
Probing into the Effects of Grapevine Leafroll-Associated Viruses on the Physiology, Fruit Quality and Gene Expression of Grapes

Abstract
:1. The Family Closteroviridae: A Brief Overview
2. Grapevine Leafroll Disease and GLRaVs
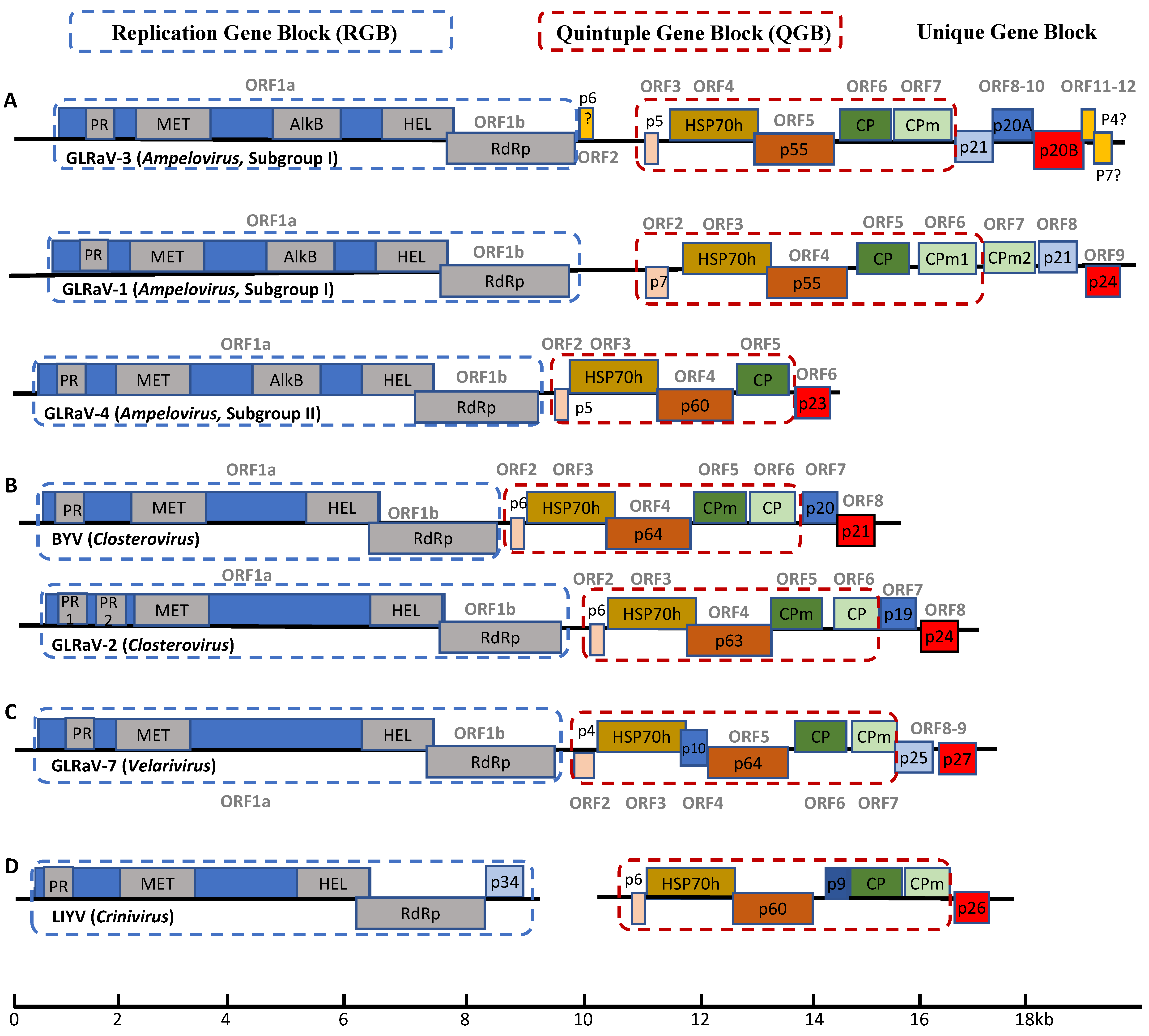
2.1. Multiple Viruses of the Genus Ampelovirus Infect Grapevine
2.2. GLRaV-2, a Member of the Genus Closterovirus
2.3. GLRaV-7, the Prototype Member of the Genus Velarivirus
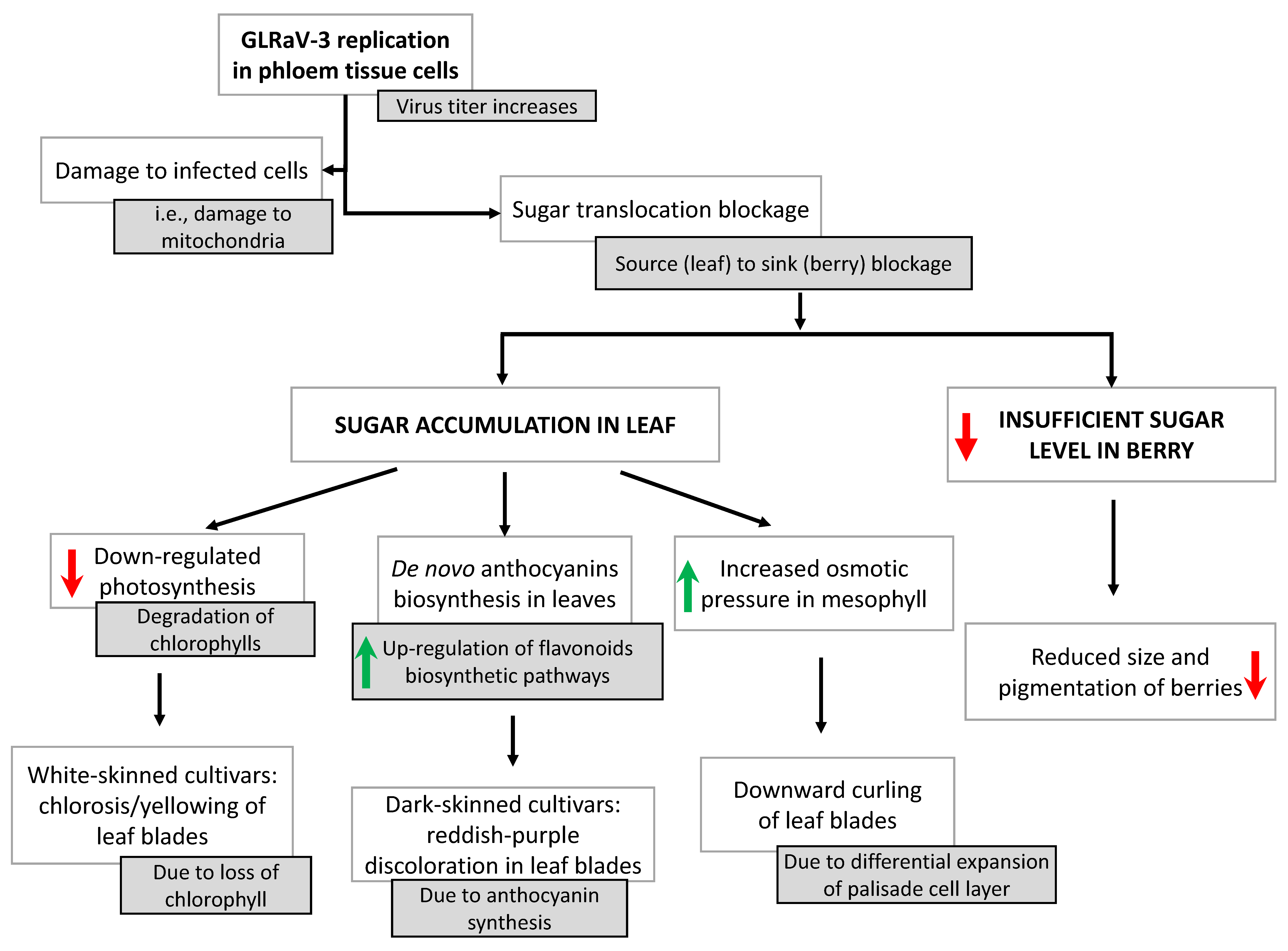
3. Effects of GLRaVs on Physiology, Fruit Quality and Gene Expression
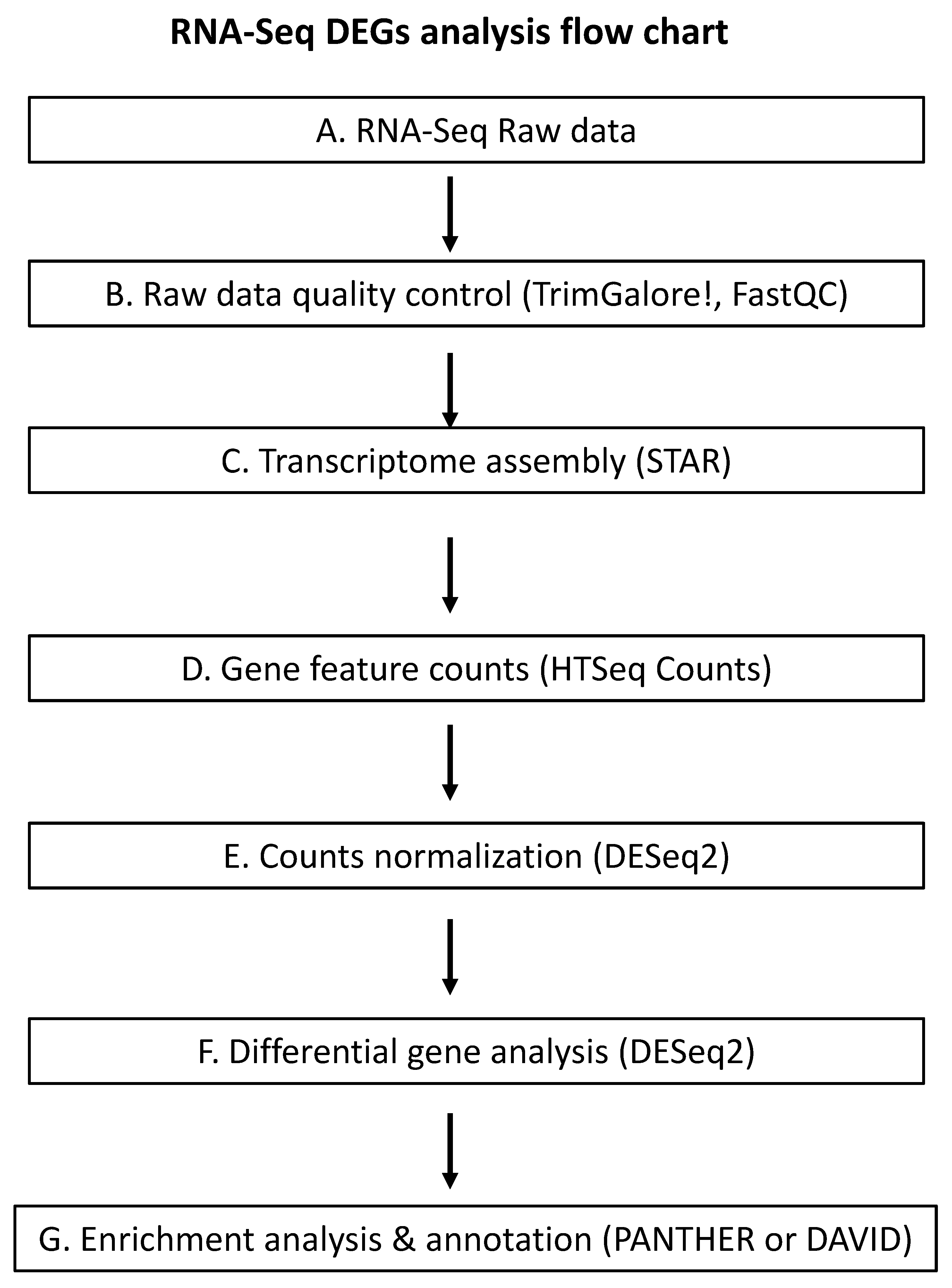
4. Methodologies to Study Effects of GLRaVs on Grapevine
5. Conclusions and Future Research
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agranovsky, A.A. Closteroviruses: Molecular Biology, Evolution and Interactions with Cells. In Plant Viruses: Evolution and Management; Springer: Singapore, 2016; pp. 231–252. [Google Scholar]
- Karasev, A.V. Genetic Diversity and Evolution of Closteroviruses. Annu. Rev. Phytopathol. 2000, 38, 293–324. [Google Scholar] [CrossRef] [PubMed]
- Candresse, T.; Fuchs, M. Closteroviridae. In eLS; John Wiley & Sons, Inc.: Chinchester, UK, 2020; pp. 1–10. [Google Scholar]
- Masters, P.S.; Perlman, S. Coronaviridae. Fields Virol. 2013, 1, 825–858. [Google Scholar]
- Dolja, V.V.; Karasev, A.V.; Koonin, E.V. Molecular Biology and Evolution of Closteroviruses: Sophisticated Build-up of Large RNA Genomes. Annu. Rev. Phytopathol. 1994, 32, 261–285. [Google Scholar] [CrossRef]
- Peremyslov, V.V.; Hagiwara, Y.; Dolja, V.V. Genes Required for Replication of the 15.5-Kilobase RNA Genome of a Plant Closterovirus. J. Virol. 1998, 72, 5870–5876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, C.-W.; Peremyslov, V.V.; Mushegian, A.R.; Dawson, W.O.; Dolja, V.V. Functional Specialization and Evolution of Leader Proteinases in the Family Closteroviridae. J. Virol. 2001, 75, 12153–12160. [Google Scholar] [CrossRef] [Green Version]
- Dolja, V.V.; Kreuze, J.F.; Valkonen, J.P.T. Comparative and functional genomics of closteroviruses. Virus Res. 2006. [Google Scholar] [CrossRef]
- Peremyslov, V.V.; Pan, Y.W.; Dolja, V.V. Movement Protein of a Closterovirus Is a Type III Integral Transmembrane Protein Localized to the EndoplasmicReticulum. J. Virol. 2004, 78, 3704–3709. [Google Scholar] [CrossRef] [Green Version]
- Alzhanova, D.V.; Hagiwara, Y.; Peremyslov, V.V.; Dolja, V.V. Genetic Analysis of the Cell-to-Cell Movement of Beet Yellows Closterovirus. Virology 2000, 268, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peremyslov, V.V.; Andreev, I.A.; Prokhnevsky, A.I.; Duncan, G.H.; Taliansky, M.E.; Dolja, V.V. Complex molecular architecture of beet yellows virus particles. Proc. Natl. Acad. Sci. USA 2004, 101, 5030–5035. [Google Scholar] [CrossRef] [Green Version]
- Dolja, V.V. Beet yellows virus: The importance of being different. Mol. Plant Pathol. 2003, 4, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Abou Ghanem-Sabanadzovic, N.; Sabanadzovic, S.; Gugerli, P.; Rowhani, A. Genome organization, serology and phylogeny of Grapevine leafroll-associated viruses 4 and 6: Taxonomic implications. Virus Res. 2012, 163, 120–128. [Google Scholar] [CrossRef]
- Kiss, Z.A.; Medina, V.; Falk, B.W. Crinivirus replication and host interactions. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Maliogka, V.I.; Wintermantel, W.M.; Orfanidou, C.G.; Katis, N.I. Criniviruses infecting vegetable crops. In Applied Plant Biotechnology for Improving Resistance to Biotic Stress; Elsevier: Amsterdam, The Netherlands, 2020; pp. 251–289. [Google Scholar]
- Stewart, L.R.; Hwang, M.S.; Falk, B.W. Two Crinivirus-specific proteins of Lettuce infectious yellows virus (LIYV), P26 and P9, are self-interacting. Virus Res. 2009, 145, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, J.; Feng, M.; Wang, X.; Luo, C.; Wang, Q.; Cheng, Y. Characterization of silencing suppressor p24 of Grapevine leafroll-associated virus 2. Mol. Plant Pathol. 2018, 19, 355–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Luo, C.; Xu, Y.; Zhang, C.; Bao, M.; Dou, J.; Wang, Q.; Cheng, Y. Expression of the p24 silencing suppressor of Grapevine leafroll-associated virus 2 from Potato virus X or Barley stripe mosaic virus vector elicits hypersensitive responses in Nicotiana benthamiana. Plant Physiol. Biochem. 2019, 142, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-W.; Liu, Q.; Zeng, Q.; Huang, W.-T.; Wang, Q.; Cheng, Y.-Q. p24G1 Encoded by Grapevine Leafroll-Associated Virus 1 Suppresses RNA Silencing and Elicits Hypersensitive Response-Like Necrosis in Nicotiana Species. Viruses 2020, 12, 1111. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Helpio, E.; Falk, B. Two Crinivirus-Conserved Small Proteins, P5 and P9, Are Indispensable for Efficient Lettuce infectious yellows virus Infectivity in Plants. Viruses 2018, 10, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, G.P.; Ghanem-sabanadzovic, N.A.; Agranovsky, A.A.; Al Rwahnih, M.; Dolja, V.V.; Dovas, C.I. Taxonomic revision of the family Closteroviridae with special reference to the Grapevine leafroll-associated members of the Genus Ampelovirus and the putative species unassigned to the family. J. Plant Pathol. 2012. [Google Scholar] [CrossRef]
- Dawson, W.O. Molecular genetics of citrus tristeza virus. In Citrus Tristeza Virus Complex and Tristeza Diseases; Karasev, A.V., Hilf, M.E., Eds.; APS Press: St. Paul, MN, USA, 2010; pp. 53–72. [Google Scholar]
- Aboughanem-Sabanadzovic, N.; Maliogka, V.; Sabanadzovic, S. Grapevine leafroll-associated virus 4. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 197–220. [Google Scholar]
- Klaassen, V.A.; Boeshore, M.L.; Koonin, E.V.; Tian, T.; Falk, B.W. Genome Structure and Phylogenetic Analysis of Lettuce Infectious Yellows Virus, a Whitefly-Transmitted, Bipartite Closterovirus. Virology 1995, 208, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.; Meng, B. Prediction of the molecular boundary and functionality of novel viral AlkB domains using homology modelling and principal component analysis. J. Gen. Virol. 2019, 100, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Van den Born, E.; Omelchenko, M.V.; Bekkelund, A.; Leihne, V.; Koonin, E.V.; Dolja, V.V.; Falnes, P.O. Viral AlkB proteins repair RNA damage by oxidative demethylation. Nucleic Acids Res. 2008, 36, 5451–5461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIV Statistical Report on World Vitiviniculture; 2019. Available online: https://www.oiv.int/public/medias/6782/oiv-2019-statistical-report-on-world-vitiviniculture.pdf (accessed on 27 November 2020).
- Organization Internationale de la Vigne et du Vin. OIV Definition of the Vitivinicultural Products by Code Sheet; Rue d’Aguesseaun: Paris, France, 2016; Available online: https://www.oiv.int/en/technical-standards-and-documents/products-definition-and-labelling/definition-of-the-vitivinicultural-products-by-code-sheet (accessed on 27 November 2020).
- Rayapati, N.; O’Neal, S.; Walsh, D. Grapevine Leafroll Disease. Washington State University Extension Bulletin EB2027E; WSU Press: Pullman, WA, USA, April 2008. [Google Scholar]
- Almeida, R.P.P.; Daane, K.M.; Bell, V.A.; Blaisdell, G.K.; Cooper, M.L.; Herrbach, E.; Pietersen, G. Ecology and management of grapevine leafroll disease. Front. Microbiol. 2013, 4, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naidu, R.A. Grapevine leafroll-associated virus 1. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Golino, D.A., Martelli, G.P., Fuchs, M.F., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 127–140. [Google Scholar]
- Tsai, C.-W.; Rowhani, A.; Golino, D.A.; Daane, K.M.; Almeida, R.P.P. Mealybug transmission of grapevine leafroll viruses: An analysis of virus–vector specificity. Phytopathology 2010, 100, 830–834. [Google Scholar] [CrossRef] [Green Version]
- Herrbach, E.; Alliaume, A.; Prator, C.A.; Daane, K.M.; Cooper, M.L.; Almeida, R.P.P. Vector transmission of trapevine leafroll-associated viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer International Publishing: Cham, Switzerland, 2017; pp. 483–503. [Google Scholar]
- Naidu, R.; Rowhani, A.; Fuchs, M.; Golino, D.; Martelli, G.P. Grapevine leafroll: A complex viral disease affecting a high-value fruit crop. Plant Dis. 2014, 98, 1172–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atallah, S.S.; Gomez, M.I.; Fuchs, M.F.; Martinson, T.E. Economic impact of grapevine leafroll disease on Vitis vinifera cv. Cabernet franc in finger lakes vineyards of New York. Am. J. Enol. Vitic. 2012, 63, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Shabanian, M.; Xiao, H.; Meng, B. Seasonal dynamics and tissue distribution of two major viruses associated with grapevine Leafroll under cool climate condition. Eur. J. Plant Pathol. 2020, 158, 1017–1031. [Google Scholar] [CrossRef]
- Donda, B.P.; Jarugula, S.; Naidu, R.A. An Analysis of the Complete Genome Sequence and Subgenomic RNAs Reveals Unique Features of the Ampelovirus, Grapevine leafroll-associated virus 1. Phytopathology 2017, 107, 1069–1079. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Holmes, E.C. Gene Duplication Is Infrequent in the Recent Evolutionary History of RNA Viruses. Mol. Biol. Evol. 2013, 30, 1263–1269. [Google Scholar] [CrossRef] [Green Version]
- Little, A.; Fazeli, C.F.; Rezaian, M.A. Hypervariable genes in Grapevine leafroll associated virus 1. Virus Res. 2001, 80, 109–116. [Google Scholar] [CrossRef]
- Alabi, O.J.; Al Rwahnih, M.; Karthikeyan, G.; Poojari, S.; Fuchs, M.; Rowhani, A.; Naidu, R.A. Grapevine leafroll-associated virus 1 Occurs as Genetically Diverse Populations. Phytopathology® 2011, 101, 1446–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteves, F.; Teixeira Santos, M.; Eiras-Dias, J.E.; Fonseca, F. Molecular data mining to improve antibody-based detection of Grapevine leafroll-associated virus 1 (GLRaV-1). J. Virol. Methods 2013, 194, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, P.M.; Wang, M.-B.; Lough, T. Gene silencing as an adaptive defence against viruses. Nature 2001, 411, 834–842. [Google Scholar] [CrossRef]
- Wang, M.-B.; Masuta, C.; Smith, N.A.; Shimura, H. RNA Silencing and Plant Viral Diseases. Mol. Plant-Microbe Interact. 2012, 25, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- García, J.A.; Pallás, V. Viral factors involved in plant pathogenesis. Curr. Opin. Virol. 2015, 11, 21–30. [Google Scholar] [CrossRef]
- Martelli, G.P. Directory of virus and virus-like diseases of the grapevine and their agents. J. Plant Pathol. 2014, 96, 1–136. [Google Scholar] [CrossRef]
- Ricketts, K.D.; Gomez, M.I.; Atallah, S.S.; Fuchs, M.F.; Martinson, T.E.; Battany, M.C.; Bettiga, L.J.; Cooper, M.L.; Verdegaal, P.S.; Smith, R.J. Reducing the economic impact of grapevine leafroll disease in california: Identifying optimal disease management strategies. Am. J. Enol. Vitic. 2015, 66, 138–149. [Google Scholar] [CrossRef]
- Burger, J.T.; Maree, H.J.; Gouveia, P.; Naidu, R.A. Grapevine leafroll-associated virus 3. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 167–195. [Google Scholar]
- Maree, H.J.; Almeida, R.P.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.C.; Martelli, G.P.; et al. Grapevine leafroll-associated virus 3. Front. Microbiol. 2013, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Bester, R.; Maree, H.J.; Burger, J.T. Complete nucleotide sequence of a new strain of grapevine leafroll-associated virus 3 in South Africa. Arch. Virol. 2012, 157, 1815–1819. [Google Scholar] [CrossRef]
- Seah, Y.M.; Sharma, A.M.; Zhang, S.; Almeida, R.P.P.; Duffy, S. A divergent variant of grapevine leafroll-associated virus 3 is present in California. Virol. J. 2012, 9, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maree, H.J.; Pirie, M.D.; Oosthuizen, K.; Bester, R.; Jasper, D.; Rees, G.; Burger, J.T. Phylogenomic analysis reveals deep divergence and recombination in an economically important grapevine virus. PLoS ONE 2015, 10, e0126819. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.D.; Dahan, J.; Lee, J.; Martin, R.R.; Karasev, A.V. A Novel Genetic Variant of Grapevine leafroll-associated virus-3 (GLRaV-3) from Idaho Grapevines. Plant Dis. 2019, 103, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chooi, K.M.; Cohen, D.; Pearson, M.N. Molecular characterisation of two divergent variants of grapevine leafroll-associated virus 3 in New Zealand. Arch. Virol. 2013, 158, 1597–1602. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, P.; Dandlen, S.; Costa, Â.; Marques, N.; Nolasco, G. Identification of an RNA silencing suppressor encoded by Grapevine leafroll-associated virus 3. Eur. J. Plant Pathol. 2012, 133, 237–245. [Google Scholar] [CrossRef]
- Jarugula, S.; Gowda, S.; Dawson, W.O.; Naidu, R.A. 3’-coterminal subgenomic RNAs and putative cis-acting elements of Grapevine leafroll-associated virus 3 reveals “unique” features of gene expression strategy in the genus Ampelovirus. Virol. J. 2010, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Navas-Castillo, J.; Albiach-Martí, M.R.; Gowda, S.; Hilf, M.E.; Garnsey, S.M.; Dawson, W.O. Kinetics of accumulation of citrus tristeza virus RNAs. Virology 1997, 228, 92–97. [Google Scholar] [CrossRef]
- Hagiwara, Y.; Peremyslov, V.V.; Dolja, V.V. Regulation of Closterovirus Gene Expression Examined by Insertion of a Self-Processing Reporter and by Northern Hybridization. J. Virol. 1999, 73, 7988–7993. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Nakaune, R. Molecular characterization of a novel putative ampelovirus tentatively named grapevine leafroll-associated virus 13. Arch. Virol. 2016, 161, 2555–2559. [Google Scholar] [CrossRef] [PubMed]
- Abou Ghanem-Sabanadzovic, N.; Sabanadzovic, S.; Uyemoto, J.K.; Golino, D.; Rowhani, A. A putative new ampelovirus associated with grapevine leafroll disease. Arch. Virol. 2010, 155, 1871–1876. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.S.; Gonsalves, D.; Teliz, D. Characterization of closterovirus-like particles associated with grapevine leafroll disease. J. Phytopathol. 1990, 128, 1–14. [Google Scholar] [CrossRef]
- Zimmermann, D.; Bass, P.; Legin, R.; Walter, B. Characterization and Serological Detection of Four Closterovirus-like Particles Associated with Leafroll Disease on Grapevine. J. Phytopathol. 1990, 130, 205–218. [Google Scholar] [CrossRef]
- Gugerli, P.; Brugger, J.J.; Ramel, M.E. Identification immuno- chimique du 6e virus associé à la maladie de l’enroulement de la vigne et amélioration des techniques de diagnostic pour la sélection sanitaire en viticulture. Rev. Suisse Vitic. Arboric. Hortic. 1997, 29, 137–141. [Google Scholar]
- Alkowni, R.; Rowhani, A.; Daubert, S.; Golino, D. Partial characterization of a new ampelovirus associated with grapevine leafroll disease. J. Plant Pathol. 2004, 86, 123–133. [Google Scholar]
- Maliogka, V.I.; Dovas, C.I.; Lotos, L.; Efthimiou, K.; Katis, N.I. Complete genome analysis and immunodetection of a member of a novel virus species belonging to the genus Ampelovirus. Arch. Virol. 2009, 154, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Nakaune, R.; Nakano, M.; Suzaki, K. Novel variants of grapevine leafroll-associated virus 4 and 7 detected from a grapevine showing leafroll symptoms. Arch. Virol. 2013, 158, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Reynard, J.-S.; Schneeberger, P.H.H.; Frey, J.E.; Schaerer, S. Biological, Serological, and Molecular Characterization of a Highly Divergent Strain of Grapevine leafroll-associated virus 4 Causing Grapevine Leafroll Disease. Phytopathology® 2015, 105, 1262–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, B.; Li, C.; Goszczynski, D.E.; Gonsalves, D. Genome Sequences and Structures of Two Biologically Distinct Strains of Grapevine leafroll-associated virus 2 and Sequence Analysis. Virus Genes 2005, 31, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkowni, R.; Zhang, Y.-P.; Rowhani, A.; Uyemoto, J.K.; Minafra, A. Biological, molecular, and serological studies of a novel strain of grapevine leafroll-associated virus 2. Virus Genes 2011, 43, 102–110. [Google Scholar] [CrossRef]
- Uyemoto, J.K.; Rowhani, A.; Luvisi, D.; Krag, C.R. New closterovirus in ‘Redglobe’ grape causes decline of grafted plants. Calif. Agric. 2001, 55, 28–31. [Google Scholar] [CrossRef]
- Bonfiglioli, R.; Edwards, F.; Hoskins, N.; Pantaleo, A. Graft incompatibility syndrome in New Zealand Merlot vines involves another possible variant of GLRaV-2. Aust. N. Z. Grapegrow. Winemak. 2003, 476, 50–54. [Google Scholar]
- Bertazzon, N.; Borgo, M.; Vanin, S.; Angelini, E. Genetic variability and pathological properties of grapevine leafroll-associated virus 2 isolates. Eur. J. Plant Pathol. 2010, 127, 185–197. [Google Scholar] [CrossRef]
- Poojari, S.; Alabi, O.J.; Naidu, R.A. Molecular characterization and impacts of a strain of grapevine leafroll-associated virus 2 causing asymptomatic infection in a wine grape cultivar. Virol. J. 2013, 10, 324. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.C.; Kasschau, K.D.; Prokhnevsky, A.I.; Gopinath, K.; Pogue, G.P.; Carrington, J.C.; Dolja, V.V. Suppressor of RNA silencing encoded by Beet yellows virus. Virology 2003, 306, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Folimonov, A.; Shintaku, M.; Li, W.-X.; Falk, B.W.; Dawson, W.O.; Ding, S.-W. Three distinct suppressors of RNA silencing encoded by a 20-kb viral RNA genome. Proc. Natl. Acad. Sci. USA 2004, 101, 15742–15747. [Google Scholar] [CrossRef] [Green Version]
- Chiba, M.; Reed, J.C.; Prokhnevsky, A.I.; Chapman, E.J.; Mawassi, M.; Koonin, E.V.; Carrington, J.C.; Dolja, V.V. Diverse suppressors of RNA silencing enhance agroinfection by a viral replicon. Virology 2006, 346, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Ling, K.S.; McFerson, J.R.; Zhu, H.Y.; Goszczynski, D.E.; Gonsalves, D. Nucleotide sequence and genome organization of grapevine leafroll-associated virus-2 are similar to beet yellows virus, the closterovirus type member. J. Gen. Virol. 1998, 79, 1289–1298. [Google Scholar] [CrossRef] [Green Version]
- Angelini, E.; Aboughanem-Sabanadzovic, N.; Dolja, V.V.; Meng, B. Grapevine leafroll-associated virus 2. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D., Fuchs, M., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 141–165. [Google Scholar]
- Prokhnevsky, A.I.; Peremyslov, V.V.; Napuli, A.J.; Dolja, V.V. Interaction between Long-Distance Transport Factor and Hsp70-Related Movement Protein of Beet Yellows Virus. J. Virol. 2002, 76, 11003–11011. [Google Scholar] [CrossRef] [Green Version]
- Rwahnih, M.A.; Dolja, V.V.; Daubert, S.; Koonin, E.V.; Rowhani, A. Genomic and biological analysis of grapevine leafroll-associated virus 7 reveals a possible new genus within the family Closteroviridae. Virus Res. 2012, 163, 302–309. [Google Scholar] [CrossRef]
- Jelkmann, W.; Mikona, C.; Turturo, C.; Navarro, B.; Rott, M.E.; Menzel, W.; Saldarelli, P.; Minafra, A.; Martelli, G.P. Molecular characterization and taxonomy of grapevine leafroll-associated virus 7. Arch. Virol. 2012, 157, 359–362. [Google Scholar] [CrossRef]
- Akbaş, B.; Kunter, B.; Ilhan, D. Occurrence and distribution of grapevine leafroll-associated viruses 1, 2, 3 and 7 in Turkey. J. Phytopathol. 2007. [Google Scholar] [CrossRef]
- Lyu, M.D.; Li, M.J.; Li, J.; Li, X.M.; Cheng, Y.-Q. First Report of Grapevine leafroll-associated virus 7 in Two Native Grape Varieties in China. Plant Dis. 2013, 97, 150. [Google Scholar] [CrossRef]
- Engel, E.A.; Escobar, P.; Montt, C.; Gómez-Talquenca, S.; Valenzuela, P.D.T. First Report on the Occurrence of Grapevine leafroll-associated virus 7 and 9 in Chilean Grapevines. Plant Dis. 2008, 92, 1252. [Google Scholar] [CrossRef]
- Morales, R.Z.; Monis, J. First Detection of Grapevine leafroll associated virus-7 in California Vineyards. Plant Dis. 2007, 91, 465. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Saldarelli, P.; Rowhani, A. Grapevine leafroll-associated virus 7. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D., Fuchs, M., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 221–228. [Google Scholar]
- Avgelis, A.; Boscia, D. Grapevine leafroll-associated closterovirus 7 in Greece. Phytopathol. Mediterr. 2001, 40, 289–292. [Google Scholar] [CrossRef]
- Giribaldi, M.; Purrotti, M.; Pacifico, D.; Santini, D.; Mannini, F.; Caciagli, P.; Rolle, L.; Cavallarin, L.; Giuffrida, M.G.; Marzachì, C. A multidisciplinary study on the effects of phloem-limited viruses on the agronomical performance and berry quality of Vitis vinifera cv. Nebbiolo. J. Proteom. 2011, 75, 306–315. [Google Scholar] [CrossRef]
- Santini, D.; Rolle, L.; Cascio, P.; Mannini, F. Modifications in Chemical, Physical and Mechanical Properties of Nebbiolo (Vitis vinifera L.) Grape Berries Induced by Mixed Virus Infection. S. Afr. J. Enol. Vitic. 2011, 32. [Google Scholar] [CrossRef] [Green Version]
- Guidoni, S.; Mannini, F.; Ferrandino, A.; Argamante, N.; Di Stefano, R. Efffect of Virus Status on Leaf and Berry Phenolic Compounds in Two Wine Grapevine Vitis Vinifera Cultivars. Acta Hortic. 2000, 445–452. [Google Scholar] [CrossRef]
- Ghaffari, S.; Reynard, J.S.; Rienth, M. Single berry reconstitution prior to RNA-sequencing reveals novel insights into transcriptomic remodeling by leafroll virus infections in grapevines. Sci. Rep. 2020, 10, 12905. [Google Scholar] [CrossRef]
- Carmona, M.J.; Chaib, J.; Martinez-Zapater, J.M.; Thomas, M.R. A molecular genetic perspective of reproductive development in grapevine. J. Exp. Bot. 2008, 59, 2579–2596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-Microbe Interactions: Shaping the Evolution of the Plant Immune Response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Naidu, R.A.; Maree, H.J.; Burger, J.T. Grapevine Leafroll Disease and Associated Viruses: A Unique Pathosystem. Annu. Rev. Phytopathol. 2015, 53, 613–634. [Google Scholar] [CrossRef]
- Kovacs, L.G.; Hanami, H.; Fortenberry, M.; Kaps, M.L. Latent Infection by Leafroll Agent GLRaV-3 Is Linked to Lower Fruit Quality in French-American Hybrid Grapevines Vidal blanc and St. Vincent. Am. J. Enol. Vitic. 2001, 52, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Hale, C.R.; Woodham, R.C. Effect of Grapevine Leafroll Disease on the Acid and Potassium Composition of Sultana Grapes. Am. J. Enol. Vitic. 1979, 30, 91–92. [Google Scholar]
- Endeshaw, S.T.; Sabbatini, P.; Romanazzi, G.; Schilder, A.C.; Neri, D. Effects of grapevine leafroll associated virus 3 infection on growth, leaf gas exchange, yield and basic fruit chemistry of Vitis vinifera L. cv. Cabernet Franc. Sci. Hortic. 2014, 170, 228–236. [Google Scholar] [CrossRef]
- Singh Brar, H.; Singh, Z.; Swinny, E.; Cameron, I. Girdling and grapevine leafroll associated viruses affect berry weight, colour development and accumulation of anthocyanins in ‘Crimson Seedless’ grapes during maturation and ripening. Plant Sci. 2008, 175, 885–897. [Google Scholar] [CrossRef]
- Lee, J.; Martin, R.R. Influence of grapevine leafroll associated viruses (GLRaV-2 and -3) on the fruit composition of Oregon Vitis vinifera L. cv. Pinot noir: Phenolics. Food Chem. 2009, 112, 889–896. [Google Scholar] [CrossRef]
- Wolpert, J.A.; Vilas, E.P. Effect of Mild Leafroll Disease on Growth, Yield, and Fruit Maturity Indices of Riesling and Zinfandel. Am. J. Enol. Vitic. 1992, 43, 367–369. [Google Scholar]
- Cabaleiro, C.; Segura, A.; Garcia-Berrios, J.J. Effects of grapevine leafroll-associated virus 3 on the physiology and must of Vitis vinifera L. cv. Albariño following contamination in the field. Am. J. Enol. Vitic. 1999, 50, 40–44. [Google Scholar]
- Montero, R.; Mundy, D.; Albright, A.; Grose, C.; Trought, M.C.T.; Cohen, D.; Chooi, K.M.; MacDiarmid, R.; Flexas, J.; Bota, J. Effects of Grapevine Leafroll associated Virus 3 (GLRaV-3) and duration of infection on fruit composition and wine chemical profile of Vitis vinifera L. cv. Sauvignon blanc. Food Chem. 2016, 197, 1177–1183. [Google Scholar] [CrossRef]
- Mannini, F.; Mollo, A.; Credi, R. Field Performance and Wine Quality Modification in a Clone of Vitis vinifera cv. Dolcetto after GLRaV-3 Elimination. Am. J. Enol. Vitic. 2012, 63, 144–147. [Google Scholar] [CrossRef] [Green Version]
- Alabi, O.J.; Casassa, L.F.; Gutha, L.R.; Larsen, R.C.; Henick-Kling, T.; Harbertson, J.F.; Naidu, R.A. Impacts of grapevine leafroll disease on fruit yield and grape and wine chemistry in a wine grape (Vitis vinifera L.) cultivar. PLoS ONE 2016, 11, e0149666. [Google Scholar] [CrossRef] [Green Version]
- Christov, I.; Stefanov, D.; Velinov, T.; Goltsev, V.; Georgieva, K.; Abracheva, P.; Genova, Y.; Christov, N. The symptomless leaf infection with grapevine leafroll associated virus 3 in grown in vitro plants as a simple model system for investigation of viral effects on photosynthesis. J. Plant Physiol. 2007, 164, 1124–1133. [Google Scholar] [CrossRef]
- Bertamini, M.; Muthuchelian, K.; Nedunchezhian, N. Effect of grapevine leafroll on the photosynthesis of field grown grapevine plants (Vitis vinifera L. cv. Lagrein). J. Phytopathol. 2004, 152, 145–152. [Google Scholar] [CrossRef]
- Moutinho-Pereira, J.; Correia, C.M.; Gonçalves, B.; Bacelar, E.A.; Coutinho, J.F.; Ferreira, H.F.; Lousada, J.L.; Cortez, M.I. Impacts of leafroll-associated viruses (GLRaV-1 and -3) on the physiology of the Portuguese grapevine cultivar ‘Touriga Nacional’ growing under field conditions. Ann. Appl. Biol. 2012, 160, 237–249. [Google Scholar] [CrossRef]
- El Aou-ouad, H.; Montero, R.; Medrano, H.; Bota, J. Interactive effects of grapevine leafroll-associated virus 3 (GLRaV-3) and water stress on the physiology of Vitis vinifera L. cv. Malvasia de Banyalbufar and Giro-Ros. J. Plant Physiol. 2016, 196–197, 106–115. [Google Scholar] [CrossRef]
- Montero, R.; El aou ouad, H.; Pacifico, D.; Marzachì, C.; Castillo, N.; García, E.; Del Saz, N.F.; Florez-Sarasa, I.; Flexas, J.; Bota, J. Effects of grapevine leafroll-associated virus 3 on the physiology in asymptomatic plants of Vitis vinifera. Ann. Appl. Biol. 2017, 171, 155–171. [Google Scholar] [CrossRef]
- Halldorson, M.M.; Keller, M. Grapevine leafroll disease alters leaf physiology but has little effect on plant cold hardiness. Planta 2018, 248, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.-H.; Bi, W.-L.; Hao, X.-Y.; Xu, Y.; Li, P.-M.; Walker, M.A.; Wang, Q.-C. Responses of In vitro-Grown Plantlets (Vitis vinifera) to Grapevine leafroll-Associated Virus-3 and PEG-Induced Drought Stress. Front. Physiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, A.; Gutiérrez, R.A.; Peña-Neira, A.; Cramer, G.R.; Arce-Johnson, P. Compatible GLRaV-3 viral infections affect berry ripening decreasing sugar accumulation and anthocyanin biosynthesis in Vitis vinifera. Plant Mol. Biol. 2011, 77, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Gutha, L.R.; Casassa, L.F.; Harbertson, J.F.; Naidu, R.A. Modulation of flavonoid biosynthetic pathway genes and anthocyanins due to virus infection in grapevine (Vitis vinifera L.) leaves. BMC Plant Biol. 2010, 10, 187. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, C.; Vega, A.; Medina, C.; Schlauch, K.; Cramer, G.; Arce-Johnson, P. Gene expression associated with compatible viral diseases in grapevine cultivars. Funct. Integr. Genom. 2007, 7, 95–110. [Google Scholar] [CrossRef]
- Espinoza, C.; Medina, C.; Somerville, S.; Arce-Johnson, P. Senescence-associated genes induced during compatible viral interactions with grapevine and Arabidopsis. J. Exp. Bot. 2007, 58, 3197–3212. [Google Scholar] [CrossRef]
- Alabi, O.J.; Zheng, Y.; Jagadeeswaran, G.; Sunkar, R.; Naidu, R.A. High-throughput sequence analysis of small RNAs in grapevine (Vitis vinifera L.) affected by grapevine leafroll disease. Mol. Plant Pathol. 2012, 13, 1060–1076. [Google Scholar] [CrossRef]
- Bester, R.; Burger, J.T.; Maree, H.J. Differential expression of miRNAs and associated gene targets in grapevine leafroll-associated virus 3-infected plants. Arch. Virol. 2017, 162, 987–996. [Google Scholar] [CrossRef]
- Coito, J.L.; Rocheta, M.; Carvalho, L.; Amâncio, S. Microarray-based uncovering reference genes for quantitative real time PCR in grapevine under abiotic stress. BMC Res. Notes 2012, 5, 220. [Google Scholar] [CrossRef] [Green Version]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development. BMC Plant Biol. 2006, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Selim, M.; Legay, S.; Berkelmann-Löhnertz, B.; Langen, G.; Kogel, K.-H.; Evers, D. Identification of suitable reference genes for real-time RT-PCR normalization in the grapevine-downy mildew pathosystem. Plant Cell Rep. 2012, 31, 205–216. [Google Scholar] [CrossRef]
- Borges, A.F.; Fonseca, C.; Ferreira, R.B.; Lourenço, A.M.; Monteiro, S. Reference Gene Validation for Quantitative RT-PCR during Biotic and Abiotic Stresses in Vitis vinifera. PLoS ONE 2014, 9, e111399. [Google Scholar] [CrossRef]
- Gamm, M.; Héloir, M.-C.; Kelloniemi, J.; Poinssot, B.; Wendehenne, D.; Adrian, M. Identification of reference genes suitable for qRT-PCR in grapevine and application for the study of the expression of genes involved in pterostilbene synthesis. Mol. Genet. Genom. 2011, 285, 273–285. [Google Scholar] [CrossRef]
- González-Agüero, M.; García-Rojas, M.; Di Genova, A.; Correa, J.; Maass, A.; Orellana, A.; Hinrichsen, P. Identification of two putative reference genes from grapevine suitable for gene expression analysis in berry and related tissues derived from RNA-Seq data. BMC Genom. 2013, 14, 878. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.O.; Botstein, D. Exploring the new world of the genome with DNA microarrays. Nat. Genet. 1999, 21, 33–37. [Google Scholar] [CrossRef]
- Stefano, G.B. Comparing Bioinformatic Gene Expression Profiling Methods: Microarray and RNA-Seq. Med. Sci. Monit. Basic Res. 2014, 20, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Ulate, B.; Hopfer, H.; Figueroa-Balderas, R.; Ye, Z.; Rivero, R.M.; Albacete, A.; Pérez-Alfocea, F.; Koyama, R.; Anderson, M.M.; Smith, R.J.; et al. Red blotch disease alters grape berry development and metabolism by interfering with the transcriptional and hormonal regulation of ripening. J. Exp. Bot. 2017, 68, 1225–1238. [Google Scholar] [CrossRef] [Green Version]
- Marguerat, S.; Bähler, J. RNA-seq: From technology to biology. Cell. Mol. Life Sci. 2010, 67, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.R.; Baccarella, A.; Parrish, J.Z.; Kim, C.C. Trimming of sequence reads alters RNA-Seq gene expression estimates. BMC Bioinform. 2016, 17, 103. [Google Scholar] [CrossRef] [Green Version]
- Krueger, F. Trim Galore! Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 22 January 2021).
- Andrews, S. FastQC. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 22 January 2020).
- Kukurba, K.R.; Montgomery, S.B. RNA Sequencing and Analysis. Cold Spring Harb. Protoc. 2015, 2015, pdb.top084970. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B. Aligning Short Sequencing Reads with Bowtie. Curr. Protoc. Bioinform. 2010, 32. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Weeda, S.; Zhang, N.; Zhao, X.; Ndip, G.; Guo, Y.; Buck, G.A.; Fu, C.; Ren, S. Arabidopsis Transcriptome Analysis Reveals Key Roles of Melatonin in Plant Defense Systems. PLoS ONE 2014, 9, e93462. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, Y.; Oono, Y.; Kanamori, H.; Matsumoto, T.; Itoh, T.; Minami, E. Simultaneous RNA-Seq Analysis of a Mixed Transcriptome of Rice and Blast Fungus Interaction. PLoS ONE 2012, 7, e49423. [Google Scholar] [CrossRef] [Green Version]
- Phule, A.S.; Barbadikar, K.M.; Maganti, S.M.; Seguttuvel, P.; Subrahmanyam, D.; Babu, M.B.B.P.; Kumar, P.A. RNA-seq reveals the involvement of key genes for aerobic adaptation in rice. Sci. Rep. 2019, 9, 5235. [Google Scholar] [CrossRef] [Green Version]
- Lukoszek, R.; Feist, P.; Ignatova, Z. Insights into the adaptive response of Arabidopsis thaliana to prolonged thermal stress by ribosomal profiling and RNA-Seq. BMC Plant Biol. 2016, 16, 221. [Google Scholar] [CrossRef] [Green Version]
- Haider, M.S.; Zhang, C.; Kurjogi, M.M.; Pervaiz, T.; Zheng, T.; Zhang, C.; Lide, C.; Shangguan, L.; Fang, J. Insights into grapevine defense response against drought as revealed by biochemical, physiological and RNA-Seq analysis. Sci. Rep. 2017, 7, 13134. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.D.; Nacu, S. Fast and SNP-tolerant detection of complex variants and splicing in short reads. Bioinformatics 2010, 26, 873–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Singh, D.; Zeng, Z.; Coleman, S.J.; Huang, Y.; Savich, G.L.; He, X.; Mieczkowski, P.; Grimm, S.A.; Perou, C.M.; et al. MapSplice: Accurate mapping of RNA-seq reads for splice junction discovery. Nucleic Acids Res. 2010, 38, e178. [Google Scholar] [CrossRef] [Green Version]
- Engström, P.G.; Steijger, T.; Sipos, B.; Grant, G.R.; Kahles, A.; Rätsch, G.; Goldman, N.; Hubbard, T.J.; Harrow, J.; Guigó, R.; et al. Systematic evaluation of spliced alignment programs for RNA-seq data. Nat. Methods 2013, 10, 1185–1191. [Google Scholar] [CrossRef]
- EcSeq Best RNA-Seq Aligner: A Comparison of Mapping Tools. Available online: https://www.ecseq.com/support/ngs/best-RNA-seq-aligner-comparison-of-mapping-tools (accessed on 10 January 2020).
- Evans, C.; Hardin, J.; Stoebel, D.M. Selecting between-sample RNA-Seq normalization methods from the perspective of their assumptions. Brief. Bioinform. 2018, 19, 776–792. [Google Scholar] [CrossRef]
- Dillies, M.-A.; Rau, A.; Aubert, J.; Hennequet-Antier, C.; Jeanmougin, M.; Servant, N.; Keime, C.; Marot, G.; Castel, D.; Estelle, J.; et al. A comprehensive evaluation of normalization methods for Illumina high-throughput RNA sequencing data analysis. Brief. Bioinform. 2013, 14, 671–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullard, J.H.; Purdom, E.; Hansen, K.D.; Dudoit, S. Evaluation of statistical methods for normalization and differential expression in mRNA-Seq experiments. BMC Bioinform. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Golovnina, K.; Chen, Z.-X.; Lee, H.N.; Negron, Y.L.S.; Sultana, H.; Oliver, B.; Harbison, S.T. Comparison of normalization and differential expression analyses using RNA-Seq data from 726 individual Drosophila melanogaster. BMC Genom. 2016, 17, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapaport, F.; Khanin, R.; Liang, Y.; Pirun, M.; Krek, A.; Zumbo, P.; Mason, C.E.; Socci, N.D.; Betel, D. Comprehensive evaluation of differential gene expression analysis methods for RNA-seq data. Genome Biol. 2013, 14, R95. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, J.A.; Yang, S.S.; Miller, S.S.; Bucciarelli, B.; Liu, J.; Rydeen, A.; Bozsoki, Z.; Uhde-Stone, C.; Tu, Z.J.; Allan, D.; et al. An RNA-Seq Transcriptome Analysis of Orthophosphate-Deficient White Lupin Reveals Novel Insights into Phosphorus Acclimation in Plants. Plant Physiol. 2013, 161, 705–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Gao, Y.; Wang, J. Transcriptomic Analysis of Rice (Oryza sativa) Developing Embryos Using the RNA-Seq Technique. PLoS ONE 2012, 7, e30646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Xu, H.; Shen, Y.; Wang, J. Transcriptomic analysis of rice (Oryza sativa) endosperm using the RNA-Seq technique. Plant Mol. Biol. 2013, 81, 363–378. [Google Scholar] [CrossRef]
- Yang, S.; Hao, D.; Song, Z.; Yang, G.; Wang, L.; Su, Y. RNA-Seq analysis of differentially expressed genes in rice under varied nitrogen supplies. Gene 2015, 555, 305–317. [Google Scholar] [CrossRef]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A high resolution map of the Arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.D. PANTHER: A Library of Protein Families and Subfamilies Indexed by Function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Liu, Y.-P.; Peremyslov, V.V.; Medina, V.; Dolja, V.V. Tandem leader proteases of Grapevine leafroll-associated virus-2: Host-specific functions in the infection cycle. Virology 2009, 383, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarugula, S.; Gowda, S.; Dawson, W.O.; Naidu, R.A. Development of infectious cDNA clones of Grapevine leafroll-associated virus 3 and analyses of the 5′ non-translated region for replication and virion formation. Virology 2018, 523, 89–99. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Virus Genus | Natural Transmission (Biological Vector) | Manual Transmission |
|---|---|---|---|
| GLRaV-1 | Ampelovirus (Subgroup I) | Mealybugs and soft scale insects | Global exchange and propagation of virus-infected grapevine material |
| GLRaV-2 | Closterovirus | Not identified | |
| GLRaV-3 | Ampelovirus (Subgroup I) | Mealybugs and soft scale insects | |
| GLRaV-4 | Ampelovirus (Subgroup II) | Mealybugs and soft scale insects | |
| GLRaV-7 | Velarivirus | Parasitic dodder plants |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Hanner, R.H.; Meng, B. Probing into the Effects of Grapevine Leafroll-Associated Viruses on the Physiology, Fruit Quality and Gene Expression of Grapes. Viruses 2021, 13, 593. https://doi.org/10.3390/v13040593
Song Y, Hanner RH, Meng B. Probing into the Effects of Grapevine Leafroll-Associated Viruses on the Physiology, Fruit Quality and Gene Expression of Grapes. Viruses. 2021; 13(4):593. https://doi.org/10.3390/v13040593
Chicago/Turabian StyleSong, Yashu, Robert H. Hanner, and Baozhong Meng. 2021. "Probing into the Effects of Grapevine Leafroll-Associated Viruses on the Physiology, Fruit Quality and Gene Expression of Grapes" Viruses 13, no. 4: 593. https://doi.org/10.3390/v13040593
APA StyleSong, Y., Hanner, R. H., & Meng, B. (2021). Probing into the Effects of Grapevine Leafroll-Associated Viruses on the Physiology, Fruit Quality and Gene Expression of Grapes. Viruses, 13(4), 593. https://doi.org/10.3390/v13040593