
In Vivo Models of HDV Infection: Is Humanizing NTCP Enough?

Abstract
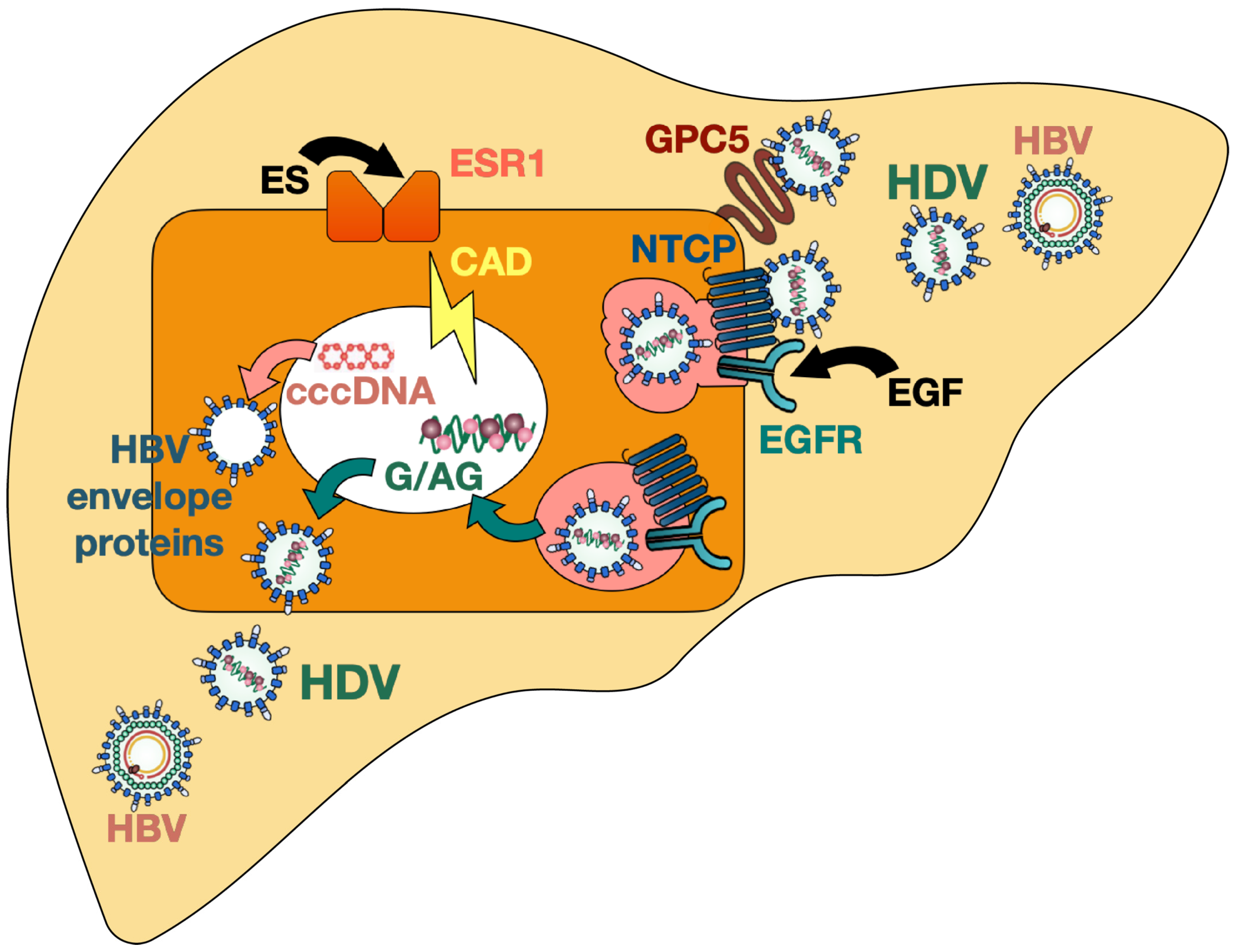
:1. Hepatitis Delta Virus
2. Natural Hosts of HDV and HDV-Related Viruses
3. In Vivo Models to Study HDV Infection
3.1. Early HDV Experiments in Mice
3.2. Transgenic Mouse Models
3.3. Hydrodynamic Mouse Models
3.4. Human Liver Chimeric Mice
3.5. AAV-Based Mouse Models
3.6. NTCP-Based Mouse Models
4. HDV Host Restrictions Factors
5. Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rizzetto, M.; Canese, M.G.; Aricò, S.; Crivelli, O.; Trepo, C.; Bonino, F.; Verme, G. Immunofluorescence Detection of New Antigen-Antibody System (Delta/Anti-Delta) Associated to Hepatitis B Virus in Liver and in Serum of HBsAg Carriers. Gut 1977, 18, 997–1003. [Google Scholar] [CrossRef] [Green Version]
- Vlachogiannakos, J.; Papatheodoridis, G.V. New Epidemiology of Hepatitis Delta. Liver Int. 2020, 40 (Suppl. 1), 48–53. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-Y.; Shen, D.-T.; Ji, D.-Z.; Han, P.-C.; Zhang, W.-M.; Ma, J.-F.; Chen, W.-S.; Goyal, H.; Pan, S.; Xu, H.-G. Prevalence and Burden of Hepatitis D Virus Infection in the Global Population: A Systematic Review and Meta-Analysis. Gut 2019, 68, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Rizzetto, M.; Hamid, S.; Negro, F. The Changing Scenario of Hepatitis D. J. Hepatol. 2021. [Google Scholar] [CrossRef]
- Fattovich, G.; Giustina, G.; Christensen, E.; Pantalena, M.; Zagni, I.; Realdi, G.; Schalm, S.W. Influence of Hepatitis Delta Virus Infection on Morbidity and Mortality in Compensated Cirrhosis Type B. The European Concerted Action on Viral Hepatitis (Eurohep). Gut 2000, 46, 420–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, C.; Heller, T.; Glenn, J.S. Pathogenesis of and New Therapies for Hepatitis D. Gastroenterology 2019, 156, 461–476.e1. [Google Scholar] [CrossRef]
- Weiner, A.J.; Choo, Q.L.; Wang, K.S.; Govindarajan, S.; Redeker, A.G.; Gerin, J.L.; Houghton, M. A Single Antigenomic Open Reading Frame of the Hepatitis Delta Virus Encodes the Epitope(s) of Both Hepatitis Delta Antigen Polypeptides P24 Delta and P27 Delta. J. Virol. 1988, 62, 594–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudima, S.; Chang, J.; Moraleda, G.; Azvolinsky, A.; Taylor, J. Parameters of Human Hepatitis Delta Virus Genome Replication: The Quantity, Quality, and Intracellular Distribution of Viral Proteins and RNA. J. Virol. 2002, 76, 3709–3719. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.S.; Choo, Q.L.; Weiner, A.J.; Ou, J.H.; Najarian, R.C.; Thayer, R.M.; Mullenbach, G.T.; Denniston, K.J.; Gerin, J.L.; Houghton, M. Structure, Sequence and Expression of the Hepatitis Delta (Delta) Viral Genome. Nature 1986, 323, 508–514. [Google Scholar] [CrossRef]
- Lai, M.M.C. RNA Replication without RNA-Dependent RNA Polymerase: Surprises from Hepatitis Delta Virus. J. Virol. 2005, 79, 7951–7958. [Google Scholar] [CrossRef] [Green Version]
- Chang, F.L.; Chen, P.J.; Tu, S.J.; Wang, C.J.; Chen, D.S. The Large Form of Hepatitis Delta Antigen Is Crucial for Assembly of Hepatitis Delta Virus. Proc. Natl. Acad. Sci. USA 1991, 88, 8490–8494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.J.; Chen, P.J.; Wu, J.C.; Patel, D.; Chen, D.S. Small-Form Hepatitis B Surface Antigen is Sufficient to Help in the Assembly of Hepatitis Delta Virus-like Particles. J. Virol. 1991, 65, 6630–6636. [Google Scholar] [CrossRef] [Green Version]
- Lucifora, J.; Delphin, M. Current Knowledge on Hepatitis Delta Virus Replication. Antivir. Res. 2020, 179, 104812. [Google Scholar] [CrossRef]
- Huang, W.H.; Chen, C.W.; Wu, H.L.; Chen, P.J. Post-Translational Modification of Delta Antigen of Hepatitis D Virus. Curr. Top Microbiol. Immunol. 2006, 307, 91–112. [Google Scholar] [CrossRef]
- Rizzetto, M.; Hoyer, B.; Canese, M.G.; Shih, J.W.; Purcell, R.H.; Gerin, J.L. Delta Agent: Association of Delta Antigen with Hepatitis B Surface Antigen and RNA in Serum of Delta-Infected Chimpanzees. Proc. Natl. Acad. Sci. USA 1980, 77, 6124–6128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lempp, F.A.; Ni, Y.; Urban, S. Hepatitis Delta Virus: Insights into a Peculiar Pathogen and Novel Treatment Options. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Perez-Vargas, J.; Amirache, F.; Boson, B.; Mialon, C.; Freitas, N.; Sureau, C.; Fusil, F.; Cosset, F.-L. Enveloped Viruses Distinct from HBV Induce Dissemination of Hepatitis D Virus in Vivo. Nat. Commun. 2019, 10, 2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappy, P.; Lucas, Q.; Kankarafou, N.; Sureau, C.; Laperche, S. No Evidence of HCV-Assisted HDV Propagation in a Large Cohort of Hepatitis C Positive Blood Donors. J. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chemin, I.; Pujol, F.H.; Scholtès, C.; Loureiro, C.L.; Amirache, F.; Levrero, M.; Zoulim, F.; Pérez-Vargas, J.; Cosset, F.-L. Preliminary Evidence for Hepatitis Delta Virus Exposure in Patients Who Are Apparently Not Infected With Hepatitis B Virus. Hepatology 2021, 73, 861–864. [Google Scholar] [CrossRef]
- Pflüger, L.S.; Schulze Zur Wiesch, J.; Polywka, S.; Lütgehetmann, M. Hepatitis Delta Virus Propagation Enabled by Hepatitis C Virus-Scientifically Intriguing, but Is It Relevant to Clinical Practice? J. Viral Hepat. 2021, 28, 213–216. [Google Scholar] [CrossRef]
- Pollicino, T.; Raffa, G.; Santantonio, T.; Gaeta, G.B.; Iannello, G.; Alibrandi, A.; Squadrito, G.; Cacciola, I.; Calvi, C.; Colucci, G.; et al. Replicative and Transcriptional Activities of Hepatitis B Virus in Patients Coinfected with Hepatitis B and Hepatitis Delta Viruses. J. Virol. 2011, 85, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Schaper, M.; Rodriguez-Frias, F.; Jardi, R.; Tabernero, D.; Homs, M.; Ruiz, G.; Quer, J.; Esteban, R.; Buti, M. Quantitative Longitudinal Evaluations of Hepatitis Delta Virus RNA and Hepatitis B Virus DNA Shows a Dynamic, Complex Replicative Profile in Chronic Hepatitis B and D. J. Hepatol. 2010, 52, 658–664. [Google Scholar] [CrossRef]
- Mederacke, I.; Filmann, N.; Yurdaydin, C.; Bremer, B.; Puls, F.; Zacher, B.J.; Heidrich, B.; Tillmann, H.L.; Rosenau, J.; Bock, C.-T.; et al. Rapid Early HDV RNA Decline in the Peripheral Blood but Prolonged Intrahepatic Hepatitis Delta Antigen Persistence after Liver Transplantation. J. Hepatol. 2012, 56, 115–122. [Google Scholar] [CrossRef]
- Samuel, D.; Zignego, A.L.; Reynes, M.; Feray, C.; Arulnaden, J.L.; David, M.F.; Gigou, M.; Bismuth, A.; Mathieu, D.; Gentilini, P. Long-Term Clinical and Virological Outcome after Liver Transplantation for Cirrhosis Caused by Chronic Delta Hepatitis. Hepatology 1995, 21, 333–339. [Google Scholar] [CrossRef]
- Le Gal, F.; Gault, E.; Ripault, M.-P.; Serpaggi, J.; Trinchet, J.-C.; Gordien, E.; Dény, P. Eighth Major Clade for Hepatitis Delta Virus. Emerg. Infect. Dis. 2006, 12, 1447–1450. [Google Scholar] [CrossRef]
- Le Gal, F.; Brichler, S.; Drugan, T.; Alloui, C.; Roulot, D.; Pawlotsky, J.-M.; Dény, P.; Gordien, E. Genetic Diversity and Worldwide Distribution of the Deltavirus Genus: A Study of 2,152 Clinical Strains. Hepatology 2017, 66, 1826–1841. [Google Scholar] [CrossRef] [Green Version]
- Casey, J.L.; Niro, G.A.; Engle, R.E.; Vega, A.; Gomez, H.; McCarthy, M.; Watts, D.M.; Hyams, K.C.; Gerin, J.L. Hepatitis B Virus (HBV)/Hepatitis D Virus (HDV) Coinfection in Outbreaks of Acute Hepatitis in the Peruvian Amazon Basin: The Roles of HDV Genotype III and HBV Genotype F. J. Infect. Dis. 1996, 174, 920–926. [Google Scholar] [CrossRef]
- Yurdaydin, C.; Abbas, Z.; Buti, M.; Cornberg, M.; Esteban, R.; Etzion, O.; Gane, E.J.; Gish, R.G.; Glenn, J.S.; Hamid, S.; et al. Treating Chronic Hepatitis Delta: The Need for Surrogate Markers of Treatment Efficacy. J. Hepatol. 2019, 70, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Dény, P. Hepatitis Delta Virus Genetic Variability: From Genotypes I, II, III to Eight Major Clades? Curr. Top Microbiol. Immunol. 2006, 307, 151–171. [Google Scholar] [CrossRef]
- Castelnau, C.; Le Gal, F.; Ripault, M.-P.; Gordien, E.; Martinot-Peignoux, M.; Boyer, N.; Pham, B.-N.; Maylin, S.; Bedossa, P.; Dény, P.; et al. Efficacy of Peginterferon Alpha-2b in Chronic Hepatitis Delta: Relevance of Quantitative RT-PCR for Follow-Up. Hepatology 2006, 44, 728–735. [Google Scholar] [CrossRef]
- Bremer, B.; Anastasiou, O.E.; Hardtke, S.; Caruntu, F.A.; Curescu, M.G.; Yalcin, K.; Akarca, U.S.; Gürel, S.; Zeuzem, S.; Erhardt, A.; et al. Residual Low HDV Viraemia Is Associated HDV RNA Relapse after PEG-IFNa-Based Antiviral Treatment of Hepatitis Delta: Results from the HIDIT-II Study. Liver Int. 2021, 41, 295–299. [Google Scholar] [CrossRef]
- Gheorghe, L.; Iacob, S.; Simionov, I.; Vadan, R.; Constantinescu, I.; Caruntu, F.; Sporea, I.; Grigorescu, M. Weight-Based Dosing Regimen of Peg-Interferon α-2b for Chronic Hepatitis Delta: A Multicenter Romanian Trial. J. Gastrointest. Liver Dis. 2011, 20, 377–382. [Google Scholar]
- Wedemeyer, H.; Yurdaydìn, C.; Dalekos, G.N.; Erhardt, A.; Çakaloğlu, Y.; Değertekin, H.; Gürel, S.; Zeuzem, S.; Zachou, K.; Bozkaya, H.; et al. Peginterferon plus Adefovir versus Either Drug Alone for Hepatitis Delta. N. Engl. J. Med. 2011, 364, 322–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niro, G.A.; Ciancio, A.; Gaeta, G.B.; Smedile, A.; Marrone, A.; Olivero, A.; Stanzione, M.; David, E.; Brancaccio, G.; Fontana, R.; et al. Pegylated Interferon Alpha-2b as Monotherapy or in Combination with Ribavirin in Chronic Hepatitis Delta. Hepatology 2006, 44, 713–720. [Google Scholar] [CrossRef]
- Erhardt, A.; Gerlich, W.; Starke, C.; Wend, U.; Donner, A.; Sagir, A.; Heintges, T.; Häussinger, D. Treatment of Chronic Hepatitis Delta with Pegylated Interferon-Alpha2b. Liver Int. 2006, 26, 805–810. [Google Scholar] [CrossRef]
- Bogomolov, P.; Alexandrov, A.; Voronkova, N.; Macievich, M.; Kokina, K.; Petrachenkova, M.; Lehr, T.; Lempp, F.A.; Wedemeyer, H.; Haag, M.; et al. Treatment of Chronic Hepatitis D with the Entry Inhibitor Myrcludex B: First Results of a Phase Ib/IIa Study. J. Hepatol. 2016, 65, 490–498. [Google Scholar] [CrossRef]
- Kang, C.; Syed, Y.Y. Bulevirtide: First Approval. Drugs 2020, 80, 1601–1605. [Google Scholar] [CrossRef]
- Caviglia, G.P.; Rizzetto, M. Treatment of Hepatitis D: An Unmet Medical Need. Clin. Microbiol. Infect. 2020, 26, 824–827. [Google Scholar] [CrossRef]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium Taurocholate Cotransporting Polypeptide Is a Functional Receptor for Human Hepatitis B and D Virus. Elife 2012, 1, e00049. [Google Scholar] [CrossRef]
- Longarela, O.L.; Schmidt, T.T.; Schöneweis, K.; Romeo, R.; Wedemeyer, H.; Urban, S.; Schulze, A. Proteoglycans Act as Cellular Hepatitis Delta Virus Attachment Receptors. PLoS ONE 2013, 8, e58340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, Y.; Lempp, F.A.; Mehrle, S.; Nkongolo, S.; Kaufman, C.; Fälth, M.; Stindt, J.; Königer, C.; Nassal, M.; Kubitz, R.; et al. Hepatitis B and D Viruses Exploit Sodium Taurocholate Co-Transporting Polypeptide for Species-Specific Entry into Hepatocytes. Gastroenterology 2014, 146, 1070–1083. [Google Scholar] [CrossRef]
- Xia, Y.P.; Yeh, C.T.; Ou, J.H.; Lai, M.M. Characterization of Nuclear Targeting Signal of Hepatitis Delta Antigen: Nuclear Transport as a Protein Complex. J. Virol. 1992, 66, 914–921. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Ding, M.; Wang, H. The infection of hepatitis D virus in adult tupaia. Zhonghua Yi Xue Za Zhi 1995, 75, 611–613, 639–640. [Google Scholar] [PubMed]
- Negro, F.; Bergmann, K.F.; Baroudy, B.M.; Satterfield, W.C.; Popper, H.; Purcell, R.H.; Gerin, J.L. Chronic Hepatitis D Virus (HDV) Infection in Hepatitis B Virus Carrier Chimpanzees Experimentally Superinfected with HDV. J. Infect. Dis. 1988, 158, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Negro, F.; Shapiro, M.; Satterfield, W.C.; Gerin, J.L.; Purcell, R.H. Reappearance of Hepatitis D Virus (HDV) Replication in Chronic Hepatitis B Virus Carrier Chimpanzees Rechallenged with HDV. J. Infect. Dis. 1989, 160, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Engle, R.E.; De Battista, D.; Danoff, E.J.; Nguyen, H.; Chen, Z.; Lusso, P.; Purcell, R.H.; Farci, P. Distinct Cytokine Profiles Correlate with Disease Severity and Outcome in Longitudinal Studies of Acute Hepatitis B Virus and Hepatitis D Virus Infection in Chimpanzees. mBio 2020, 11. [Google Scholar] [CrossRef]
- Barrera, A.; Guerra, B.; Lee, H.; Lanford, R.E. Analysis of Host Range Phenotypes of Primate Hepadnaviruses by in Vitro Infections of Hepatitis D Virus Pseudotypes. J. Virol. 2004, 78, 5233–5243. [Google Scholar] [CrossRef] [Green Version]
- Lempp, F.A.; Wiedtke, E.; Qu, B.; Roques, P.; Chemin, I.; Vondran, F.W.R.; Le Grand, R.; Grimm, D.; Urban, S. Sodium Taurocholate Cotransporting Polypeptide is the Limiting Host Factor of Hepatitis B Virus Infection in Macaque and Pig Hepatocytes. Hepatology 2017, 66, 703–716. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Peng, B.; He, W.; Zhong, G.; Qi, Y.; Ren, B.; Gao, Z.; Jing, Z.; Song, M.; Xu, G.; et al. Molecular Determinants of Hepatitis B and D Virus Entry Restriction in Mouse Sodium Taurocholate Cotransporting Polypeptide. J. Virol. 2013, 87, 7977–7991. [Google Scholar] [CrossRef] [Green Version]
- Caselmann, W.H. HBV and HDV Replication in Experimental Models: Effect of Interferon. Antivir. Res. 1994, 24, 121–129. [Google Scholar] [CrossRef]
- Ponzetto, A.; Cote, P.J.; Popper, H.; Hoyer, B.H.; London, W.T.; Ford, E.C.; Bonino, F.; Purcell, R.H.; Gerin, J.L. Transmission of the Hepatitis B Virus-Associated Delta Agent to the Eastern Woodchuck. Proc. Natl. Acad. Sci. USA 1984, 81, 2208–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlipköter, U.; Ponzetto, A.; Fuchs, K.; Rasshofer, R.; Choi, S.S.; Roos, S.; Rapicetta, M.; Roggendorf, M. Different Outcomes of Chronic Hepatitis Delta Virus Infection in Woodchucks. Liver 1990, 10, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Ciccaglione, A.R.; Rapicetta, M.; Fabiano, A.; Argentini, C.; Silvestro, M.; Giuseppetti, R.; Varano, F.; D’Urso, N.; Dinolfo, L.; Morgando, A. Chronic Infection in Woodchucks Infected by a Cloned Hepatitis Delta Virus. Arch. Virol. Suppl. 1993, 8, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Lukash, T.; Freitas, N.; Menne, S.; Gudima, S.O. Down-Regulation of Hepatitis Delta Virus Super-Infection in the Woodchuck Model. Virology 2019, 531, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, M.; Lu, M.; Siegel, F.; Whipple, J.; Roggendorf, M. Immunization of Woodchucks (Marmota Monax) with Hepatitis Delta Virus DNA Vaccine. Vaccine 2001, 19, 4618–4626. [Google Scholar] [CrossRef]
- Fiedler, M.; Kosinska, A.; Schumann, A.; Brovko, O.; Walker, A.; Lu, M.; Johrden, L.; Mayer, A.; Wildner, O.; Roggendorf, M. Prime/Boost Immunization with DNA and Adenoviral Vectors Protects from Hepatitis D Virus (HDV) Infection after Simultaneous Infection with HDV and Woodchuck Hepatitis Virus. J. Virol. 2013, 87, 7708–7716. [Google Scholar] [CrossRef] [Green Version]
- D’Ugo, E.; Paroli, M.; Palmieri, G.; Giuseppetti, R.; Argentini, C.; Tritarelli, E.; Bruni, R.; Barnaba, V.; Houghton, M.; Rapicetta, M. Immunization of Woodchucks with Adjuvanted SHDAg (P24): Immune Response and Outcome Following Challenge. Vaccine 2004, 22, 457–466. [Google Scholar] [CrossRef]
- Paraskevopoulou, S.; Pirzer, F.; Goldmann, N.; Schmid, J.; Corman, V.M.; Gottula, L.T.; Schroeder, S.; Rasche, A.; Muth, D.; Drexler, J.F.; et al. Mammalian Deltavirus without Hepadnavirus Coinfection in the Neotropical Rodent Proechimys Semispinosus. Proc. Natl. Acad. Sci. USA 2020, 117, 17977–17983. [Google Scholar] [CrossRef] [PubMed]
- Hetzel, U.; Szirovicza, L.; Smura, T.; Prähauser, B.; Vapalahti, O.; Kipar, A.; Hepojoki, J. Identification of a Novel Deltavirus in Boa Constrictors. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Szirovicza, L.; Hetzel, U.; Kipar, A.; Martinez-Sobrido, L.; Vapalahti, O.; Hepojoki, J. Snake Deltavirus Utilizes Envelope Proteins of Different Viruses To Generate Infectious Particles. mBio 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Wille, M.; Netter, H.J.; Littlejohn, M.; Yuen, L.; Shi, M.; Eden, J.-S.; Klaassen, M.; Holmes, E.C.; Hurt, A.C. A Divergent Hepatitis D-Like Agent in Birds. Viruses 2018, 10, 720. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.-S.; Pettersson, J.H.-O.; Le Lay, C.; Shi, M.; Lo, N.; Wille, M.; Eden, J.-S.; Holmes, E.C. Novel Hepatitis D-like Agents in Vertebrates and Invertebrates. Virus Evol. 2019, 5, vez021. [Google Scholar] [CrossRef]
- Iwamoto, M.; Shibata, Y.; Kawasaki, J.; Kojima, S.; Li, Y.-T.; Iwami, S.; Muramatsu, M.; Wu, H.-L.; Wada, K.; Tomonaga, K.; et al. Identification of Novel Avian and Mammalian Deltaviruses Provides New Insights into Deltavirus Evolution. Virus Evol. 2021, 7. [Google Scholar] [CrossRef]
- Netter, H.J.; Kajino, K.; Taylor, J.M. Experimental Transmission of Human Hepatitis Delta Virus to the Laboratory Mouse. J. Virol. 1993, 67, 3357–3362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polo, J.M.; Lim, B.; Govindarajan, S.; Lai, M.M. Replication of Hepatitis Delta Virus RNA in Mice after Intramuscular Injection of Plasmid DNA. J. Virol. 1995, 69, 5203–5207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilhot, S.; Huang, S.N.; Xia, Y.P.; La Monica, N.; Lai, M.M.; Chisari, F.V. Expression of the Hepatitis Delta Virus Large and Small Antigens in Transgenic Mice. J. Virol. 1994, 68, 1052–1058. [Google Scholar] [CrossRef] [Green Version]
- Polo, J.M.; Jeng, K.S.; Lim, B.; Govindarajan, S.; Hofman, F.; Sangiorgi, F.; Lai, M.M. Transgenic Mice Support Replication of Hepatitis Delta Virus RNA in Multiple Tissues, Particularly in Skeletal Muscle. J. Virol. 1995, 69, 4880–4887. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Song, Y.; Liu, D. Hydrodynamics-Based Transfection in Animals by Systemic Administration of Plasmid DNA. Gene Ther. 1999, 6, 1258–1266. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Sigal, L.J.; Lerro, A.; Taylor, J. Replication of the Human Hepatitis Delta Virus Genome Is Initiated in Mouse Hepatocytes Following Intravenous Injection of Naked DNA or RNA Sequences. J. Virol. 2001, 75, 3469–3473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dandri, M.; Lütgehetmann, M. Mouse Models of Hepatitis B and Delta Virus Infection. J. Immunol. Methods 2014, 410, 39–49. [Google Scholar] [CrossRef]
- Jirtle, R.L.; Biles, C.; Michalopoulos, G. Morphologic and Histochemical Analysis of Hepatocytes Transplanted into Syngeneic Hosts. Am. J. Pathol. 1980, 101, 115–126. [Google Scholar]
- Kusano, M.; Mito, M. Observations on the Fine Structure of Long-Survived Isolated Hepatocytes Inoculated into Rat Spleen. Gastroenterology 1982, 82, 616–628. [Google Scholar] [CrossRef]
- Demetriou, A.A.; Levenson, S.M.; Novikoff, P.M.; Novikoff, A.B.; Chowdhury, N.R.; Whiting, J.; Reisner, A.; Chowdhury, J.R. Survival, Organization, and Function of Microcarrier-Attached Hepatocytes Transplanted in Rats. Proc. Natl. Acad. Sci. USA 1986, 83, 7475–7479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricordi, C.; Lacy, P.E.; Callery, M.P.; Park, P.W.; Flye, M.W. Trophic Factors from Pancreatic Islets in Combined Hepatocyte-Islet Allografts Enhance Hepatocellular Survival. Surgery 1989, 105, 218–223. [Google Scholar]
- Ohashi, K.; Marion, P.L.; Nakai, H.; Meuse, L.; Cullen, J.M.; Bordier, B.B.; Schwall, R.; Greenberg, H.B.; Glenn, J.S.; Kay, M.A. Sustained Survival of Human Hepatocytes in Mice: A Model for in Vivo Infection with Human Hepatitis B and Hepatitis Delta Viruses. Nat. Med. 2000, 6, 327–331. [Google Scholar] [CrossRef]
- Heckel, J.L.; Sandgren, E.P.; Degen, J.L.; Palmiter, R.D.; Brinster, R.L. Neonatal Bleeding in Transgenic Mice Expressing Urokinase-Type Plasminogen Activator. Cell 1990, 62, 447–456. [Google Scholar] [CrossRef]
- Dandri, M.; Burda, M.R.; Török, E.; Pollok, J.M.; Iwanska, A.; Sommer, G.; Rogiers, X.; Rogler, C.E.; Gupta, S.; Will, H.; et al. Repopulation of Mouse Liver with Human Hepatocytes and in Vivo Infection with Hepatitis B Virus. Hepatology 2001, 33, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Mercer, D.F.; Schiller, D.E.; Elliott, J.F.; Douglas, D.N.; Hao, C.; Rinfret, A.; Addison, W.R.; Fischer, K.P.; Churchill, T.A.; Lakey, J.R.; et al. Hepatitis C Virus Replication in Mice with Chimeric Human Livers. Nat. Med. 2001, 7, 927–933. [Google Scholar] [CrossRef]
- Meuleman, P.; Libbrecht, L.; De Vos, R.; de Hemptinne, B.; Gevaert, K.; Vandekerckhove, J.; Roskams, T.; Leroux-Roels, G. Morphological and Biochemical Characterization of a Human Liver in a UPA-SCID Mouse Chimera. Hepatology 2005, 41, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, A.; Stift, J.; Maric, D.; Cui, Q.; Dienes, H.-P.; Feinstone, S.M. Chimeric Mouse Model for the Infection of Hepatitis B and C Viruses. PLoS ONE 2013, 8, e77298. [Google Scholar] [CrossRef]
- Lütgehetmann, M.; Mancke, L.V.; Volz, T.; Helbig, M.; Allweiss, L.; Bornscheuer, T.; Pollok, J.M.; Lohse, A.W.; Petersen, J.; Urban, S.; et al. Humanized Chimeric UPA Mouse Model for the Study of Hepatitis B and D Virus Interactions and Preclinical Drug Evaluation. Hepatology 2012, 55, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Giersch, K.; Helbig, M.; Volz, T.; Allweiss, L.; Mancke, L.V.; Lohse, A.W.; Polywka, S.; Pollok, J.M.; Petersen, J.; Taylor, J.; et al. Persistent Hepatitis D Virus Mono-Infection in Humanized Mice Is Efficiently Converted by Hepatitis B Virus to a Productive Co-Infection. J. Hepatol. 2014, 60, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Giersch, K.; Bhadra, O.D.; Volz, T.; Allweiss, L.; Riecken, K.; Fehse, B.; Lohse, A.W.; Petersen, J.; Sureau, C.; Urban, S.; et al. Hepatitis Delta Virus Persists during Liver Regeneration and Is Amplified through Cell Division Both in Vitro and in Vivo. Gut 2019, 68, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Giersch, K.; Allweiss, L.; Volz, T.; Helbig, M.; Bierwolf, J.; Lohse, A.W.; Pollok, J.M.; Petersen, J.; Dandri, M.; Lütgehetmann, M. Hepatitis Delta Co-Infection in Humanized Mice Leads to Pronounced Induction of Innate Immune Responses in Comparison to HBV Mono-Infection. J. Hepatol. 2015, 63, 346–353. [Google Scholar] [CrossRef]
- Giersch, K.; Homs, M.; Volz, T.; Helbig, M.; Allweiss, L.; Lohse, A.W.; Petersen, J.; Buti, M.; Pollicino, T.; Sureau, C.; et al. Both Interferon Alpha and Lambda Can Reduce All Intrahepatic HDV Infection Markers in HBV/HDV Infected Humanized Mice. Sci. Rep. 2017, 7, 3757. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Tateno, C.; Thi, E.P.; Kakuni, M.; Snead, N.M.; Ishida, Y.; Barnard, T.R.; Sofia, M.J.; Shimada, T.; Lee, A.C.H. Hepatitis B Virus Therapeutic Agent ARB-1740 Has Inhibitory Effect on Hepatitis Delta Virus in a New Dually-Infected Humanized Mouse Model. ACS Infect. Dis. 2019, 5, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, G.; Ruppert, S.; Schedl, A.; Schmid, E.; Thies, E.; Schütz, G. Multiple Effects on Liver-Specific Gene Expression in Albino Lethal Mice Caused by Deficiency of an Enzyme in Tyrosine Metabolism. J. Cell Sci. Suppl. 1992, 16, 117–122. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Zhang, H.; Zhang, X.; Xie, D.; Chen, Y.; Wangensteen, K.J.; Ekker, S.C.; Firpo, M.; Liu, C.; Xiang, D.; et al. Liver Xeno-Repopulation with Human Hepatocytes in Fah-/-Rag2-/- Mice after Pharmacological Immunosuppression. Am. J. Pathol. 2010, 177, 1311–1319. [Google Scholar] [CrossRef]
- Burwitz, B.J.; Zhou, Z.; Li, W. Animal Models for the Study of Human Hepatitis B and D Virus Infection: New Insights and Progress. Antivir. Res. 2020, 182, 104898. [Google Scholar] [CrossRef]
- Washburn, M.L.; Bility, M.T.; Zhang, L.; Kovalev, G.I.; Buntzman, A.; Frelinger, J.A.; Barry, W.; Ploss, A.; Rice, C.M.; Su, L. A Humanized Mouse Model to Study Hepatitis C Virus Infection, Immune Response, and Liver Disease. Gastroenterology 2011, 140, 1334–1344. [Google Scholar] [CrossRef] [Green Version]
- Bility, M.T.; Cheng, L.; Zhang, Z.; Luan, Y.; Li, F.; Chi, L.; Zhang, L.; Tu, Z.; Gao, Y.; Fu, Y.; et al. Hepatitis B Virus Infection and Immunopathogenesis in a Humanized Mouse Model: Induction of Human-Specific Liver Fibrosis and M2-like Macrophages. PLoS Pathog. 2014, 10, e1004032. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Jiang, J.; Liu, X.; Zhang, Y.; Zhang, L.; Xin, J.; Wu, K.; Li, X.; Cao, J.; Guo, X.; et al. HBV Infection-Induced Liver Cirrhosis Development in Dual-Humanised Mice with Human Bone Mesenchymal Stem Cell Transplantation. Gut 2019, 68, 2044–2056. [Google Scholar] [CrossRef] [Green Version]
- Strick-Marchand, H.; Dusséaux, M.; Darche, S.; Huntington, N.D.; Legrand, N.; Masse-Ranson, G.; Corcuff, E.; Ahodantin, J.; Weijer, K.; Spits, H.; et al. A Novel Mouse Model for Stable Engraftment of a Human Immune System and Human Hepatocytes. PLoS ONE 2015, 10, e0119820. [Google Scholar] [CrossRef]
- Kremsdorf, D.; Strick-Marchand, H. Modeling Hepatitis Virus Infections and Treatment Strategies in Humanized Mice. Curr. Opin. Virol. 2017, 25, 119–125. [Google Scholar] [CrossRef]
- Tham, C.Y.L.; Kah, J.; Tan, A.T.; Volz, T.; Chia, A.; Giersch, K.; Ladiges, Y.; Loglio, A.; Borghi, M.; Sureau, C.; et al. Hepatitis Delta Virus Acts as an Immunogenic Adjuvant in Hepatitis B Virus-Infected Hepatocytes. Cell Rep. Med. 2020, 1, 100060. [Google Scholar] [CrossRef]
- Suárez-Amarán, L.; Usai, C.; Di Scala, M.; Godoy, C.; Ni, Y.; Hommel, M.; Palomo, L.; Segura, V.; Olagüe, C.; Vales, A.; et al. A New HDV Mouse Model Identifies Mitochondrial Antiviral Signaling Protein (MAVS) as a Key Player in IFN-β Induction. J. Hepatol. 2017, 67, 669–679. [Google Scholar] [CrossRef]
- Usai, C.; Maestro, S.; Camps, G.; Olague, C.; Suárez-Amaran, L.; Vales, A.; Aragon, T.; Hommel, M.; Aldabe, R.; Gonzalez-Aseguinolaza, G. TNF-Alpha Inhibition Ameliorates HDV-Induced Liver Damage in a Mouse Model of Acute Severe Infection. JHEP Rep. 2020, 2, 100098. [Google Scholar] [CrossRef] [Green Version]
- Giersch, K.; Hermanussen, L.; Volz, T.; Kah, J.; Allweiss, L.; Casey, J.; Sureau, C.; Dandri, M.; Lütgehetmann, M. Murine Hepatocytes Do Not Support Persistence of Hepatitis D Virus Mono-Infection in Vivo. Liver Int. 2021, 41, 410–419. [Google Scholar] [CrossRef]
- Meier, A.; Mehrle, S.; Weiss, T.S.; Mier, W.; Urban, S. Myristoylated PreS1-Domain of the Hepatitis B Virus L-Protein Mediates Specific Binding to Differentiated Hepatocytes. Hepatology 2013, 58, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Schieck, A.; Schulze, A.; Gähler, C.; Müller, T.; Haberkorn, U.; Alexandrov, A.; Urban, S.; Mier, W. Hepatitis B Virus Hepatotropism is Mediated by Specific Receptor Recognition in the Liver and Not Restricted to Susceptible Hosts. Hepatology 2013, 58, 43–53. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Ren, B.; Mao, F.; Jing, Z.; Li, Y.; Liu, Y.; Peng, B.; Yan, H.; Qi, Y.; Sun, Y.; et al. Hepatitis D Virus Infection of Mice Expressing Human Sodium Taurocholate Co-Transporting Polypeptide. PLoS Pathog. 2015, 11, e1004840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, O. Innate Immunity of the Newborn: Basic Mechanisms and Clinical Correlates. Nat. Rev. Immunol. 2007, 7, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Dakic, A.; Shao, Q.; D’Amico, A.; O’Keeffe, M.; Chen, W.; Shortman, K.; Wu, L. Development of the Dendritic Cell System during Mouse Ontogeny. J. Immunol. 2004, 172, 1018–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, D.M.P.; Vaughn, M.M.; Belman, A.R.; Cole, J.S.; Rall, G.F. Immune Response-Mediated Protection of Adult but Not Neonatal Mice from Neuron-Restricted Measles Virus Infection and Central Nervous System Disease. J. Virol. 1999, 73, 1795–1801. [Google Scholar] [CrossRef] [Green Version]
- Winer, B.Y.; Shirvani-Dastgerdi, E.; Bram, Y.; Sellau, J.; Low, B.E.; Johnson, H.; Huang, T.; Hrebikova, G.; Heller, B.; Sharon, Y.; et al. Preclinical Assessment of Antiviral Combination Therapy in a Genetically Humanized Mouse Model for Hepatitis Delta Virus Infection. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Cao, Z.; Mao, F.; Ren, B.; Li, Y.; Li, D.; Li, H.; Peng, B.; Yan, H.; Qi, Y.; et al. Modification of Three Amino Acids in Sodium Taurocholate Cotransporting Polypeptide Renders Mice Susceptible to Infection with Hepatitis D Virus In Vivo. J. Virol. 2016, 90, 8866–8874. [Google Scholar] [CrossRef] [Green Version]
- Sureau, C.; Jacob, J.R.; Eichberg, J.W.; Lanford, R.E. Tissue Culture System for Infection with Human Hepatitis Delta Virus. J. Virol. 1991, 65, 3443–3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Zhuang, Q.; Wang, Y.; Zhang, T.; Zhao, J.; Zhang, Y.; Zhang, J.; Lin, Y.; Yuan, Q.; Xia, N.; et al. HBV Life Cycle Is Restricted in Mouse Hepatocytes Expressing Human NTCP. Cell. Mol. Immunol. 2014, 11, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Guidotti, L.G.; Matzke, B.; Schaller, H.; Chisari, F.V. High-Level Hepatitis B Virus Replication in Transgenic Mice. J. Virol. 1995, 69, 6158–6169. [Google Scholar] [CrossRef] [Green Version]
- Lempp, F.A.; Mutz, P.; Lipps, C.; Wirth, D.; Bartenschlager, R.; Urban, S. Evidence That Hepatitis B Virus Replication in Mouse Cells Is Limited by the Lack of a Host Cell Dependency Factor. J. Hepatol. 2016, 64, 556–564. [Google Scholar] [CrossRef]
- Marchetti, A.L.; Guo, H. New Insights on Molecular Mechanism of Hepatitis B Virus Covalently Closed Circular DNA Formation. Cells 2020, 9, 2430. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Ploss, A. Hepatitis B Virus CccDNA is Formed through Distinct Repair Processes of Each Strand. Nat. Commun. 2021, 12, 1591. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Saso, W.; Sugiyama, R.; Ishii, K.; Ohki, M.; Nagamori, S.; Suzuki, R.; Aizaki, H.; Ryo, A.; Yun, J.-H.; et al. Epidermal Growth Factor Receptor Is a Host-Entry Cofactor Triggering Hepatitis B Virus Internalization. Proc. Natl. Acad. Sci. USA 2019, 116, 8487–8492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, J.L.; Phelps, E.D.; Doll, M.A.; Schaal, S.; Ceresa, B.P. The Role of Endogenous Epidermal Growth Factor Receptor Ligands in Mediating Corneal Epithelial Homeostasis. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2870–2880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verrier, E.R.; Colpitts, C.C.; Bach, C.; Heydmann, L.; Weiss, A.; Renaud, M.; Durand, S.C.; Habersetzer, F.; Durantel, D.; Abou-Jaoudé, G.; et al. A Targeted Functional RNA Interference Screen Uncovers Glypican 5 as an Entry Factor for Hepatitis B and D Viruses. Hepatology 2016, 63, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Verrier, E.R.; Weiss, A.; Bach, C.; Heydmann, L.; Turon-Lagot, V.; Kopp, A.; El Saghire, H.; Crouchet, E.; Pessaux, P.; Garcia, T.; et al. Combined Small Molecule and Loss-of-Function Screen Uncovers Estrogen Receptor Alpha and CAD as Host Factors for HDV Infection and Antiviral Targets. Gut 2020, 69, 158–167. [Google Scholar] [CrossRef]


{kind=link}
| Model | Pros | Cons | Ref. |
|---|---|---|---|
| Chimpanzees | Full HDV life cycle, innate and adaptive immune system, HBV infection | Highly restricted availability, ethical concerns and high costs | [15,44,45,46] |
| Woodchucks | Full HDV life cycle, innate and adaptive immune system | Co-infection only with woodchuck hepatitis virus (WHV), limited availability, high costs and difficulties in the experimental performance | [50,51,52,53,54,55,56,57] |
| Hydrodynamic mouse models | Innate and adaptive immune system | No HDV entry, HDV clearance within 30 days, liver damage due to large injection volume | [68,69] |
| Human liver chimeric mice | Full HDV life cycle, innate immune system, HBV infection, life-long HDV persistence | No adaptive immune system, limited availability, high costs | [75,81,82,83,84,85,86] |
| AAV based mouse models | Innate and adaptive immune system, liver inflammation and damage, HBV replication | No HDV entry, HDV decline after 21 days until 45 days (long-term data missing) | [96,97] |
| hNTCP transgenic mice hNTCP/BAC mice | Innate and adaptive immune system | Low HDV infection efficacy, HDV clearance within 21 days, no HBV infection, presence of mNTCP | [101,105] |
| Mice with humanized NTCP (CRISPR/Cas, TALENs) | Innate and adaptive immune system | Low HDV infection efficacy, HDV clearance within 21 days, no HBV infection | [98,106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giersch, K.; Dandri, M. In Vivo Models of HDV Infection: Is Humanizing NTCP Enough? Viruses 2021, 13, 588. https://doi.org/10.3390/v13040588
Giersch K, Dandri M. In Vivo Models of HDV Infection: Is Humanizing NTCP Enough? Viruses. 2021; 13(4):588. https://doi.org/10.3390/v13040588
Chicago/Turabian StyleGiersch, Katja, and Maura Dandri. 2021. "In Vivo Models of HDV Infection: Is Humanizing NTCP Enough?" Viruses 13, no. 4: 588. https://doi.org/10.3390/v13040588
APA StyleGiersch, K., & Dandri, M. (2021). In Vivo Models of HDV Infection: Is Humanizing NTCP Enough? Viruses, 13(4), 588. https://doi.org/10.3390/v13040588