
From Submerged Cultures to 3D Cell Culture Models: Evolution of Nasal Epithelial Cells in Asthma Research and Virus Infection


 , and
, and 
Abstract
:1. Introduction
2. A General Overview on Cell Culture Techniques in Respiratory Medicine
3. Bronchial Epithelial versus Nasal Epithelial Cell Culture: Benefits and Drawbacks
4. The Application of Nasal Epithelial Cell Culture Models in Asthma and Virus Research
5. Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Braman, S.S. The global burden of asthma. Chest 2006, 130, 4S–12S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambrecht, B.N.; Hammad, H. The immunology of asthma. Nat. Immunol 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Asher, I.; Pearce, N. Global burden of asthma among children. Int. J. Tuberc Lung Dis. 2014, 18, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. The airway epithelium in asthma. Nat. Med. 2012, 18, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, S.E. Asthma phenotypes: The evolution from clinical to molecular approaches. Nat. Med. 2012, 18, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Clarisse, B.; Nikasinovic, L.; Poinsard, R.; Just, J.; Momas, I. The Paris prospective birth cohort study: Which design and who participates? Eur. J. Epidemiol. 2007, 22, 203–210. [Google Scholar] [CrossRef]
- Fuchs, O.; Genuneit, J.; Latzin, P.; Buchele, G.; Horak, E.; Loss, G.; Sozanska, B.; Weber, J.; Boznanski, A.; Heederik, D.; et al. Farming environments and childhood atopy, wheeze, lung function, and exhaled nitric oxide. J. Allergy Clin. Immunol. 2012, 130, 382–388.e386. [Google Scholar] [CrossRef]
- Fuertes, E.; Standl, M.; Cyrys, J.; Berdel, D.; von Berg, A.; Bauer, C.P.; Kramer, U.; Sugiri, D.; Lehmann, I.; Koletzko, S.; et al. A longitudinal analysis of associations between traffic-related air pollution with asthma, allergies and sensitization in the GINIplus and LISAplus birth cohorts. Peer J. 2013, 1, e193. [Google Scholar] [CrossRef] [PubMed]
- Illi, S.; von Mutius, E.; Lau, S.; Niggemann, B.; Gruber, C.; Wahn, U.; Multicentre Allergy Study, g. Perennial allergen sensitisation early in life and chronic asthma in children: A birth cohort study. Lancet 2006, 368, 763–770. [Google Scholar] [CrossRef]
- Lau, S.; Nickel, R.; Niggemann, B.; Gruber, C.; Sommerfeld, C.; Illi, S.; Kulig, M.; Forster, J.; Wahn, U.; Groeger, M.; et al. The development of childhood asthma: Lessons from the German Multicentre Allergy Study (MAS). Paediatr. Respir. Rev. 2002, 3, 265–272. [Google Scholar] [CrossRef]
- Kusel, M.M.; de Klerk, N.H.; Kebadze, T.; Vohma, V.; Holt, P.G.; Johnston, S.L.; Sly, P.D. Early-life respiratory viral infections, atopic sensitization, and risk of subsequent development of persistent asthma. J. Allergy Clin. Immunol. 2007, 119, 1105–1110. [Google Scholar] [CrossRef]
- Loss, G.J.; Depner, M.; Hose, A.J.; Genuneit, J.; Karvonen, A.M.; Hyvarinen, A.; Roduit, C.; Kabesch, M.; Lauener, R.; Pfefferle, P.I.; et al. The Early Development of Wheeze. Environmental Determinants and Genetic Susceptibility at 17q21. Am. J. Respir. Crit. Care Med. 2016, 193, 889–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, J.; Hilliard, T.N.; Sherriff, A.; Stalker, D.; Al Shammari, N.; Thomas, H.M. Hospitalization for RSV bronchiolitis before 12 months of age and subsequent asthma, atopy and wheeze: A longitudinal birth cohort study. Pediatr. Allergy Immunol. 2005, 16, 386–392. [Google Scholar] [CrossRef]
- Sigurs, N.; Bjarnason, R.; Sigurbergsson, F.; Kjellman, B. Respiratory syncytial virus bronchiolitis in infancy is an important risk factor for asthma and allergy at age 7. Am. J. Respir. Crit. Care Med. 2000, 161, 1501–1507. [Google Scholar] [CrossRef]
- Message, S.D.; Laza-Stanca, V.; Mallia, P.; Parker, H.L.; Zhu, J.; Kebadze, T.; Contoli, M.; Sanderson, G.; Kon, O.M.; Papi, A.; et al. Rhinovirus-induced lower respiratory illness is increased in asthma and related to virus load and Th1/2 cytokine and IL-10 production. Proc. Natl. Acad. Sci. USA 2008, 105, 13562–13567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olenec, J.P.; Kim, W.K.; Lee, W.M.; Vang, F.; Pappas, T.E.; Salazar, L.E.; Evans, M.D.; Bork, J.; Roberg, K.; Lemanske, R.F., Jr.; et al. Weekly monitoring of children with asthma for infections and illness during common cold seasons. J. Allergy Clin. Immunol. 2010, 125, 1001–1006.e1001. [Google Scholar] [CrossRef]
- Christensen, A.; Kesti, O.; Elenius, V.; Eskola, A.L.; Dollner, H.; Altunbulakli, C.; Akdis, C.A.; Soderlund-Venermo, M.; Jartti, T. Human bocaviruses and paediatric infections. Lancet Child. Adolesc. Health 2019, 3, 418–426. [Google Scholar] [CrossRef]
- Coverstone, A.M.; Wang, L.; Sumino, K. Beyond Respiratory Syncytial Virus and Rhinovirus in the Pathogenesis and Exacerbation of Asthma: The Role of Metapneumovirus, Bocavirus and Influenza Virus. Immunol. Allergy Clin. N. Am. 2019, 39, 391–401. [Google Scholar] [CrossRef]
- Docherty, A.B.; Harrison, E.M.; Green, C.A.; Hardwick, H.E.; Pius, R.; Norman, L.; Holden, K.A.; Read, J.M.; Dondelinger, F.; Carson, G.; et al. Features of 20 133 UK patients in hospital with covid-19 using the ISARIC WHO Clinical Characterisation Protocol: Prospective observational cohort study. BMJ 2020, 369, m1985. [Google Scholar] [CrossRef]
- Papadopoulos, N.G.; Christodoulou, I.; Rohde, G.; Agache, I.; Almqvist, C.; Bruno, A.; Bonini, S.; Bont, L.; Bossios, A.; Bousquet, J.; et al. Viruses and bacteria in acute asthma exacerbations--a GA(2) LEN-DARE systematic review. Allergy 2011, 66, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Soto-Quiros, M.; Avila, L.; Platts-Mills, T.A.; Hunt, J.F.; Erdman, D.D.; Carper, H.; Murphy, D.D.; Odio, S.; James, H.R.; Patrie, J.T.; et al. High titers of IgE antibody to dust mite allergen and risk for wheezing among asthmatic children infected with rhinovirus. J. Allergy Clin. Immunol. 2012, 129, 1499–1505.e1495. [Google Scholar] [CrossRef]
- Aydin, M.; Naumova, E.A.; Paulsen, F.; Zhang, W.; Gopon, F.; Theis, C.; Lutz, S.; Ehrke-Schulz, E.; Arnold, W.H.; Wirth, S.; et al. House Dust Mite Exposure Causes Increased Susceptibility of Nasal Epithelial Cells to Adenovirus Infection. Viruses 2020, 12, 1151. [Google Scholar] [CrossRef] [PubMed]
- Bochkov, Y.A.; Palmenberg, A.C.; Lee, W.M.; Rathe, J.A.; Amineva, S.P.; Sun, X.; Pasic, T.R.; Jarjour, N.N.; Liggett, S.B.; Gern, J.E. Molecular modeling, organ culture and reverse genetics for a newly identified human rhinovirus C. Nat. Med. 2011, 17, 627–632. [Google Scholar] [CrossRef]
- Zeng, H.; Goldsmith, C.S.; Kumar, A.; Belser, J.A.; Sun, X.; Pappas, C.; Brock, N.; Bai, Y.; Levine, M.; Tumpey, T.M.; et al. Tropism and Infectivity of a Seasonal A(H1N1) and a Highly Pathogenic Avian A(H5N1) Influenza Virus in Primary Differentiated Ferret Nasal Epithelial Cell Cultures. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergougnoux, A.; Claustres, M.; De Sario, A. Nasal epithelial cells: A tool to study DNA methylation in airway diseases. Epigenomics 2015, 7, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Kohno, T.; Kakuki, T.; Takano, K.I.; Ogasawara, N.; Miyata, R.; Kikuchi, S.; Konno, T.; Ohkuni, T.; Yajima, R.; et al. The role of transcriptional factor p63 in regulation of epithelial barrier and ciliogenesis of human nasal epithelial cells. Sci. Rep. 2017, 7, 10935. [Google Scholar] [CrossRef] [PubMed]
- McDougall, C.M.; Blaylock, M.G.; Douglas, J.G.; Brooker, R.J.; Helms, P.J.; Walsh, G.M. Nasal epithelial cells as surrogates for bronchial epithelial cells in airway inflammation studies. Am. J. Respir. Cell Mol. Biol. 2008, 39, 560–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sungnak, W.; Huang, N.; Becavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Yao, T.; Asayama, Y. Animal-cell culture media: History, characteristics, and current issues. Reprod. Med. Biol. 2017, 16, 99–117. [Google Scholar] [CrossRef]
- Jones, H.W., Jr. Record of the first physician to see Henrietta Lacks at the Johns Hopkins Hospital: History of the beginning of the HeLa cell line. Am. J. Obstet. Gynecol. 1997, 176, S227–S228. [Google Scholar] [CrossRef]
- Lucey, B.P.; Nelson-Rees, W.A.; Hutchins, G.M. Henrietta Lacks, HeLa cells, and cell culture contamination. Arch. Pathol. Lab. Med. 2009, 133, 1463–1467. [Google Scholar] [CrossRef] [PubMed]
- Freishtat, R.J.; Nino, G.; Tsegaye, Y.; Alcala, S.E.; Benton, A.S.; Watson, A.M.; Reeves, E.K.; Haider, S.K.; Damsker, J.M. Pharmacologically-induced mitotic synchrony in airway epithelial cells as a mechanism of action of anti-inflammatory drugs. Respir. Res. 2015, 16, 132. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.C.; Leyko, B.; Frieri, M. Effects of omalizumab and budesonide on markers of inflammation in human bronchial epithelial cells. Ann. Allergy Asthma Immunol. 2005, 95, 443–451. [Google Scholar] [CrossRef]
- Cao, X.; Coyle, J.P.; Xiong, R.; Wang, Y.; Heflich, R.H.; Ren, B.; Gwinn, W.M.; Hayden, P.; Rojanasakul, L. Invited review: Human air-liquid-interface organotypic airway tissue models derived from primary tracheobronchial epithelial cells-overview and perspectives. Vitro Cell. Dev. Biol. Anim. 2020. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.; Palmberg, L. Air-Liquid Interface: Relevant In Vitro Models for Investigating Air Pollutant-Induced Pulmonary Toxicity. Toxicol. Sci. 2018, 164, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorissen, M.; Van der Schueren, B.; Van den Berghe, H.; Cassiman, J.J. The preservation and regeneration of cilia on human nasal epithelial cells cultured in vitro. Arch. Otorhinolaryngol. 1989, 246, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Fennema, E.; Rivron, N.; Rouwkema, J.; van Blitterswijk, C.; de Boer, J. Spheroid culture as a tool for creating 3D complex tissues. Trends Biotechnol. 2013, 31, 108–115. [Google Scholar] [CrossRef]
- Barrett, L.A.; McDowell, E.M.; Frank, A.L.; Harris, C.C.; Trump, B.F. Long-term organ culture of human bronchial epithelium. Cancer Res. 1976, 36, 1003–1010. [Google Scholar]
- Lechner, J.F.; Haugen, A.; McClendon, I.A.; Pettis, E.W. Clonal growth of normal adult human bronchial epithelial cells in a serum-free medium. Vitro 1982, 18, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Stoner, G.D.; Katoh, Y.; Foidart, J.M.; Myers, G.A.; Harris, C.C. Identification and culture of human bronchial epithelial cells. Methods Cell Biol. 1980, 21A, 15–35. [Google Scholar] [CrossRef]
- Fulcher, M.L.; Randell, S.H. Human nasal and tracheo-bronchial respiratory epithelial cell culture. Methods Mol. Biol. 2013, 945, 109–121. [Google Scholar] [CrossRef]
- Pezzulo, A.A.; Starner, T.D.; Scheetz, T.E.; Traver, G.L.; Tilley, A.E.; Harvey, B.G.; Crystal, R.G.; McCray, P.B., Jr.; Zabner, J. The air-liquid interface and use of primary cell cultures are important to recapitulate the transcriptional profile of in vivo airway epithelia. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 300, L25–L31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prytherch, Z.; Job, C.; Marshall, H.; Oreffo, V.; Foster, M.; BeruBe, K. Tissue-Specific stem cell differentiation in an in vitro airway model. Macromol. Biosci. 2011, 11, 1467–1477. [Google Scholar] [CrossRef] [PubMed]
- Gruenert, D.C.; Finkbeiner, W.E.; Widdicombe, J.H. Culture and transformation of human airway epithelial cells. Am. J. Physiol. 1995, 268, L347–L360. [Google Scholar] [CrossRef]
- Leung, C.; Wadsworth, S.J.; Yang, S.J.; Dorscheid, D.R. Structural and functional variations in human bronchial epithelial cells cultured in air-liquid interface using different growth media. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 318, L1063–L1073. [Google Scholar] [CrossRef] [PubMed]
- Gras, D.; Petit, A.; Charriot, J.; Knabe, L.; Alagha, K.; Gamez, A.S.; Garulli, C.; Bourdin, A.; Chanez, P.; Molinari, N.; et al. Epithelial ciliated beating cells essential for ex vivo ALI culture growth. BMC Pulm. Med. 2017, 17, 80. [Google Scholar] [CrossRef] [Green Version]
- Gras, D.; Bourdin, A.; Vachier, I.; de Senneville, L.; Bonnans, C.; Chanez, P. An ex vivo model of severe asthma using reconstituted human bronchial epithelium. J. Allergy Clin. Immunol. 2012, 129, 1259–1266.e1251. [Google Scholar] [CrossRef]
- Whitcutt, M.J.; Adler, K.B.; Wu, R. A biphasic chamber system for maintaining polarity of differentiation of cultured respiratory tract epithelial cells. Vitro Cell Dev. Biol. 1988, 24, 420–428. [Google Scholar] [CrossRef]
- Ravi, M.; Paramesh, V.; Kaviya, S.R.; Anuradha, E.; Solomon, F.D. 3D cell culture systems: Advantages and applications. J. Cell Physiol 2015, 230, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; David, B.T.; Trawczynski, M.; Fessler, R.G. Advances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and Applications. Stem. Cell Rev. Rep. 2020, 16, 3–32. [Google Scholar] [CrossRef] [Green Version]
- Bolker, J.A. Animal Models in Translational Research: Rosetta Stone or Stumbling Block? Bioessays 2017, 39. [Google Scholar] [CrossRef]
- Robinson, N.B.; Krieger, K.; Khan, F.M.; Huffman, W.; Chang, M.; Naik, A.; Yongle, R.; Hameed, I.; Krieger, K.; Girardi, L.N.; et al. The current state of animal models in research: A review. Int. J. Surg. 2019, 72, 9–13. [Google Scholar] [CrossRef]
- Kips, J.C.; Anderson, G.P.; Fredberg, J.J.; Herz, U.; Inman, M.D.; Jordana, M.; Kemeny, D.M.; Lotvall, J.; Pauwels, R.A.; Plopper, C.G.; et al. Murine models of asthma. Eur. Respir. J. 2003, 22, 374–382. [Google Scholar] [CrossRef]
- Zscheppang, K.; Berg, J.; Hedtrich, S.; Verheyen, L.; Wagner, D.E.; Suttorp, N.; Hippenstiel, S.; Hocke, A.C. Human Pulmonary 3D Models For Translational Research. Biotechnol. J. 2018, 13. [Google Scholar] [CrossRef]
- Berube, K.; Prytherch, Z.; Job, C.; Hughes, T. Human primary bronchial lung cell constructs: The new respiratory models. Toxicology 2010, 278, 311–318. [Google Scholar] [CrossRef]
- Ramanan, V.; Scull, M.A.; Sheahan, T.P.; Rice, C.M.; Bhatia, S.N. New Methods in Tissue Engineering: Improved Models for Viral Infection. Annu Rev. Virol 2014, 1, 475–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.Z.; Chang, H.Y. Recent advances in three-dimensional multicellular spheroid culture for biomedical research. Biotechnol. J. 2008, 3, 1172–1184. [Google Scholar] [CrossRef]
- Moroni, L.; de Wijn, J.R.; van Blitterswijk, C.A. Integrating novel technologies to fabricate smart scaffolds. J. Biomater. Sci. Polym. Ed. 2008, 19, 543–572. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, B.V.; Khurshid, S.S.; Fisher, O.Z.; Khademhosseini, A.; Peppas, N.A. Hydrogels in regenerative medicine. Adv. Mater. 2009, 21, 3307–3329. [Google Scholar] [CrossRef] [Green Version]
- Bridges, M.A.; Walker, D.C.; Harris, R.A.; Wilson, B.R.; Davidson, A.G. Cultured human nasal epithelial multicellular spheroids: Polar cyst-like model tissues. Biochem. Cell Biol. 1991, 69, 102–108. [Google Scholar] [CrossRef]
- Carterson, A.J.; Honer zu Bentrup, K.; Ott, C.M.; Clarke, M.S.; Pierson, D.L.; Vanderburg, C.R.; Buchanan, K.L.; Nickerson, C.A.; Schurr, M.J. A549 lung epithelial cells grown as three-dimensional aggregates: Alternative tissue culture model for Pseudomonas aeruginosa pathogenesis. Infect. Immun. 2005, 73, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Deslee, G.; Dury, S.; Perotin, J.M.; Al Alam, D.; Vitry, F.; Boxio, R.; Gangloff, S.C.; Guenounou, M.; Lebargy, F.; Belaaouaj, A. Bronchial epithelial spheroids: An alternative culture model to investigate epithelium inflammation-mediated COPD. Respir. Res. 2007, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unsworth, B.R.; Lelkes, P.I. Growing tissues in microgravity. Nat. Med. 1998, 4, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Castillon, N.; Avril-Delplanque, A.; Coraux, C.; Delenda, C.; Peault, B.; Danos, O.; Puchelle, E. Regeneration of a well-differentiated human airway surface epithelium by spheroid and lentivirus vector-transduced airway cells. J. Gene Med. 2004, 6, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Castillon, N.; Hinnrasky, J.; Zahm, J.M.; Kaplan, H.; Bonnet, N.; Corlieu, P.; Klossek, J.M.; Taouil, K.; Avril-Delplanque, A.; Peault, B.; et al. Polarized expression of cystic fibrosis transmembrane conductance regulator and associated epithelial proteins during the regeneration of human airway surface epithelium in three-dimensional culture. Lab. Investig. 2002, 82, 989–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schogler, A.; Blank, F.; Brugger, M.; Beyeler, S.; Tschanz, S.A.; Regamey, N.; Casaulta, C.; Geiser, T.; Alves, M.P. Characterization of pediatric cystic fibrosis airway epithelial cell cultures at the air-liquid interface obtained by non-invasive nasal cytology brush sampling. Respir. Res. 2017, 18, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.C.; Thavagnanam, S.; Skibinski, G.; Lyons, J.; Bell, J.; Heaney, L.G.; Shields, M.D. Chronic IL9 and IL-13 exposure leads to an altered differentiation of ciliated cells in a well-differentiated paediatric bronchial epithelial cell model. PLoS ONE 2013, 8, e61023. [Google Scholar] [CrossRef] [Green Version]
- Brewington, J.J.; Filbrandt, E.T.; LaRosa, F.J., 3rd; Moncivaiz, J.D.; Ostmann, A.J.; Strecker, L.M.; Clancy, J.P. Generation of Human Nasal Epithelial Cell Spheroids for Individualized Cystic Fibrosis Transmembrane Conductance Regulator Study. J. Vis. Exp. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gamarra, F.; Bergner, A.; Stauss, E.; Stocker, I.; Grundler, S.; Huber, R.M. Rotation frequency of human bronchial and nasal epithelial spheroids as an indicator of mucociliary function. Respiration 2006, 73, 664–672. [Google Scholar] [CrossRef] [Green Version]
- Ebsen, M.; Mogilevski, G.; Anhenn, O.; Maiworm, V.; Theegarten, D.; Schwarze, J.; Morgenroth, K. Infection of murine precision cut lung slices (PCLS) with respiratory syncytial virus (RSV) and chlamydophila pneumoniae using the Krumdieck technique. Pathol. Res. Pract. 2002, 198, 747–753. [Google Scholar] [CrossRef]
- Liu, G.; Betts, C.; Cunoosamy, D.M.; Aberg, P.M.; Hornberg, J.J.; Sivars, K.B.; Cohen, T.S. Use of precision cut lung slices as a translational model for the study of lung biology. Respir. Res. 2019, 20, 162. [Google Scholar] [CrossRef] [PubMed]
- Faisy, C.; Grassin-Delyle, S.; Blouquit-Laye, S.; Brollo, M.; Naline, E.; Chapelier, A.; Devillier, P. Wnt/beta-catenin signaling modulates human airway sensitization induced by beta2-adrenoceptor stimulation. PLoS ONE 2014, 9, e111350. [Google Scholar] [CrossRef] [Green Version]
- Morin, C.; Sirois, M.; Echave, V.; Gomes, M.M.; Rousseau, E. EET displays anti-inflammatory effects in TNF-alpha stimulated human bronchi: Putative role of CPI-17. Am. J. Respir. Cell Mol. Biol. 2008, 38, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Carletti, E.; Motta, A.; Migliaresi, C. Scaffolds for tissue engineering and 3D cell culture. Methods Mol Biol 2011, 695, 17–39. [Google Scholar] [CrossRef]
- Nichols, J.E.; Niles, J.; Riddle, M.; Vargas, G.; Schilagard, T.; Ma, L.; Edward, K.; La Francesca, S.; Sakamoto, J.; Vega, S.; et al. Production and assessment of decellularized pig and human lung scaffolds. Tissue Eng Part. A 2013, 19, 2045–2062. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.H.; Calle, E.A.; Colehour, M.B.; Niklason, L.E. Matrix composition and mechanics of decellularized lung scaffolds. Cells Tissues Organs 2012, 195, 222–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Anderson, J.D.; Deng, L.; Mackay, S.; Bailey, J.; Kersh, L.; Rowe, S.M.; Guimbellot, J.S. Human Nasal Epithelial Organoids for Therapeutic Development in Cystic Fibrosis. Genes 2020, 11, 603. [Google Scholar] [CrossRef]
- Barkauskas, C.E.; Chung, M.I.; Fioret, B.; Gao, X.; Katsura, H.; Hogan, B.L. Lung organoids: Current uses and future promise. Development 2017, 144, 986–997. [Google Scholar] [CrossRef] [Green Version]
- Bartfeld, S.; Clevers, H. Stem cell-derived organoids and their application for medical research and patient treatment. J. Mol Med. (Berl) 2017, 95, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachs, N.; Papaspyropoulos, A.; Zomer-van Ommen, D.D.; Heo, I.; Bottinger, L.; Klay, D.; Weeber, F.; Huelsz-Prince, G.; Iakobachvili, N.; Amatngalim, G.D.; et al. Long-term expanding human airway organoids for disease modeling. EMBO J. 2019, 38. [Google Scholar] [CrossRef] [PubMed]
- Paschini, M.; Kim, C.F. An airway organoid is forever. EMBO J. 2019, 38. [Google Scholar] [CrossRef] [PubMed]
- Guimbellot, J.S.; Leach, J.M.; Chaudhry, I.G.; Quinney, N.L.; Boyles, S.E.; Chua, M.; Aban, I.; Jaspers, I.; Gentzsch, M. Nasospheroids permit measurements of CFTR-dependent fluid transport. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keglowich, L.; Roth, M.; Philippova, M.; Resink, T.; Tjin, G.; Oliver, B.; Lardinois, D.; Dessus-Babus, S.; Gosens, R.; Hostettler Haack, K.; et al. Bronchial smooth muscle cells of asthmatics promote angiogenesis through elevated secretion of CXC-chemokines (ENA-78, GRO-alpha, and IL-8). PLoS ONE 2013, 8, e81494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randell, S.H.; Fulcher, M.L.; O’Neal, W.; Olsen, J.C. Primary epithelial cell models for cystic fibrosis research. Methods Mol. Biol. 2011, 742, 285–310. [Google Scholar] [CrossRef] [PubMed]
- Comer, D.M.; Elborn, J.S.; Ennis, M. Comparison of nasal and bronchial epithelial cells obtained from patients with COPD. PLoS ONE 2012, 7, e32924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLellan, K.; Shields, M.; Power, U.; Turner, S. Primary airway epithelial cell culture and asthma in children-lessons learnt and yet to come. Pediatr. Pulmonol. 2015, 50, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Batra, H.; Yarmus, L. Indications and complications of rigid bronchoscopy. Expert Rev. Respir. Med. 2018, 12, 509–520. [Google Scholar] [CrossRef]
- Soyer, T. The role bronchoscopy in the diagnosis of airway disease in children. J. Thorac. Dis. 2016, 8, 3420–3426. [Google Scholar] [CrossRef] [Green Version]
- Webster, I.; Goussard, P.; Gie, R.; Janson, J.; Rossouw, G. The indications and role of paediatric bronchoscopy in a developing country, with high prevalence of pulmonary tuberculosis and HIV. Expert Rev. Respir. Med. 2017, 11, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Lane, C.; Burgess, S.; Kicic, A.; Knight, D.; Stick, S. The use of non-bronchoscopic brushings to study the paediatric airway. Respir. Res. 2005, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Doherty, G.M.; Christie, S.N.; Skibinski, G.; Puddicombe, S.M.; Warke, T.J.; de Courcey, F.; Cross, A.L.; Lyons, J.D.; Ennis, M.; Shields, M.D.; et al. Non-bronchoscopic sampling and culture of bronchial epithelial cells in children. Clin. Exp. Allergy 2003, 33, 1221–1225. [Google Scholar] [CrossRef]
- Muller, L.; Brighton, L.E.; Carson, J.L.; Fischer, W.A., 2nd; Jaspers, I. Culturing of human nasal epithelial cells at the air liquid interface. J. Vis. Exp. 2013. [Google Scholar] [CrossRef]
- Devalia, J.L.; Sapsford, R.J.; Wells, C.W.; Richman, P.; Davies, R.J. Culture and comparison of human bronchial and nasal epithelial cells in vitro. Respir. Med. 1990, 84, 303–312. [Google Scholar] [CrossRef]
- Maestre-Batlle, D.; Pena, O.M.; Hirota, J.A.; Gunawan, E.; Rider, C.F.; Sutherland, D.; Alexis, N.E.; Carlsten, C. Novel flow cytometry approach to identify bronchial epithelial cells from healthy human airways. Sci. Rep. 2017, 7, 42214. [Google Scholar] [CrossRef] [PubMed]
- Aydin, M.; Naumova, E.A.; Lutz, S.; Meyer-Bahlburg, A.; Arnold, W.H.; Kreppel, F.; Ehrhardt, A.; Postberg, J.; Wirth, S. Do Current Asthma-Preventive Measures Appropriately Face the World Health Organization’s Concerns: A Study Presentation of a New Clinical, Prospective, Multicentric Pediatric Asthma Exacerbation Cohort in Germany. Front. Pediatr. 2020, 8, 574462. [Google Scholar] [CrossRef]
- Zhang, W.; Fu, J.; Liu, J.; Wang, H.; Schiwon, M.; Janz, S.; Schaffarczyk, L.; von der Goltz, L.; Ehrke-Schulz, E.; Dorner, J.; et al. An Engineered Virus Library as a Resource for the Spectrum-wide Exploration of Virus and Vector Diversity. Cell Rep. 2017, 19, 1698–1709. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Mead, B.E.; Safaee, H.; Langer, R.; Karp, J.M.; Levy, O. Engineering Stem Cell Organoids. Cell Stem. Cell 2016, 18, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayner, R.E.; Makena, P.; Prasad, G.L.; Cormet-Boyaka, E. Optimization of Normal Human Bronchial Epithelial (NHBE) Cell 3D Cultures for in vitro Lung Model Studies. Sci. Rep. 2019, 9, 500. [Google Scholar] [CrossRef]
- Brodlie, M.; McKean, M.C.; Johnson, G.E.; Perry, J.D.; Nicholson, A.; Verdon, B.; Gray, M.A.; Dark, J.H.; Pearson, J.P.; Fisher, A.J.; et al. Primary bronchial epithelial cell culture from explanted cystic fibrosis lungs. Exp. Lung Res. 2010, 36, 101–110. [Google Scholar] [CrossRef]
- Fulcher, M.L.; Gabriel, S.; Burns, K.A.; Yankaskas, J.R.; Randell, S.H. Well-differentiated human airway epithelial cell cultures. Methods Mol. Med. 2005, 107, 183–206. [Google Scholar] [CrossRef]
- Iverson, E.; Kaler, L.; Agostino, E.L.; Song, D.; Duncan, G.A.; Scull, M.A. Leveraging 3D Model Systems to Understand Viral Interactions with the Respiratory Mucosa. Viruses 2020, 12, 1425. [Google Scholar] [CrossRef]
- Kimura, H.; Francisco, D.; Conway, M.; Martinez, F.D.; Vercelli, D.; Polverino, F.; Billheimer, D.; Kraft, M. Type 2 inflammation modulates ACE2 and TMPRSS2 in airway epithelial cells. J. Allergy Clin. Immunol. 2020, 146, 80–88.e88. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Guo, J.; Huang, H.; Xia, B.; Liu, H.; Li, J.; Lin, S.; Li, T.; Liu, J.; Li, H. Human normal bronchial epithelial cells: A novel in vitro cell model for toxicity evaluation. PLoS ONE 2015, 10, e0123520. [Google Scholar] [CrossRef]
- Jang, Y.J.; Lee, Y.H.; Shin, S.H. Rhinovirus-infected nasal polyp epithelial cells: Effect on the activation and migration of eosinophils by airborne fungi. Ann. Allergy Asthma Immunol. 2010, 104, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, L.; Manzoor, S.; Zarcone, M.C.; Barabas, J.; Shields, M.D.; Saglani, S.; Lloyd, C.M.; Bush, A.; Custovic, A.; Ghazal, P.; et al. Comparative primary paediatric nasal epithelial cell culture differentiation and RSV-induced cytopathogenesis following culture in two commercial media. PLoS ONE 2020, 15, e0228229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luengen, A.E.; Kniebs, C.; Buhl, E.M.; Cornelissen, C.G.; Schmitz-Rode, T.; Jockenhoevel, S.; Thiebes, A.L. Choosing the Right Differentiation Medium to Develop Mucociliary Phenotype of Primary Nasal Epithelial Cells In Vitro. Sci. Rep. 2020, 10, 6963. [Google Scholar] [CrossRef]
- Roberts, N.; Al Mubarak, R.; Francisco, D.; Kraft, M.; Chu, H.W. Comparison of paired human nasal and bronchial airway epithelial cell responses to rhinovirus infection and IL-13 treatment. Clin. Transl. Med. 2018, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Jones, N. The nose and paranasal sinuses physiology and anatomy. Adv. Drug Deliv. Rev. 2001, 51, 5–19. [Google Scholar] [CrossRef]
- Leopold, D.A. Pollution: The nose and sinuses. Otolaryngol Head Neck Surg 1992, 106, 713–719. [Google Scholar] [CrossRef]
- Jiao, J.; Wang, C.; Zhang, L. Epithelial physical barrier defects in chronic rhinosinusitis. Expert. Rev. Clin. Immunol. 2019, 15, 679–688. [Google Scholar] [CrossRef]
- Baturcam, E.; Snape, N.; Yeo, T.H.; Schagen, J.; Thomas, E.; Logan, J.; Galbraith, S.; Collinson, N.; Phipps, S.; Fantino, E.; et al. Human Metapneumovirus Impairs Apoptosis of Nasal Epithelial Cells in Asthma via HSP70. J. Innate Immun. 2017, 9, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Thavagnanam, S.; Parker, J.C.; McBrien, M.E.; Skibinski, G.; Shields, M.D.; Heaney, L.G. Nasal epithelial cells can act as a physiological surrogate for paediatric asthma studies. PLoS ONE 2014, 9, e85802. [Google Scholar] [CrossRef] [Green Version]
- Vanders, R.L.; Hsu, A.; Gibson, P.G.; Murphy, V.E.; Wark, P.A.B. Nasal epithelial cells to assess in vitro immune responses to respiratory virus infection in pregnant women with asthma. Respir. Res. 2019, 20, 259. [Google Scholar] [CrossRef]
- Driehuis, E.; Kretzschmar, K.; Clevers, H. Establishment of patient-derived cancer organoids for drug-screening applications. Nat. Protoc. 2020, 15, 3380–3409. [Google Scholar] [CrossRef] [PubMed]
- VanDussen, K.L.; Marinshaw, J.M.; Shaikh, N.; Miyoshi, H.; Moon, C.; Tarr, P.I.; Ciorba, M.A.; Stappenbeck, T.S. Development of an enhanced human gastrointestinal epithelial culture system to facilitate patient-based assays. Gut 2015, 64, 911–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, D.J.; Makrinioti, H.; Rana, B.M.; Shamji, B.W.; Trujillo-Torralbo, M.B.; Footitt, J.; Jerico, D.-R.; Telcian, A.G.; Nikonova, A.; Zhu, J.; et al. IL-33-dependent type 2 inflammation during rhinovirus-induced asthma exacerbations in vivo. Am. J. Respir Crit. Care Med. 2014, 190, 1373–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pech, M.; Weckmann, M.; Konig, I.R.; Franke, A.; Heinsen, F.A.; Oliver, B.; Ricklefs, I.; Fuchs, O.; Rabe, K.; Hansen, G.; et al. Rhinovirus infections change DNA methylation and mRNA expression in children with asthma. PLoS ONE 2018, 13, e0205275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaya, M.; Nomura, K.; Arakawa, K.; Sugawara, M.; Deng, X.; Lusamba Kalonji, N.; Nishimura, H.; Yamada, M.; Nagatomi, R.; Kawase, T. Clarithromycin decreases rhinovirus replication and cytokine production in nasal epithelial cells from subjects with bronchial asthma: Effects on IL-6, IL-8 and IL-33. Arch. Pharm Res. 2020, 43, 526–539. [Google Scholar] [CrossRef]
- Lopez-Souza, N.; Favoreto, S.; Wong, H.; Ward, T.; Yagi, S.; Schnurr, D.; Finkbeiner, W.E.; Dolganov, G.M.; Widdicombe, J.H.; Boushey, H.A.; et al. In vitro susceptibility to rhinovirus infection is greater for bronchial than for nasal airway epithelial cells in human subjects. J. Allergy Clin. Immunol. 2009, 123, 1384–1390.e1382. [Google Scholar] [CrossRef] [Green Version]
- Spann, K.M.; Baturcam, E.; Schagen, J.; Jones, C.; Straub, C.P.; Preston, F.M.; Chen, L.; Phipps, S.; Sly, P.D.; Fantino, E. Viral and host factors determine innate immune responses in airway epithelial cells from children with wheeze and atopy. Thorax 2014, 69, 918–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McErlean, P.; Favoreto, S., Jr.; Costa, F.F.; Shen, J.; Quraishi, J.; Biyasheva, A.; Cooper, J.J.; Scholtens, D.M.; Vanin, E.F.; de Bonaldo, M.F.; et al. Human rhinovirus infection causes different DNA methylation changes in nasal epithelial cells from healthy and asthmatic subjects. BMC Med. Genomics 2014, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaya, M.; Nishimura, H.; Deng, X.; Sugawara, M.; Watanabe, O.; Nomura, K.; Shimotai, Y.; Momma, H.; Ichinose, M.; Kawase, T. Inhibitory effects of glycopyrronium, formoterol, and budesonide on coronavirus HCoV-229E replication and cytokine production by primary cultures of human nasal and tracheal epithelial cells. Respir. Investig. 2020, 58, 155–168. [Google Scholar] [CrossRef] [PubMed]




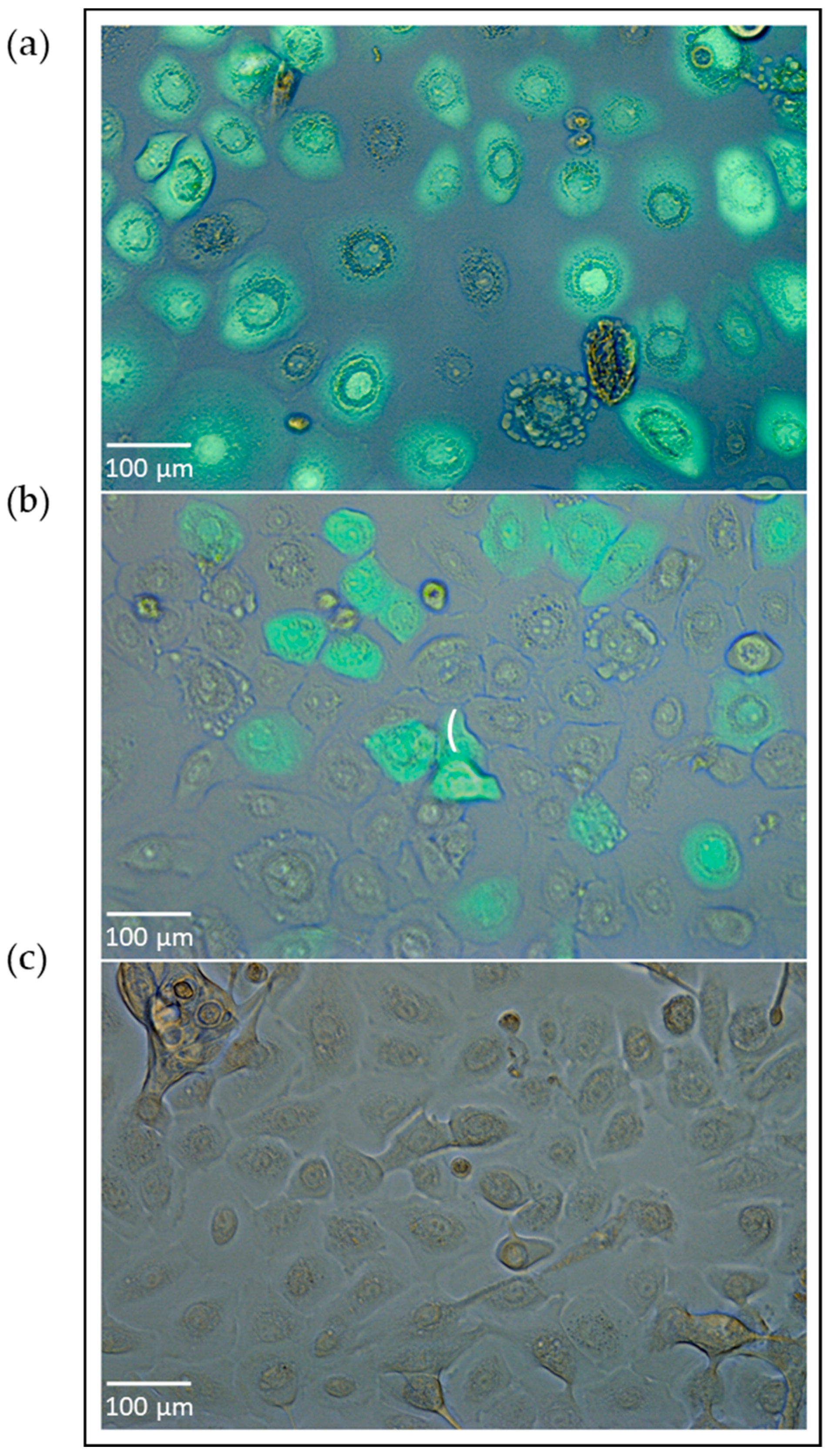{kind=link}
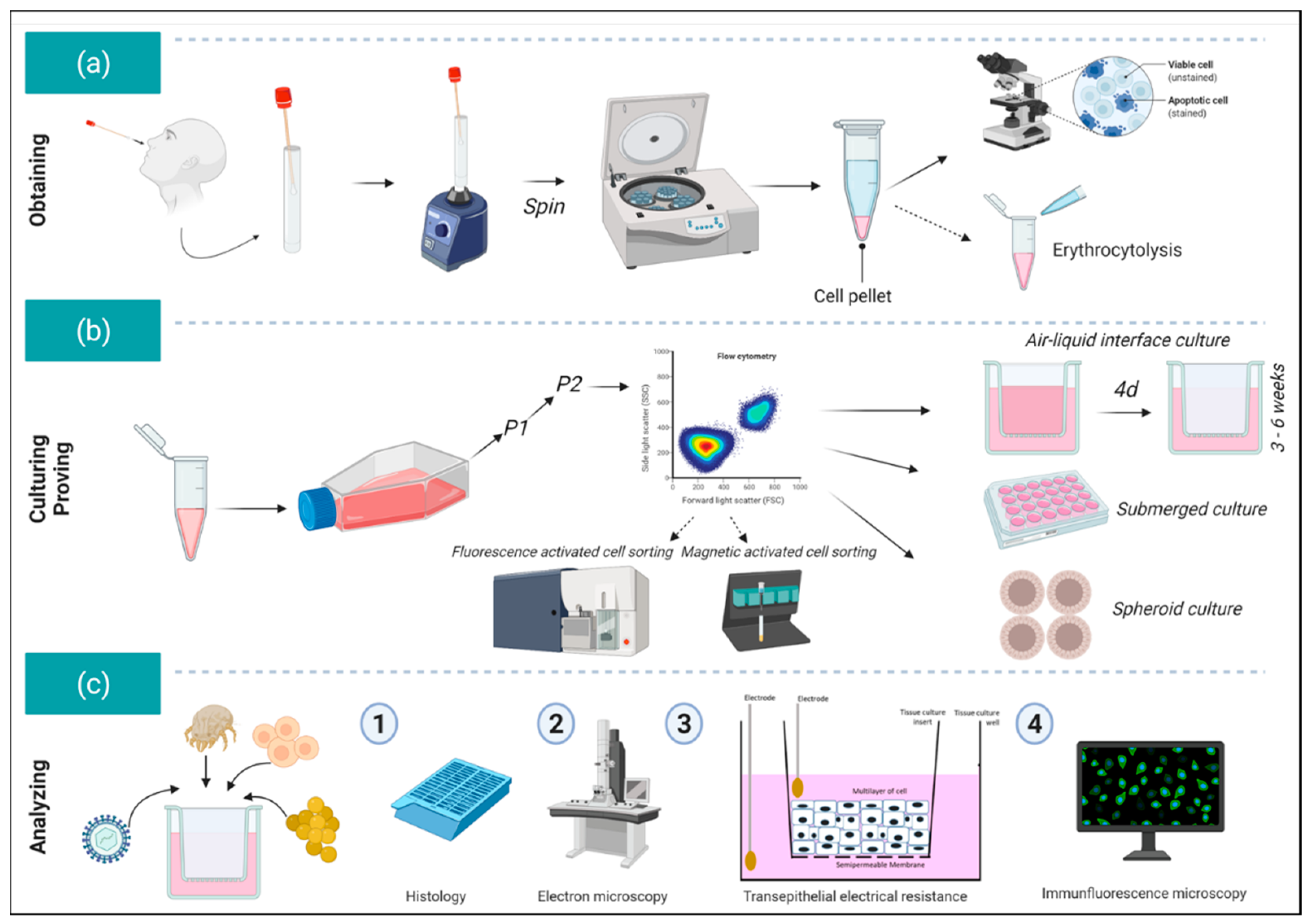{kind=link}
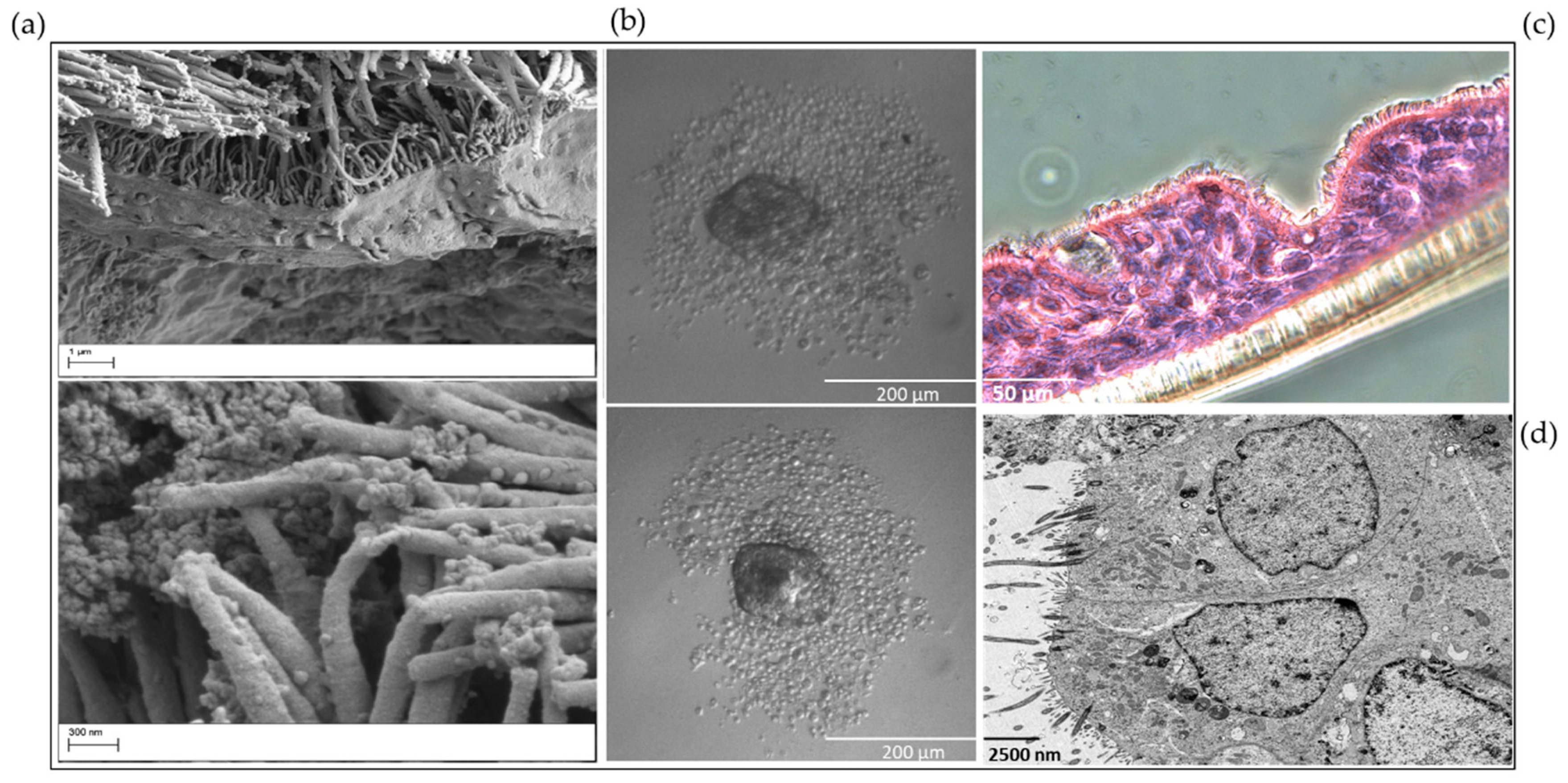{kind=link}
| Research Article | Virus | Disease | Type of Nasal Epithelial Cell Culture |
|---|---|---|---|
| Pech and colleagues (2018) [118] Rhinovirus infections change DNA methylation and mRNA expression in children with asthma | RV 1 16 | Asthma | submerged |
| Yamaya and colleagues (2020) [119] Clarithromycin decreases rhinovirus replication and cytokine production in nasal epithelial cells from subjects with bronchial asthma: effects on IL-6, IL-8 and IL-33 | RV 14 | Asthma | submerged |
| Lopez-Souza and colleagues (2009) [120] In vitro susceptibility to rhinovirus infection is greater for bronchial than for nasal airway epithelial cells in human subjects | RV 16 | Asthma | ALI 2 |
| Spann and colleagues (2014) [121] Viral and host factors determine innate immune responses in airway epithelial cells from children with wheeze and atopy | RSV 3 & HMPV 4 | Wheeze/Atopy | submerged |
| McErlean and colleagues (2014) [122] Human rhinovirus infection causes different DNA methylation changes in nasal epithelial cells from healthy and asthmatic subjects | RV | Asthma | submerged |
| Baturcam and colleagues (2017) [112] Human Metapneumovirus Impairs Apoptosis of Nasal Epithelial Cells in Asthma via HSP70 | RSV & HMPV | Asthma | submerged |
| Vanders and colleagues (2019) [114] Nasal epithelial cells to assess in vitro immune responses to respiratory virus infection in pregnant women with asthma | RV 43 & H3N2 5 | Asthma | submerged and ALI |
| Aydin and colleagues (2020) [22] House Dust Mite Exposure Causes Increased Susceptibility of Nasal Epithelial Cells to Adenovirus Infection. | AdV 6 | Asthma | submerged and ALI |
| Yamaya and colleagues (2020) [123] Inhibitory effects of glycopyrronium, formoterol, and budesonide on coronavirus HCoV-229E replication and cytokine production by primary cultures of human nasal and tracheal epithelial cells | HCoV-229E 7 | among others Asthma | ALI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aydin, M.; Naumova, E.A.; Bellm, A.; Behrendt, A.-K.; Giachero, F.; Bahlmann, N.; Zhang, W.; Wirth, S.; Paulsen, F.; Arnold, W.H.; et al. From Submerged Cultures to 3D Cell Culture Models: Evolution of Nasal Epithelial Cells in Asthma Research and Virus Infection. Viruses 2021, 13, 387. https://doi.org/10.3390/v13030387
Aydin M, Naumova EA, Bellm A, Behrendt A-K, Giachero F, Bahlmann N, Zhang W, Wirth S, Paulsen F, Arnold WH, et al. From Submerged Cultures to 3D Cell Culture Models: Evolution of Nasal Epithelial Cells in Asthma Research and Virus Infection. Viruses. 2021; 13(3):387. https://doi.org/10.3390/v13030387
Chicago/Turabian StyleAydin, Malik, Ella A. Naumova, Aliyah Bellm, Ann-Kathrin Behrendt, Federica Giachero, Nora Bahlmann, Wenli Zhang, Stefan Wirth, Friedrich Paulsen, Wolfgang H. Arnold, and et al. 2021. "From Submerged Cultures to 3D Cell Culture Models: Evolution of Nasal Epithelial Cells in Asthma Research and Virus Infection" Viruses 13, no. 3: 387. https://doi.org/10.3390/v13030387
APA StyleAydin, M., Naumova, E. A., Bellm, A., Behrendt, A. -K., Giachero, F., Bahlmann, N., Zhang, W., Wirth, S., Paulsen, F., Arnold, W. H., & Ehrhardt, A. (2021). From Submerged Cultures to 3D Cell Culture Models: Evolution of Nasal Epithelial Cells in Asthma Research and Virus Infection. Viruses, 13(3), 387. https://doi.org/10.3390/v13030387