Review; OIV Publications: Paris, France, 2016; 24p. [Google Scholar]
- Bertsch, C.; Ramirez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clement, C.; Fontaine, F. Grapevine trunck diseases: Complex and still poorly understood. Plant Pathol. 2013, 62, 243–265. [Google Scholar] [CrossRef] [Green Version]
- Claverie, M.; Notaro, M.; Fontaine, F.; Wery, J. Current knowledge on Grapevine Trunk Diseases with complex etiology: A systemic approach. Phytopathol. Mediter 2020, 59, 29–53. [Google Scholar]
- Uber-Torres, J.R. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr 2011, 50, 5–45. [Google Scholar]
- Pitt, W.M.; Huang, R.; Steel, C.C.; Savocchia, S. Pathogenicity and epidemiology of Botryosphaeriaceae species isolated from grapevines in Australia. Australas. Plant Pathol. 2013, 42, 573–582. [Google Scholar] [CrossRef]
- Comont, C.; Mayet, V.; Corio-Costet, M.F. First report of Lasiodiplodia viticola, Spencermartinsia viticola and Diplodia intermedia associated with Vitis vinifera decline in French Vineyards. Plant Dis. 2016, 100, 2328. [Google Scholar] [CrossRef]
- Urbez-Torres, J.R.; Adams, P.; Kamas, J.; Gubler, W.D. Identification, incidence, and pathogenicity of fungal species associated with grapevine dieback in Texas. Am. J. Enol. Viticult. 2009, 60, 497–507. [Google Scholar]
- Larignon, P.; Fulchic, R.; Cere, L.; Dubos, B. Observation of black dead arm in French vineyards. Phytopathol. Mediterr. 2001, 40, S336–S342. [Google Scholar]
- Bellée, A.; Comont, G.; Nivault, A.; Abou-Mansour, E.; Coppin, C.; Dufour, M.C.; Corio-Costet, M.F. Life traits of four Botryosphaeriaceae species and molecular responses of different grapevine cultivars or hybrids. Plant Pathol. 2017, 66, 763–776. [Google Scholar] [CrossRef] [Green Version]
- Amponsah, N.T.; Jones, E.E.; Ridgway, H.J.; Jaspers, M.V. Identification, potential inoculum sources and pathogenicity of botryospaeriaceous species associated with grapevine dieback disease in New Zealand. Eur. J. Plant Pathol. 2011, 131, 467–482. [Google Scholar] [CrossRef]
- Reveglia, P.; Savocchia, S.; Billones-Baaijens, R.; Masi, M.; Cimmino, A.; Evidente, A. Phytotoxic metabolites by nine species of Botryosphaeriaceae involved in grapevine dieback in Australia and identification of those produced by Diplodia mutila, Diplodia seriata, Neofusicoccum australe and Neofusicoccum Luteum. Nat. Prod. Res. 2019, 33, 2223–2229. [Google Scholar] [CrossRef] [PubMed]
- Stempien, E.; Goddard, M.L.; Wilhelm, K.; Tarnus, C.; Bertsch, C.; Chong, J. Grapevine Botryosphaeria dieback fungi have specific aggressiveness factor repertory involved in wood decay and stilbene metabolization. PLoS ONE 2017, 12, e0188766. [Google Scholar] [CrossRef]
- Massonnet, M.; Morales-Cruz, A.; Figueroa-Balderas, R.; Lawrence, D.P.; Baumgartner, K.; Cantu, D. Condition-dependent co-regulation of genomic clusters of virulence factors in the grapevine trunk pathogen Neofusicoccum parvum. Mol. Plant Pathol. 2018, 19, 21–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskarathevan, J.; Jaspers, M.V.; Jones, E.E.; Cruickshank, R.H.; Ridgway, H.J. Genetic and pathogenic diversity of Neofusicoccum parvum in New Zealand vineyards. Fungal Biol. 2012, 116, 276–288. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Caston, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, M.; Zhu, H.; Zhong, J.; Liao, X.; Zhou, Q. Molecular characterization of a novel mitovirus from the plant-pathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2020. [Google Scholar] [CrossRef]
- Zhai, L.; Xiang, J.; Zhang, M.; Fu, M.; Yang, Z.; Wang, G. Characterization of a novel double-stranded RNA mycovirus conferring hypovirulence to the phytopathogenic fungus Botryosphaeria dothidea. Virology 2016, 493, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Luo, H.; Hu, W.; Yang, Y.; Hong, N.; Wang, G. De novo transcriptomic assembly and mRNA expression patterns of Botryosphaeria dothidea infection with mycoviruses chrysovirus 1 (BdCV1) and partitivirus 1 (BdPV1). Virol. J. 2018, 15, 126–142. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Luo, H.; Yang, Y.; Wang, Q.; Hong, N.; Wang, G.; Wang, A.; Wang, L. Comprehensive analysis of full genome sequence and Bd-milRNA/target mRNAs to discover the mechanism of hypovirulence in Botryosphaeria dothidea strains on pear infection with BdCV1 and BdPV1. Ima Fungus 2019, 10, 3–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marais, A.; Nivault, A.; Faure, C.; Theil, S.; Comont, G.; Candresse, T.; Corio-Costet, M.F. Determination of the complete genomic sequence of neofusicoccum luteum mitovirus 1 (NlMV1), a novel mitovirus associated with a pathogenic Botryosphaeriaceae. Arch. Virol. 2017, 162, 2477–2480. [Google Scholar] [CrossRef] [Green Version]
- Marais, A.; Nivault, A.; Faure, C.; Comont, G.; Theil, S.; Candresse, T.; Corio-Costet, M.F. Molecular charaterizationof a novel fusarivirus infecting the plant-pathogenic fungus Neofusicoccum luteum. Arch. Virol. 2018, 163, 559–562. [Google Scholar] [CrossRef]
- Nerva, L.; Turina, M.; Zanzotto, A.; Gardiman, M.; Gaiotti, F.; Gambino, G.; Chitarra, W. Isolation, molecular characterization and virome analysis of culturable wood fungal endophytes in esca symptomatic and asymptomatic grapevines plants. Environ. Microbiol. 2019, 21, 2886–2904. [Google Scholar] [CrossRef]
- Comont, G.; Corio-Costet, M.F.; Larignon, P.; Delmotte, F. AFLP markers reveal two genetic groups in the French population of the grapevine fungal pathogen Phaeomoniella chlamydospora. Eur. J. Plant Pathol. 2010, 127, 451–464. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marais, A.; Faure, C.; Couture, C.; Bergey, B.; Gentit, P.; Candresse, T. Characterization by deep sequencing of divergent Plum bark necrosis stem pitting-associated virus isolates and development of a broad-spectrum PBNSPaV detection assay. Phytopathology 2014, 104, 660–666. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, F.; Xie, J.; Cheng, S.; You, M.P.; Barbetti, M.J.; Jia, J.; Wang, Q.; Cheng, J.; Fu, Y.; Chen, T.; et al. Virome Characterization of a collection of Sclerotinia sclerotiorum from Australia. Front. Microbiol. 2018, 8, 2540–2557. [Google Scholar] [CrossRef] [Green Version]
- Preisig, O.; Wingfield, B.D.; Wingfield, M.J. Coinfection of a fungal pathogen by two distinct double-stranded RNA viruses. Virology 1998, 252, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osaki, H.; Sasaki, A.; Nomiyama, K.; Tomioka, K. Multiple virus infection in a single strain of Fusarium poae shown by deep sequencing. Virus Genes 2016, 52, 835–847. [Google Scholar] [CrossRef]
- Yang, M.; Zhai, L.; Xiao, F.; Guo, Y.; Fu, M.; Hong, N.; Wang, G. Characterization of a novel victorivirus isolated from the phytopathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2019, 164, 1609–1617. [Google Scholar] [CrossRef]
- Valverde, R.A.; Khalifa, M.E.; Okada, R.; Fukuhara, T.; Sabanadzovic, S.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Endornaviridae. J. Gen Virol. 2019, 100, 1204–1205. [Google Scholar] [CrossRef]
- Hillman, B.I.; Cai, G. The family Narnaviridae: Simplest of RNA viruses. Adv. Virus Res. 2013, 86, 149–176. [Google Scholar]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Dover, S.L.; Coel, T.E.; Brasier, C.M.; Buck, K.W. Multiple mitochondrial viruses in an isolate of the Dutch elm disease fungus Ophiostoma novo-ulmi. Virology 1999, 258, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Zhou, J.; Zhou, X.; Shuai, S.; Zhou, R.; An, H.; Fang, S.; Zhang, S.; Deng, Q. A novel narnavirus from the plant-pathogenic fungus Magnaporthe Oryzae. Arch. Virol. 2020, 165, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Hillman, B.I.; Esteban, R. Family Narnaviridae. In Virus Taxonomy-Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2012; pp. 1055–1060. [Google Scholar]
- Ghabrial, S.A.; Nibert, M.L. Victorivirus, a new genus of fungal viruses in the family Totiviridae. Arch. Virol. 2009, 154, 373–379. [Google Scholar] [CrossRef]
- Wickner, R.B.; Ghabrial, S.A.; Nibert, M.L.; Patterson, J.L.; Wang, C.C. Family Totiviridae. In Virus Taxonomy-Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2012; pp. 639–650. [Google Scholar]
- Zoll, J.; Verweij, P.E.; Melchers, W.J. G Discovery and characterization of novel Aspergillus fumigatus mycoviruses. PLoS ONE 2018, 13, e0200511. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Z.; Zhu, H.J.; Gao, B.D.; Zhou, Q.; Zhong, J. Diverse, novel mycovirses from the virome of a hypovirulent Sclerotium rolfsii strain. Front. Plant Sci. 2018, 9, 1738. [Google Scholar] [CrossRef] [Green Version]
- Okada, R.; Ichinose, S.; Takeshita, K.; Urayama, S.; Fukuhara, T.; Komatsu, K.; Arie, T.; Ishihara, A.; Egusa, M.; Kodama, M.; et al. Molecular characterization of a novel mycovirus in Alternaria alternata manifesting two-sided effects: Down-regulation of host growth and upregulation of host plant pathogenicity. Virology 2018, 519, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Havens, W.; Lin, Y.H.; Suzuki, N.; Ghabrial, S.A. The victorivirus Helminthosporium victoriae virus 190S is the primary cause of disease/hypovirulence in its natural host and a heterologous host. Virus Res. 2016, 213, 238–245. [Google Scholar] [CrossRef]
- Liu, H.; Liu, R.; Li, C.X.; Wang, H.; Zhy, H.J.; Gao, B.D.; Zhou, Q.; Zhong, J. A victorivirus and two novel mitoviruses co-infected the plant pathogen Nigrospora oryzae. Viruses 2019, 11, 83. [Google Scholar] [CrossRef] [Green Version]
- Xu, A.; Wu, S.; Liu, L.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2015, 197, 127–136. [Google Scholar] [CrossRef]
- Fukuhara, T. Endornaviruses: Persistent dsRNA viruses with symbiotic properties in diverse eukaryotes. Virus Genes 2019, 55, 165–173. [Google Scholar] [CrossRef]
- Zheng, L.; Shu, C.; Zhang, M.; Yang, M.; Zhou, E. Molecuar characterization of a novel endornavirus conferring hypovirulence in rice sheath blight fungus Rhizoctonia solani AG-1 IA strain GD-2. Viruses 2019, 11, 178. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Wu, M.; Zhang, J.; Chen, W.; Li, G.; Yang, L. Sclerotinia minor endornavirus 1, a novel pathogenicity debilitation-associated mycovirus with a wide spectrum of horizontal transmissibility. Viruses 2018, 10, 589. [Google Scholar] [CrossRef] [Green Version]
- Owashi, Y.; Aihara, M.; Moriyama, H.; Arie, T.; Teraoka, T.; Komatsu, K. Population structure of double stranded RNA mycoviruses that infect the rice blast fungus Magnaporthe oryzae in Japan. Front. Microbiol. 2020, 11, 593784. [Google Scholar] [CrossRef]
- Thapa, V.; Roossinck, M.J. Determinants of coinfection in the mycoviruses. Front Cell Infect Microbiol. 2019, 9, 169. [Google Scholar] [CrossRef] [Green Version]
- Hillman, B.I.; Annisa, A.; Suzuki, N. Viruses of plant-interacting fungi. Adv. Virus Res. 2018, 100, 99–116. [Google Scholar]
- Sasaki, A.; Nakamura, H.; Suzuki, N.; Kanematsu, S. Characterization of a new megabirnavirus that confers hypovirulence with the aid of a co-infecting partitivirus to the host fungus, Rosellinia necatrix. Virus Res. 2016, 219, 73–82. [Google Scholar] [CrossRef]
- Cao, C.; Li, H.; Jones, M.G.K.; Wylie, S.J. Challenges to elucidating how endornaviruses influence fungal hosts: Creating mycovirus-free isogenic fungal lines and testing them. J. Virol. Methods 2019, 272, 113745. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Targeted Virus (ORF) 1 | Sequence 5′-3′ | Size (nt) | Ta (°C) 2 | Purpose 3 |
|---|---|---|---|---|---|
| COLB-Endorna-Race3 | NpEV1 (3′end) | GGACGGTATGAGACTGACTGGACAGGCG | 674 | 72 | RACE |
| COLB-Alpha-Endorna-FD | NpEV1 (ORF1) | TCCAATGTGGTCCAATGCATC | 771 | 52 | Detection |
| COLB-Alpha-Endorna-RD | GTGTCAACTATTTTTGAAGTG | ||||
| COLB-Narna-Race5 | NpNV3 (5′end) | TGATTGAAGATGTACAGGGTAAAGA | 259 | 64 | RACE |
| COLB-Narna-Race3 | NpNV3 (3′end) | TCATCCTACACATGTGCCATTTAGTGG | 168 | 72 | RACE |
| COLB-Narna-FD | NpNV3 (RdRp) | AAGGGACCTATTGGTTCCGCA | 417 | 58 | Detection |
| COLB-Narna-RD | GGATGTCGATGGACGAAATGT | ||||
| COLB-Mito2-Race5 | NpMV2 (5′end) | TTCCGTAGAACCGGCATGACACCCACT | 141 | 68 | RACE |
| COLB-Mito2-Race3 | NpMV2 (3′end) | TTCGAGCTCTGACTTCGGTCAGTAGGG | 361 | 68 | RACE |
| COLB-Mito2-FD | NpMV2 (RdRp) | AGTCCAACCGTTCCTAAAGAC | 568 | 56 | Detection |
| COLB-Mito2-RD | CGGAATAAGTATAATCAGATCC | ||||
| COLB-Mito3-Race5 | NpMV3 (5′end) | CCACATCCTGTTGCTCAGACTGCTACAC | 444 | 68 | RACE |
| COLB-Mito3-Race3 | NpMV3 (3′end) | CCACAACCTTACTTCCAATCTAACCT | 282 | 58 | RACE |
| COLB-Mito3-FD | NpMV3 (RdRp) | TATTCTTTCACTCCTTGGAGT | 538 | 52 | Detection |
| COLB-Mito3-RD | TCACATAATGAGACCATAACT | ||||
| COLB-Victori1-Race5 | NpVV1 (5′end) | GCTGGCGGCGGCAAGGGTGGA | 128 | 72 | RACE |
| COLB-Victori1-Race3 | NpVV1 (3′end) | GTGACCACGCTACTAATCGAAGCTGGAA | 253 | RACE | |
| COLB-Victori1-FD | NpVV1 (RdRp) | TCTCCGTCGTTGAGCGGTACCCTG | 649 | 67 | Detection |
| COLB-Victori1-RD | GGTGCTCCGAAGTCATGGGCATT | ||||
| COLB-Victori2-Race5 | NpVV2 (5′end) | AGGGTAGGCTTGGTGGGAAACGGGAG | 326 | 65 | RACE |
| COLB-Victori2-F5 | NpVV2 (RdRp) | AAGCTGAAGCGACATACCGCACG | 338 | 62 | Detection |
| COLB-Victori2-R5 | CGAGGCTGAGACGTAAGTGTGTC |
| Contig Name | Contig Length (nt) | Provisional Virus Name | BlastX Best Hit 1 (Accession Number) | Amino Acid Identity | Mapped Reads | % Total Teads | Family | Reference |
|---|---|---|---|---|---|---|---|---|
| Ct 1 | 2885 | NpMV2 | RdRp Mitovirus sp. (QDH8995) | 40.7% | 29,121,096 | 25.3% | Mitoviridae | unpublished |
| Ct 2 | 2629 | NpMV3 | RdRp Botryosphaeria dothidea mitovirus 1(QMU24933) | 39.8% | 22,033,854 | 19.1% | Mitoviridae | unpublished |
| Ct 5 | 13,816 | NpEV1 | Polyprotein Sclerotinia sclerotiorum endornavirus 3 (AWY10956) | 55.2% | 1,193,076 | 1.0% | Endornaviridae | [27] |
| Ct 9 | 5195 | NpVV2 | CP Sphaeropsis sapinea RNA virus 2 (NP047559) | 71.2% | 1,143,697 | 1.0% | Totiviridae | [28] |
| Ct 14 | 2064 | NpNV3 | RdRp Fusarium poae narnavirus 2 (YP009272903) | 79.4% | 598,911 | 0.5% | Narnaviridae | [29] |
| Ct 16 | 5194 | NpVV1 | CP Botryosphaeria dothidea victorivirus 2 (QBA82442) | 72.7% | 552,571 | 0.5% | Totiviridae | [30] |
| Virus Name 1 | NpNV3 | FpNV2 | NpNV2 | AfNV1 | ScNV-20S | NpNV1 | ScNV-23S |
|---|---|---|---|---|---|---|---|
| FpNV2 | 77.6 2 | ||||||
| NpNV2 | 67.8 | 67.2 | |||||
| AfNV1 | 55.4 | 53.5 | 54.1 | ||||
| ScNV-20S | 7.9 | 8.2 | 6.7 | 8 | |||
| NpNV1 | 7.4 | 7.6 | 7.7 | 8.1 | 8.6 | ||
| ScNV-23S | 7.1 | 7.4 | 7.9 | 6.6 | 16.8 | 7.9 | |
| FpNV1 | 5.9 | 6.1 | 5.5 | 6 | 11.9 | 9.2 | 10.7 |
| Virus Name 1 | NpVV1 | NpVV2 | SsRV12 | BdVV2 | SsRV2 | FaVV1 | RnVV1 |
|---|---|---|---|---|---|---|---|
| NpVV1 | 34.6% | 66.3% | 73.9% | 34.4% | 48.8% | 47.5% | |
| NpVV2 | 35.2% | 34.6% | 35.2% | 67.2% | 37.9% | 36.2% | |
| SsRV1 | 62.9% | 34.4% | 67.9% | 36% | 49.7% | 49.8% | |
| BdVV2 | 60.5% | 34.2% | 60.4% | 35.8% | 48.5% | 48.3% | |
| SsRV2 | 33.9% | 66% | 35.4% | 33.7% | 37.8% | 37.4% | |
| FaVV1 | 45% | 36% | 45.4% | 43.5% | 35.9% | 77.6% | |
| RnVV1 | 45% | 34.9% | 44.5% | 43.3% | 35.7% | 75.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marais, A.; Faure, C.; Comont, G.; Candresse, T.; Stempien, E.; Corio-Costet, M.-F. Characterization of the Mycovirome of the Phytopathogenic Fungus, Neofusicoccum parvum. Viruses 2021, 13, 375. https://doi.org/10.3390/v13030375
Marais A, Faure C, Comont G, Candresse T, Stempien E, Corio-Costet M-F. Characterization of the Mycovirome of the Phytopathogenic Fungus, Neofusicoccum parvum. Viruses. 2021; 13(3):375. https://doi.org/10.3390/v13030375
Chicago/Turabian StyleMarais, Armelle, Chantal Faure, Gwenaëlle Comont, Thierry Candresse, Elodie Stempien, and Marie-France Corio-Costet. 2021. "Characterization of the Mycovirome of the Phytopathogenic Fungus, Neofusicoccum parvum" Viruses 13, no. 3: 375. https://doi.org/10.3390/v13030375
APA StyleMarais, A., Faure, C., Comont, G., Candresse, T., Stempien, E., & Corio-Costet, M. -F. (2021). Characterization of the Mycovirome of the Phytopathogenic Fungus, Neofusicoccum parvum. Viruses, 13(3), 375. https://doi.org/10.3390/v13030375