
Antivirals Targeting the Surface Glycoproteins of Influenza Virus: Mechanisms of Action and Resistance


Abstract
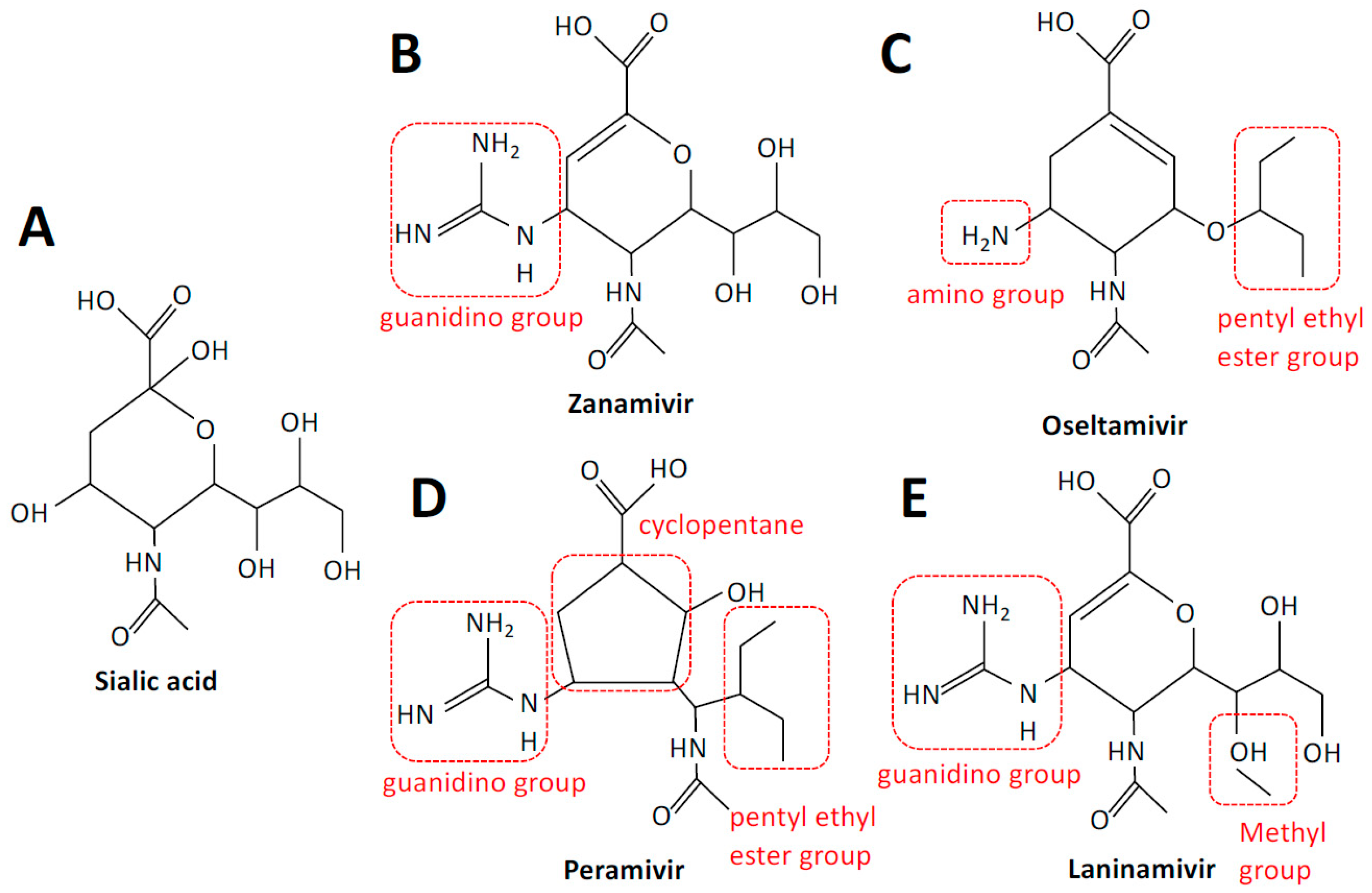
:1. Introduction
2. Neuraminidase Inhibitors
2.1. Neuraminidase Inhibitors—Mechanisms of Action
2.2. Epidemiology of NAI Resistance
2.2.1. Zanamivir
2.2.2. Oseltamivir
2.2.3. Peramivir and Laninamivir
2.3. Functional Neuraminidase Inhibitor Resistance Motifs
2.3.1. Zanamivir
Resistance in Influenza A Viruses
Resistance in Influenza B Viruses
2.3.2. Oseltamivir
Resistance in Influenza A Viruses
Resistance in Influenza B Viruses
2.3.3. Peramivir
Resistance in Influenza A Viruses
Resistance in Influenza B Viruses
2.3.4. Laninamivir
Resistance in Influenza A Viruses
Resistance in Influenza B Viruses
3. Hemagglutinin Inhibitors
3.1. HA Inhibitors—Mechanisms of Action and Resistance
3.1.1. Umifenovir
3.1.2. Newer HA Fusion Inhibitors
3.1.3. Peptide Fusion Inhibitors
3.1.4. Receptor Binding Site Inhibitors
4. Therapeutic Strategies to Combat Drug Resistance
5. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hay, A.J.; Wolstenholme, A.J.; Skehel, J.J.; Smith, M.H. The molecular basis of the specific anti-influenza action of amantadine. EMBO J. 1985, 4, 3021–3024. [Google Scholar] [CrossRef]
- Bright, R.A.; Shay, D.K.; Shu, B.; Cox, N.J.; Klimov, A.I. Adamantane resistance among influenza A viruses isolated early during the 2005-2006 influenza season in the United States. JAMA 2006, 295, 891–894. [Google Scholar] [CrossRef] [Green Version]
- Furuta, Y.; Gowen, B.B.; Takahashi, K.; Shiraki, K.; Smee, D.F.; Barnard, D.L. Favipiravir (T-705), a novel viral RNA polymerase inhibitor. Antivir. Res. 2013, 100, 446–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oakley, A.J.; Barrett, S.; Peat, T.S.; Newman, J.; Streltsov, V.A.; Waddington, L.; Saito, T.; Tashiro, M.; McKimm-Breschkin, J.L. Structural and functional basis of resistance to neuraminidase inhibitors of influenza B viruses. J. Med. Chem. 2010, 53, 6421–6431. [Google Scholar] [CrossRef] [PubMed]
- McAuley, J.L.; Gilbertson, B.P.; Trifkovic, S.; Brown, L.E.; McKimm-Breschkin, J.L. Influenza Virus Neuraminidase Structure and Functions. Front. Microbiol. 2019, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, K.C.; Hung, H.C.; Horng, J.T.; Fang, M.Y.; Chang, C.Y.; Li, L.T.; Chen, I.J.; Chen, Y.C.; Chou, D.L.; Chang, C.W.; et al. Parallel screening of wild-type and drug-resistant targets for anti-resistance neuraminidase inhibitors. PLoS ONE 2013, 8, e56704. [Google Scholar] [CrossRef]
- Maring, C.J.; Stoll, V.S.; Zhao, C.; Sun, M.; Krueger, A.C.; Stewart, K.D.; Madigan, D.L.; Kati, W.M.; Xu, Y.; Carrick, R.J.; et al. Structure-based characterization and optimization of novel hydrophobic binding interactions in a series of pyrrolidine influenza neuraminidase inhibitors. J. Med. Chem. 2005, 48, 3980–3990. [Google Scholar] [CrossRef]
- Stoll, V.; Stewart, K.D.; Maring, C.J.; Muchmore, S.; Giranda, V.; Gu, Y.G.; Wang, G.; Chen, Y.; Sun, M.; Zhao, C.; et al. Influenza neuraminidase inhibitors: Structure-based design of a novel inhibitor series. Biochemistry 2003, 42, 718–727. [Google Scholar] [CrossRef]
- Marquez-Dominguez, L.; Reyes-Leyva, J.; Herrera-Camacho, I.; Santos-Lopez, G.; Scior, T. Five Novel Non-Sialic Acid-Like Scaffolds Inhibit In Vitro H1N1 and H5N2 Neuraminidase Activity of Influenza a Virus. Molecules 2020, 25, 4248. [Google Scholar] [CrossRef]
- Alame, M.M.; Massaad, E.; Zaraket, H. Peramivir: A Novel Intravenous Neuraminidase Inhibitor for Treatment of Acute Influenza Infections. Front. Microbiol 2016, 7, 450. [Google Scholar] [CrossRef]
- Komeda, T.; Ishii, S.; Itoh, Y.; Sanekata, M.; Yoshikawa, T.; Shimada, J. Post-marketing safety evaluation of the intravenous anti-influenza neuraminidase inhibitor peramivir: A drug-use investigation in patients with high risk factors. J. Infect. Chemother. 2016, 22, 677–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKimm-Breschkin, J.L. Influenza neuraminidase inhibitors: Antiviral action and mechanisms of resistance. Influenza Other Respir. Viruses 2013, 7 (Suppl. 1), 25–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diggory, P.; Fernandez, C.; Humphrey, A.; Jones, V.; Murphy, M. Comparison of elderly people’s technique in using two dry powder inhalers to deliver zanamivir: Randomised controlled trial. BMJ 2001, 322, 577–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Medicines Agency. Summary on Compassionate Use for IV Zanamivir. Available online: https://www.ema.europa.eu/en/documents/other/summary-compassionate-use-iv-zanamivirrev-1_en.pdf (accessed on 15 February 2021).
- Davies, B.E. Pharmacokinetics of oseltamivir: An oral antiviral for the treatment and prophylaxis of influenza in diverse populations. J. Antimicrob. Chemother. 2010, 65 (Suppl. 2), ii5–ii10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, R.; Kodama, M.; Kobayashi, M.; Kitano, M.; Sato, A.; Yamano, Y. Therapeutic effect of peramivir (S-021812, BCX-1812) after single intravenous treatment of mice infected with influenza A and B viruses. In 52nd Interscience Conference on Antimicrobial Agents and Chemotherapy; American Society for Microbiology: Washington, DC, USA, 2009. [Google Scholar]
- Ikematsu, H.; Kawai, N. Laninamivir octanoate: A new long-acting neuraminidase inhibitor for the treatment of influenza. Expert Rev. Anti-Infect. 2011, 9, 851–857. [Google Scholar] [CrossRef]
- Hanpaibool, C.; Leelawiwat, M.; Takahashi, K.; Rungrotmongkol, T. Source of oseltamivir resistance due to single E119D and double E119D/H274Y mutations in pdm09H1N1 influenza neuraminidase. J. Comput. Mol. Des. 2020, 34, 27–37. [Google Scholar] [CrossRef]
- Prachanronarong, K.L.; Ozen, A.; Thayer, K.M.; Yilmaz, L.S.; Zeldovich, K.B.; Bolon, D.N.; Kowalik, T.F.; Jensen, J.D.; Finberg, R.W.; Wang, J.P.; et al. Molecular Basis for Differential Patterns of Drug Resistance in Influenza N1 and N2 Neuraminidase. J. Chem. Theory Comput. 2016, 12, 6098–6108. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, J.; Li, W.; Wang, X.; Xu, J.; Xie, J.; Tao, K.; Shen, L.; Zhang, R. Molecular docking of potential inhibitors for influenza H7N9. Comput Math. Methods Med. 2015, 2015, 480764. [Google Scholar] [CrossRef]
- Babu, Y.S.; Chand, P.; Bantia, S.; Kotian, P.; Dehghani, A.; El-Kattan, Y.; Lin, T.H.; Hutchison, T.L.; Elliott, A.J.; Parker, C.D.; et al. BCX-1812 (RWJ-270201): Discovery of a novel, highly potent, orally active, and selective influenza neuraminidase inhibitor through structure-based drug design. J. Med. Chem. 2000, 43, 3482–3486. [Google Scholar] [CrossRef]
- Meijer, A.; Rebelo-de-Andrade, H.; Correia, V.; Besselaar, T.; Drager-Dayal, R.; Fry, A.; Gregory, V.; Gubareva, L.; Kageyama, T.; Lackenby, A.; et al. Global update on the susceptibility of human influenza viruses to neuraminidase inhibitors, 2012–2013. Antivir. Res. 2014, 110, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Takashita, E.; Meijer, A.; Lackenby, A.; Gubareva, L.; Rebelo-de-Andrade, H.; Besselaar, T.; Fry, A.; Gregory, V.; Leang, S.K.; Huang, W.; et al. Global update on the susceptibility of human influenza viruses to neuraminidase inhibitors, 2013–2014. Antivir. Res. 2015, 117, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Hurt, A.C.; Besselaar, T.G.; Daniels, R.S.; Ermetal, B.; Fry, A.; Gubareva, L.; Huang, W.; Lackenby, A.; Lee, R.T.; Lo, J.; et al. Global update on the susceptibility of human influenza viruses to neuraminidase inhibitors, 2014–2015. Antivir. Res. 2016, 132, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Gubareva, L.V.; Besselaar, T.G.; Daniels, R.S.; Fry, A.; Gregory, V.; Huang, W.; Hurt, A.C.; Jorquera, P.A.; Lackenby, A.; Leang, S.K.; et al. Global update on the susceptibility of human influenza viruses to neuraminidase inhibitors, 2015–2016. Antivir. Res. 2017, 146, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Lackenby, A.; Besselaar, T.G.; Daniels, R.S.; Fry, A.; Gregory, V.; Gubareva, L.V.; Huang, W.; Hurt, A.C.; Leang, S.K.; Lee, R.T.C.; et al. Global update on the susceptibility of human influenza viruses to neuraminidase inhibitors and status of novel antivirals, 2016-2017. Antivir. Res. 2018, 157, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Takashita, E.; Daniels, R.S.; Fujisaki, S.; Gregory, V.; Gubareva, L.V.; Huang, W.; Hurt, A.C.; Lackenby, A.; Nguyen, H.T.; Pereyaslov, D.; et al. Global update on the susceptibilities of human influenza viruses to neuraminidase inhibitors and the cap-dependent endonuclease inhibitor baloxavir, 2017–2018. Antivir. Res. 2020, 175, 104718. [Google Scholar] [CrossRef]
- Li, T.C.; Chan, M.C.; Lee, N. Clinical Implications of Antiviral Resistance in Influenza. Viruses 2015, 7, 4929–4944. [Google Scholar] [CrossRef] [Green Version]
- Baek, Y.H.; Song, M.S.; Lee, E.Y.; Kim, Y.I.; Kim, E.H.; Park, S.J.; Park, K.J.; Kwon, H.I.; Pascua, P.N.; Lim, G.J.; et al. Profiling and characterization of influenza virus N1 strains potentially resistant to multiple neuraminidase inhibitors. J. Virol. 2015, 89, 287–299. [Google Scholar] [CrossRef] [Green Version]
- L’Huillier, A.G.; Abed, Y.; Petty, T.J.; Cordey, S.; Thomas, Y.; Bouhy, X.; Schibler, M.; Simon, A.; Chalandon, Y.; van Delden, C.; et al. E119D Neuraminidase Mutation Conferring Pan-Resistance to Neuraminidase Inhibitors in an A(H1N1)pdm09 Isolate From a Stem-Cell Transplant Recipient. J. Infect. Dis. 2015, 212, 1726–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloren, K.K.S.; Kwon, J.J.; Choi, W.S.; Jeong, J.H.; Ahn, S.J.; Choi, Y.K.; Baek, Y.H.; Song, M.S. In Vitro and In Vivo Characterization of Novel Neuraminidase Substitutions in Influenza A(H1N1)pdm09 Virus Identified Using Laninamivir-Mediated In Vitro Selection. J. Virol. 2019, 93, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzorno, A.; Bouhy, X.; Abed, Y.; Boivin, G. Generation and characterization of recombinant pandemic influenza A(H1N1) viruses resistant to neuraminidase inhibitors. J. Infect. Dis. 2011, 203, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, D.; DeBiasi, R.L.; Okomo-Adhiambo, M.; Mishin, V.P.; Campbell, A.P.; Loechelt, B.; Wiedermann, B.L.; Fry, A.M.; Gubareva, L.V. Emergence of Multidrug-Resistant Influenza A(H1N1)pdm09 Virus Variants in an Immunocompromised Child Treated With Oseltamivir and Zanamivir. J. Infect. Dis. 2015, 212, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- Baz, M.; Abed, Y.; Papenburg, J.; Bouhy, X.; Hamelin, M.E.; Boivin, G. Emergence of oseltamivir-resistant pandemic H1N1 virus during prophylaxis. N. Engl. J. Med. 2009, 361, 2296–2297. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Trujillo, A.A.; Sheu, T.G.; Levine, M.; Mishin, V.P.; Shaw, M.; Ades, E.W.; Klimov, A.I.; Fry, A.M.; Gubareva, L.V. Analysis of influenza viruses from patients clinically suspected of infection with an oseltamivir resistant virus during the 2009 pandemic in the United States. Antivir. Res. 2012, 93, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Leang, S.K.; Kwok, S.; Sullivan, S.G.; Maurer-Stroh, S.; Kelso, A.; Barr, I.G.; Hurt, A.C. Peramivir and laninamivir susceptibility of circulating influenza A and B viruses. Influenza Other Respir. Viruses 2014, 8, 135–139. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Nguyen, T.; Mishin, V.P.; Sleeman, K.; Balish, A.; Jones, J.; Creanga, A.; Marjuki, H.; Uyeki, T.M.; Nguyen, D.H.; et al. Antiviral susceptibility of highly pathogenic avian influenza A(H5N1) viruses isolated from poultry, Vietnam, 2009–2011. Emerg. Infect. Dis. 2013, 19, 1963–1971. [Google Scholar] [CrossRef] [Green Version]
- Le, Q.M.; Kiso, M.; Someya, K.; Sakai, Y.T.; Nguyen, T.H.; Nguyen, K.H.; Pham, N.D.; Ngyen, H.H.; Yamada, S.; Muramoto, Y.; et al. Avian flu: Isolation of drug-resistant H5N1 virus. Nature 2005, 437, 1108. [Google Scholar] [CrossRef]
- Sheu, T.G.; Deyde, V.M.; Okomo-Adhiambo, M.; Garten, R.J.; Xu, X.; Bright, R.A.; Butler, E.N.; Wallis, T.R.; Klimov, A.I.; Gubareva, L.V. Surveillance for neuraminidase inhibitor resistance among human influenza A and B viruses circulating worldwide from 2004 to 2008. Antimicrob. Agents Chemother. 2008, 52, 3284–3292. [Google Scholar] [CrossRef] [Green Version]
- Abed, Y.; Baz, M.; Boivin, G. Impact of neuraminidase mutations conferring influenza resistance to neuraminidase inhibitors in the N1 and N2 genetic backgrounds. Antivir. Ther. 2006, 11, 971–976. [Google Scholar]
- Mishin, V.P.; Hayden, F.G.; Gubareva, L.V. Susceptibilities of antiviral-resistant influenza viruses to novel neuraminidase inhibitors. Antimicrob. Agents Chemother. 2005, 49, 4515–4520. [Google Scholar] [CrossRef] [Green Version]
- Okomo-Adhiambo, M.; Demmler-Harrison, G.J.; Deyde, V.M.; Sheu, T.G.; Xu, X.; Klimov, A.I.; Gubareva, L.V. Detection of E119V and E119I mutations in influenza A (H3N2) viruses isolated from an immunocompromised patient: Challenges in diagnosis of oseltamivir resistance. Antimicrob. Agents Chemother. 2010, 54, 1834–1841. [Google Scholar] [CrossRef] [Green Version]
- Tamura, D.; Sugaya, N.; Ozawa, M.; Takano, R.; Ichikawa, M.; Yamazaki, M.; Kawakami, C.; Shimizu, H.; Uehara, R.; Kiso, M.; et al. Frequency of drug-resistant viruses and virus shedding in pediatric influenza patients treated with neuraminidase inhibitors. Clin. Infect. Dis. 2011, 52, 432–437. [Google Scholar] [CrossRef]
- Simon, P.; Holder, B.P.; Bouhy, X.; Abed, Y.; Beauchemin, C.A.; Boivin, G. The I222V neuraminidase mutation has a compensatory role in replication of an oseltamivir-resistant influenza virus A/H3N2 E119V mutant. J. Clin. Microbiol. 2011, 49, 715–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiso, M.; Mitamura, K.; Sakai-Tagawa, Y.; Shiraishi, K.; Kawakami, C.; Kimura, K.; Hayden, F.G.; Sugaya, N.; Kawaoka, Y. Resistant influenza A viruses in children treated with oseltamivir: Descriptive study. Lancet 2004, 364, 759–765. [Google Scholar] [CrossRef]
- Fujisaki, S.; Takashita, E.; Yokoyama, M.; Taniwaki, T.; Xu, H.; Kishida, N.; Sato, H.; Tashiro, M.; Imai, M.; Odagiri, T. A single E105K mutation far from the active site of influenza B virus neuraminidase contributes to reduced susceptibility to multiple neuraminidase-inhibitor drugs. Biochem. Biophys. Res. Commun. 2012, 429, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Sheu, T.G.; Deyde, V.M.; Garten, R.J.; Klimov, A.I.; Gubareva, L.V. Detection of antiviral resistance and genetic lineage markers in influenza B virus neuraminidase using pyrosequencing. Antivir. Res. 2010, 85, 354–360. [Google Scholar] [CrossRef]
- Burnham, A.J.; Baranovich, T.; Marathe, B.M.; Armstrong, J.; Webster, R.G.; Govorkova, E.A. Fitness costs for Influenza B viruses carrying neuraminidase inhibitor-resistant substitutions: Underscoring the importance of E119A and H274Y. Antimicrob. Agents Chemother. 2014, 58, 2718–2730. [Google Scholar] [CrossRef] [Green Version]
- Gubareva, L.V.; Webster, R.G.; Hayden, F.G. Comparison of the activities of zanamivir, oseltamivir, and RWJ-270201 against clinical isolates of influenza virus and neuraminidase inhibitor-resistant variants. Antimicrob. Agents Chemother. 2001, 45, 3403–3408. [Google Scholar] [CrossRef] [Green Version]
- Gubareva, L.V.; Matrosovich, M.N.; Brenner, M.K.; Bethell, R.C.; Webster, R.G. Evidence for zanamivir resistance in an immunocompromised child infected with influenza B virus. J. Infect. Dis. 1998, 178, 1257–1262. [Google Scholar] [CrossRef]
- Wang, D.; Sleeman, K.; Huang, W.; Nguyen, H.T.; Levine, M.; Cheng, Y.; Li, X.; Tan, M.; Xing, X.; Xu, X.; et al. Neuraminidase inhibitor susceptibility testing of influenza type B viruses in China during 2010 and 2011 identifies viruses with reduced susceptibility to oseltamivir and zanamivir. Antivir. Res. 2013, 97, 240–244. [Google Scholar] [CrossRef]
- Carr, S.; Ilyushina, N.A.; Franks, J.; Adderson, E.E.; Caniza, M.; Govorkova, E.A.; Webster, R.G. Oseltamivir-resistant influenza A and B viruses pre- and postantiviral therapy in children and young adults with cancer. Pediatr. Infect. Dis. J. 2011, 30, 284–288. [Google Scholar] [CrossRef] [Green Version]
- Hurt, A.C.; Chotpitayasunondh, T.; Cox, N.J.; Daniels, R.; Fry, A.M.; Gubareva, L.V.; Hayden, F.G.; Hui, D.S.; Hungnes, O.; Lackenby, A.; et al. Antiviral resistance during the 2009 influenza A H1N1 pandemic: Public health, laboratory, and clinical perspectives. Lancet Infect. Dis. 2012, 12, 240–248. [Google Scholar] [CrossRef]
- Oh, D.Y.; Panozzo, J.; Vitesnik, S.; Farrukee, R.; Piedrafita, D.; Mosse, J.; Hurt, A.C. Selection of multi-drug resistant influenza A and B viruses under zanamivir pressure and their replication fitness in ferrets. Antivir. Ther. 2018, 23, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yates, P.J.; Raimonde, D.S.; Zhao, H.H.; Man, C.Y.; Steel, H.M.; Mehta, N.; Peppercorn, A.F. Phenotypic and genotypic analysis of influenza viruses isolated from adult subjects during a phase II study of intravenous zanamivir in hospitalised subjects. Antivir. Res. 2016, 134, 144–152. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Fry, A.M.; Loveless, P.A.; Klimov, A.I.; Gubareva, L.V. Recovery of a multidrug-resistant strain of pandemic influenza A 2009 (H1N1) virus carrying a dual H275Y/I223R mutation from a child after prolonged treatment with oseltamivir. Clin. Infect. Dis. 2010, 51, 983–984. [Google Scholar] [CrossRef] [Green Version]
- Dapat, C.; Suzuki, Y.; Saito, R.; Kyaw, Y.; Myint, Y.Y.; Lin, N.; Oo, H.N.; Oo, K.Y.; Win, N.; Naito, M.; et al. Rare influenza A (H3N2) variants with reduced sensitivity to antiviral drugs. Emerg. Infect. Dis. 2010, 16, 493–496. [Google Scholar] [CrossRef]
- Hurt, A.C.; Holien, J.K.; Parker, M.; Kelso, A.; Barr, I.G. Zanamivir-resistant influenza viruses with a novel neuraminidase mutation. J. Virol. 2009, 83, 10366–10373. [Google Scholar] [CrossRef] [Green Version]
- CDC Situation Update: Summary of Weekly FluView Report, Seasonal Influenza (Flu). Available online: https://espanol.cdc.gov/flu/weekly/weeklyarchives2013-2014/weekly45.htm?mobile=nocontent (accessed on 15 November 2013).
- Monto, A.S.; McKimm-Breschkin, J.L.; Macken, C.; Hampson, A.W.; Hay, A.; Klimov, A.; Tashiro, M.; Webster, R.G.; Aymard, M.; Hayden, F.G.; et al. Detection of influenza viruses resistant to neuraminidase inhibitors in global surveillance during the first 3 years of their use. Antimicrob. Agents Chemother. 2006, 50, 2395–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurt, A.C.; Barr, I.G.; Hartel, G.; Hampson, A.W. Susceptibility of human influenza viruses from Australasia and South East Asia to the neuraminidase inhibitors zanamivir and oseltamivir. Antivir. Res. 2004, 62, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Escuret, V.; Frobert, E.; Bouscambert-Duchamp, M.; Sabatier, M.; Grog, I.; Valette, M.; Lina, B.; Morfin, F.; Ferraris, O. Detection of human influenza A (H1N1) and B strains with reduced sensitivity to neuraminidase inhibitors. J. Clin. Virol. 2008, 41, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Ives, J.A.; Carr, J.A.; Mendel, D.B.; Tai, C.Y.; Lambkin, R.; Kelly, L.; Oxford, J.S.; Hayden, F.G.; Roberts, N.A. The H274Y mutation in the influenza A/H1N1 neuraminidase active site following oseltamivir phosphate treatment leave virus severely compromised both in vitro and in vivo. Antivir. Res. 2002, 55, 307–317. [Google Scholar] [CrossRef]
- Bloom, J.D.; Gong, L.I.; Baltimore, D. Permissive secondary mutations enable the evolution of influenza oseltamivir resistance. Science 2010, 328, 1272–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, J.; Hooper, K.A.; Petrie, S.; Lee, R.; Maurer-Stroh, S.; Reh, L.; Guarnaccia, T.; Baas, C.; Xue, L.; Vitesnik, S.; et al. Estimating the fitness advantage conferred by permissive neuraminidase mutations in recent oseltamivir-resistant A(H1N1)pdm09 influenza viruses. PLoS Pathog. 2014, 10, e1004065. [Google Scholar] [CrossRef] [Green Version]
- Dharan, N.J.; Gubareva, L.V.; Meyer, J.J.; Okomo-Adhiambo, M.; McClinton, R.C.; Marshall, S.A.; St George, K.; Epperson, S.; Brammer, L.; Klimov, A.I.; et al. Infections with oseltamivir-resistant influenza A(H1N1) virus in the United States. JAMA 2009, 301, 1034–1041. [Google Scholar] [CrossRef]
- World Health Organization. Influenza A(H1N1) Virus Resistance to Oseltamivir. Available online: https://www.who.int/influenza/patient_care/antivirals/oseltamivir_summary/en/ (accessed on 15 February 2021).
- Gooskens, J.; Jonges, M.; Claas, E.C.; Meijer, A.; Kroes, A.C. Prolonged influenza virus infection during lymphocytopenia and frequent detection of drug-resistant viruses. J. Infect. Dis. 2009, 199, 1435–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashita, E.; Fujisaki, S.; Shirakura, M.; Nakamura, K.; Kishida, N.; Kuwahara, T.; Shimazu, Y.; Shimomura, T.; Watanabe, S.; Odagiri, T.; et al. Influenza A(H1N1)pdm09 virus exhibiting enhanced cross-resistance to oseltamivir and peramivir due to a dual H275Y/G147R substitution, Japan, March 2016. Eurosurveillance 2016, 21, 24. [Google Scholar] [CrossRef]
- Takashita, E.; Fujisaki, S.; Yokoyama, M.; Shirakura, M.; Morita, H.; Nakamura, K.; Kishida, N.; Kuwahara, T.; Sato, H.; Doi, I.; et al. In Vitro Characterization of Multidrug-Resistant Influenza A(H1N1)pdm09 Viruses Carrying a Dual Neuraminidase Mutation Isolated from Immunocompromised Patients. Pathogens 2020, 9, 725. [Google Scholar] [CrossRef]
- Lina, B.; Boucher, C.; Osterhaus, A.; Monto, A.S.; Schutten, M.; Whitley, R.J.; Nguyen-Van-Tam, J.S. Five years of monitoring for the emergence of oseltamivir resistance in patients with influenza A infections in the Influenza Resistance Information Study. Influenza Other Respir. Viruses 2018, 12, 267–278. [Google Scholar] [CrossRef]
- Okomo-Adhiambo, M.; Sleeman, K.; Lysen, C.; Nguyen, H.T.; Xu, X.; Li, Y.; Klimov, A.I.; Gubareva, L.V. Neuraminidase inhibitor susceptibility surveillance of influenza viruses circulating worldwide during the 2011 Southern Hemisphere season. Influenza Other Respir. Viruses 2013, 7, 645–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samson, M.; Pizzorno, A.; Abed, Y.; Boivin, G. Influenza virus resistance to neuraminidase inhibitors. Antivir. Res. 2013, 98, 174–185. [Google Scholar] [CrossRef]
- Zurcher, T.; Yates, P.J.; Daly, J.; Sahasrabudhe, A.; Walters, M.; Dash, L.; Tisdale, M.; McKimm-Breschkin, J.L. Mutations conferring zanamivir resistance in human influenza virus N2 neuraminidases compromise virus fitness and are not stably maintained in vitro. J. Antimicrob. Chemother. 2006, 58, 723–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eshaghi, A.; Shalhoub, S.; Rosenfeld, P.; Li, A.; Higgins, R.R.; Stogios, P.J.; Savchenko, A.; Bastien, N.; Li, Y.; Rotstein, C.; et al. Multiple influenza A (H3N2) mutations conferring resistance to neuraminidase inhibitors in a bone marrow transplant recipient. Antimicrob. Agents Chemother. 2014, 58, 7188–7197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaymard, A.; Charles-Dufant, A.; Sabatier, M.; Cortay, J.C.; Frobert, E.; Picard, C.; Casalegno, J.S.; Rosa-Calatrava, M.; Ferraris, O.; Valette, M.; et al. Impact on antiviral resistance of E119V, I222L and R292K substitutions in influenza A viruses bearing a group 2 neuraminidase (N2, N3, N6, N7 and N9). J. Antimicrob. Chemother. 2016, 71, 3036–3045. [Google Scholar] [CrossRef] [PubMed]
- Eshaghi, A.; Patel, S.N.; Sarabia, A.; Higgins, R.R.; Savchenko, A.; Stojios, P.J.; Li, Y.; Bastien, N.; Alexander, D.C.; Low, D.E.; et al. Multidrug-resistant pandemic (H1N1) 2009 infection in immunocompetent child. Emerg. Infect. Dis. 2011, 17, 1472–1474. [Google Scholar] [CrossRef] [PubMed]
- van der Vries, E.; Stelma, F.F.; Boucher, C.A. Emergence of a multidrug-resistant pandemic influenza A (H1N1) virus. N. Engl. J. Med. 2010, 363, 1381–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, K.; Leang, S.K.; Butler, J.; Baas, C.; Harrower, B.; Mosse, J.; Barr, I.G.; Hurt, A.C. Zanamivir-resistant influenza viruses with Q136K or Q136R neuraminidase residue mutations can arise during MDCK cell culture creating challenges for antiviral susceptibility monitoring. Eurosurveillance 2015, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.; Barclay, W.; Zurcher, T. Characterization of recombinant influenza B viruses with key neuraminidase inhibitor resistance mutations. J. Antimicrob. Chemother. 2005, 55, 162–169. [Google Scholar] [CrossRef]
- Baz, M.; Abed, Y.; Simon, P.; Hamelin, M.E.; Boivin, G. Effect of the neuraminidase mutation H274Y conferring resistance to oseltamivir on the replicative capacity and virulence of old and recent human influenza A(H1N1) viruses. J. Infect. Dis. 2010, 201, 740–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashita, E.; Kiso, M.; Fujisaki, S.; Yokoyama, M.; Nakamura, K.; Shirakura, M.; Sato, H.; Odagiri, T.; Kawaoka, Y.; Tashiro, M. Characterization of a large cluster of influenza A(H1N1)pdm09 viruses cross-resistant to oseltamivir and peramivir during the 2013–2014 influenza season in Japan. Antimicrob. Agents Chemother. 2015, 59, 2607–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzorno, A.; Abed, Y.; Bouhy, X.; Beaulieu, E.; Mallett, C.; Russell, R.; Boivin, G. Impact of mutations at residue I223 of the neuraminidase protein on the resistance profile, replication level, and virulence of the 2009 pandemic influenza virus. Antimicrob. Agents Chemother. 2012, 56, 1208–1214. [Google Scholar] [CrossRef] [Green Version]
- LeGoff, J.; Rousset, D.; Abou-Jaoude, G.; Scemla, A.; Ribaud, P.; Mercier-Delarue, S.; Caro, V.; Enouf, V.; Simon, F.; Molina, J.M.; et al. I223R mutation in influenza A(H1N1)pdm09 neuraminidase confers reduced susceptibility to oseltamivir and zanamivir and enhanced resistance with H275Y. PLoS ONE 2012, 7, e37095. [Google Scholar] [CrossRef]
- Hurt, A.C.; Lee, R.T.; Leang, S.K.; Cui, L.; Deng, Y.M.; Phuah, S.P.; Caldwell, N.; Freeman, K.; Komadina, N.; Smith, D.; et al. Increased detection in Australia and Singapore of a novel influenza A(H1N1)2009 variant with reduced oseltamivir and zanamivir sensitivity due to a S247N neuraminidase mutation. Eurosurveillance 2011, 16, 23. [Google Scholar] [CrossRef]
- Gubareva, L.V.; Sleeman, K.; Guo, Z.; Yang, H.; Hodges, E.; Davis, C.T.; Baranovich, T.; Stevens, J. Drug Susceptibility Evaluation of an Influenza A(H7N9) Virus by Analyzing Recombinant Neuraminidase Proteins. J. Infect. Dis. 2017, 216 (Suppl. 4), S566–S574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baum, E.Z.; Wagaman, P.C.; Ly, L.; Turchi, I.; Le, J.; Bucher, D.; Bush, K. A point mutation in influenza B neuraminidase confers resistance to peramivir and loss of slow binding. Antivir. Res. 2003, 59, 13–22. [Google Scholar] [CrossRef]
- Jeong, J.H.; Choi, W.S.; Antigua, K.J.C.; Choi, Y.K.; Govorkova, E.A.; Webby, R.J.; Baek, Y.H.; Song, M.S. In Vitro Profiling of Laninamivir-Resistant Substitutions in N3 to N9 Avian Influenza Virus Neuraminidase Subtypes and Their Association with In Vivo Susceptibility. J. Virol. 2020, 95, 1. [Google Scholar] [CrossRef]
- Sriwilaijaroen, N.; Suzuki, Y. Molecular basis of the structure and function of H1 hemagglutinin of influenza virus. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2012, 88, 226–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soundararajan, V.; Zheng, S.; Patel, N.; Warnock, K.; Raman, R.; Wilson, I.A.; Raguram, S.; Sasisekharan, V.; Sasisekharan, R. Networks link antigenic and receptor-binding sites of influenza hemagglutinin: Mechanistic insight into fitter strain propagation. Sci. Rep. 2011, 1, 200. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.C.; Wilson, I.A. Influenza Hemagglutinin Structures and Antibody Recognition. Cold Spring Harb. Perspect. Med. 2020, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Kadam, R.U.; Wilson, I.A. Structural basis of influenza virus fusion inhibition by the antiviral drug Arbidol. Proc. Natl. Acad. Sci. USA 2017, 114, 206–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leneva, I.A.; Fediakina, I.T.; Gus’kova, T.A.; Glushkov, R.G. Sensitivity of various influenza virus strains to arbidol. Influence of arbidol combination with different antiviral drugs on reproduction of influenza virus A. Ter. Arkh. 2005, 77, 84–88. [Google Scholar]
- Leneva, I.A.; Burtseva, E.I.; Yatsyshina, S.B.; Fedyakina, I.T.; Kirillova, E.S.; Selkova, E.P.; Osipova, E.; Maleev, V.V. Virus susceptibility and clinical effectiveness of anti-influenza drugs during the 2010–2011 influenza season in Russia. Int. J. Infect. Dis. 2016, 43, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Brooks, M.J.; Burtseva, E.I.; Ellery, P.J.; Marsh, G.A.; Lew, A.M.; Slepushkin, A.N.; Crowe, S.M.; Tannock, G.A. Antiviral activity of arbidol, a broad-spectrum drug for use against respiratory viruses, varies according to test conditions. J. Med. Virol. 2012, 84, 170–181. [Google Scholar] [CrossRef]
- Shi, L.; Xiong, H.; He, J.; Deng, H.; Li, Q.; Zhong, Q.; Hou, W.; Cheng, L.; Xiao, H.; Yang, Z. Antiviral activity of arbidol against influenza A virus, respiratory syncytial virus, rhinovirus, coxsackie virus and adenovirus in vitro and in vivo. Arch. Virol. 2007, 152, 1447–1455. [Google Scholar] [CrossRef]
- Boriskin, Y.S.; Leneva, I.A.; Pecheur, E.I.; Polyak, S.J. Arbidol: A broad-spectrum antiviral compound that blocks viral fusion. Curr. Med. Chem. 2008, 15, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Leneva, I.A.; Russell, R.J.; Boriskin, Y.S.; Hay, A.J. Characteristics of arbidol-resistant mutants of influenza virus: Implications for the mechanism of anti-influenza action of arbidol. Antivir. Res. 2009, 81, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Cihan-Ustundag, G.; Zopun, M.; Vanderlinden, E.; Ozkirimli, E.; Persoons, L.; Capan, G.; Naesens, L. Superior inhibition of influenza virus hemagglutinin-mediated fusion by indole-substituted spirothiazolidinones. Bioorg. Med. Chem. 2020, 28, 115130. [Google Scholar] [CrossRef]
- Shen, X.; Zhu, Z.; Ding, Y.; Wu, W.; Yang, J.; Liu, S. An oligothiophene compound neutralized influenza A viruses by interfering with hemagglutinin. Biochim. Biophys. Acta. Biomembr. 2018, 1860, 784–791. [Google Scholar] [CrossRef]
- Zhu, Z.; Yao, Z.; Shen, X.; Chen, Z.; Liu, X.; Parquette, J.R.; Liu, S. Oligothiophene compounds inhibit the membrane fusion between H5N1 avian influenza virus and the endosome of host cell. Eur. J. Med. Chem. 2017, 130, 185–194. [Google Scholar] [CrossRef]
- Song, G.; Shen, X.; Li, S.; Li, Y.; Si, H.; Fan, J.; Li, J.; Gao, E.; Liu, S. Structure-activity relationships of 3-O-beta-chacotriosyl oleanane-type triterpenoids as potential H5N1 entry inhibitors. Eur. J. Med. Chem. 2016, 119, 109–121. [Google Scholar] [CrossRef]
- Wu, W.; Li, R.; Li, X.; He, J.; Jiang, S.; Liu, S.; Yang, J. Quercetin as an Antiviral Agent Inhibits Influenza A Virus (IAV) Entry. Viruses 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Li, Y.; Li, S.; Li, H.; Qiu, Z.; Lee, C.; Lu, H.; Lin, X.; Zhao, R.; Chen, L.; et al. Inhibition of influenza A virus (H1N1) fusion by benzenesulfonamide derivatives targeting viral hemagglutinin. PLoS ONE 2011, 6, e29120. [Google Scholar] [CrossRef] [PubMed]
- van Dongen, M.J.P.; Kadam, R.U.; Juraszek, J.; Lawson, E.; Brandenburg, B.; Schmitz, F.; Schepens, W.B.G.; Stoops, B.; van Diepen, H.A.; Jongeneelen, M.; et al. A small-molecule fusion inhibitor of influenza virus is orally active in mice. Science 2019, 363, 6431. [Google Scholar] [CrossRef] [PubMed]
- Badani, H.; Garry, R.F.; Wilson, R.B.; Wimley, W.C. Mechanism and action of flufirvitide, a peptide inhibitor of influenza virus infection. Biophys. J. 2011, 100, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Badani, H.G.R.; Wimley, W.C. Peptide entry inhibitors of enveloped viruses: The importance of interfacial hydrophobicity. Biochim. Biophys. Acta Biomembr. 2014, 1838, 2180–2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, Y.; Li, J.; Niu, Y.; Liu, X.; Ren, L.; Guo, L.; Cheng, M.; Zhou, H.; Wang, J.; Jin, Q.; et al. Entry properties and entry inhibitors of a human H7N9 influenza virus. PLoS ONE 2014, 9, e107235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spevak, W.; Nagy, J.O.; Charych, D.H.; Schaefer, M.E.; Gilbert, J.H.; Bednarski, M.D. Polymerized liposomes containing C-glycosides of sialic acid: Potent inhibitors of influenza virus in vitro infectivity. J. Am. Chem. Soc. 1993, 115, 1146–1147. [Google Scholar] [CrossRef]
- Reuter, J.D.; Myc, A.; Hayes, M.M.; Gan, Z.; Roy, R.; Qin, D.; Yin, R.; Piehler, L.T.; Esfand, R.; Tomalia, D.A.; et al. Inhibition of viral adhesion and infection by sialic-acid-conjugated dendritic polymers. Bioconjug. Chem. 1999, 10, 271–278. [Google Scholar] [CrossRef]
- Lauster, D.; Klenk, S.; Ludwig, K.; Nojoumi, S.; Behren, S.; Adam, L.; Stadtmuller, M.; Saenger, S.; Zimmler, S.; Honzke, K.; et al. Phage capsid nanoparticles with defined ligand arrangement block influenza virus entry. Nat. Nanotechnol. 2020, 15, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, T.; Onishi, A.; Saito, T.; Shimada, A.; Inoue, H.; Taki, T.; Nagata, K.; Okahata, Y.; Sato, T. Sialic acid-mimic peptides as hemagglutinin inhibitors for anti-influenza therapy. J. Med. Chem. 2010, 53, 4441–4449. [Google Scholar] [CrossRef]
- Jeon, S.H.; Kayhan, B.; Ben-Yedidia, T.; Arnon, R. A DNA aptamer prevents influenza infection by blocking the receptor binding region of the viral hemagglutinin. J. Biol. Chem. 2004, 279, 48410–48419. [Google Scholar] [CrossRef] [Green Version]
- Gopinath, S.C.; Kumar, P.K. Aptamers that bind to the hemagglutinin of the recent pandemic influenza virus H1N1 and efficiently inhibit agglutination. Acta Biomater. 2013, 9, 8932–8941. [Google Scholar] [CrossRef]
- Petersen, E.; Keld, D.B.; Ellermann-Eriksen, S.; Gubbels, S.; Ilkjaer, S.; Jensen-Fangel, S.; Lindskov, C. Failure of combination oral oseltamivir and inhaled zanamivir antiviral treatment in ventilator- and ECMO-treated critically ill patients with pandemic influenza A (H1N1)v. Scand. J. Infect. Dis. 2011, 43, 495–503. [Google Scholar] [CrossRef]
- Duval, X.; van der Werf, S.; Blanchon, T.; Mosnier, A.; Bouscambert-Duchamp, M.; Tibi, A.; Enouf, V.; Charlois-Ou, C.; Vincent, C.; Andreoletti, L.; et al. Efficacy of oseltamivir-zanamivir combination compared to each monotherapy for seasonal influenza: A randomized placebo-controlled trial. PLoS Med. 2010, 7, e1000362. [Google Scholar] [CrossRef]
- de Mello, C.P.P.; Drusano, G.L.; Adams, J.R.; Shudt, M.; Kulawy, R.; Brown, A.N. Oseltamivir-zanamivir combination therapy suppresses drug-resistant H1N1 influenza A viruses in the hollow fiber infection model (HFIM) system. Eur. J. Pharm. Sci. 2018, 111, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Hung, I.F.; To, K.K.; Lee, C.K.; Lee, K.L.; Chan, K.; Yan, W.W.; Liu, R.; Watt, C.L.; Chan, W.M.; Lai, K.Y.; et al. Convalescent plasma treatment reduced mortality in patients with severe pandemic influenza A (H1N1) 2009 virus infection. Clin. Infect. Dis. 2011, 52, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Chung, F.T.; Lin, S.M.; Huang, S.Y.; Chou, C.L.; Lee, K.Y.; Lin, T.Y.; Kuo, H.P. Adjuvant treatment with a mammalian target of rapamycin inhibitor, sirolimus, and steroids improves outcomes in patients with severe H1N1 pneumonia and acute respiratory failure. Crit. Care Med. 2014, 42, 313–321. [Google Scholar] [CrossRef]
- Seo, S.; Englund, J.A.; Nguyen, J.T.; Pukrittayakamee, S.; Lindegardh, N.; Tarning, J.; Tambyah, P.A.; Renaud, C.; Went, G.T.; de Jong, M.D.; et al. Combination therapy with amantadine, oseltamivir and ribavirin for influenza A infection: Safety and pharmacokinetics. Antivir. Ther. 2013, 18, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.Y.; Young Suh, G.; Huh, J.W.; Kim, S.H.; Kim, M.J.; Kim, Y.S.; Kim, H.R.; Ryu, Y.J.; Han, M.S.; Ko, Y.G.; et al. Triple-combination antiviral drug for pandemic H1N1 influenza virus infection in critically ill patients on mechanical ventilation. Antimicrob. Agents Chemother. 2011, 55, 5703–5709. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Neuraminidase Inhibitor | Route | Therapeutic Regimen | Peak Time (Hours) | Half-Life (Hours) | Clearance | ||
|---|---|---|---|---|---|---|---|
| Frequency | Age/ Weight | Dose | |||||
| Zanamivir | Inhalation | twice daily for 5 days | ≥7 years old | 10 mg | 1–2 | 2 | Urine within 24 h or feces (unabsorbed) |
| Oseltamivir | Oral | twice daily for 5 days | ≤15 kg | 30 mg | 3–4 | 6–10 | Renal excretion |
| 16–23 kg | 45 mg | ||||||
| 24–40 kg | 60 mg | ||||||
| >40 kg | 75 mg | ||||||
| ≥13 years | 75 mg | ||||||
| Peramivir | Intravenous | Once daily for 5 to 10 days | Adult | 600 mg | 2–4 | 7.7–20.8 | Renal excretion (90%) |
| Laninamivir | Inhalation | Once | ≤10 years | 20 mg | 4 | 67 | Renal excretion (34%) |
| ≥10 years | 40 mg | ||||||
| Influenza Type/Subtype | Neuraminidase Inhibitor | Common Substitution in Neuraminidase (N2 Numbering) | References |
|---|---|---|---|
| A(H1N1)pdm09 | Zanamivir | E119D/G and S246R | [23,25,26,27,29,30,31,32,33,34,35,36] |
| Oseltamivir | E119D, I222R, S246G/R and H274Y | ||
| Peramivir | E119D/G, S246R and H274Y | ||
| Laninamivir | E119D/G and S246R | ||
| A(H5N1) | Zanamivir | None | [23,37,38] |
| Oseltamivir | H274Y and N294S | ||
| Peramivir | H274Y | ||
| Laninamivir | None | ||
| A(H3N2) | Zanamivir | E119I | [12,39,40,41,42,43,44,45] |
| Oseltamivir | E119I/V, R292K and N294S | ||
| Peramivir | R292K | ||
| Laninamivir | None | ||
| B | Zanamivir | R152K, I222L and G404S | [26,36,41,46,47,48,49,50,51,52] |
| Oseltamivir | R152K, D198E, I222L and N294S | ||
| Peramivir | E107K, R152K, D198E, I222L/T and N294S | ||
| Laninamivir | R152K |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, Y.; Jones, J.C.; Wong, S.-S.; Zanin, M. Antivirals Targeting the Surface Glycoproteins of Influenza Virus: Mechanisms of Action and Resistance. Viruses 2021, 13, 624. https://doi.org/10.3390/v13040624
Bai Y, Jones JC, Wong S-S, Zanin M. Antivirals Targeting the Surface Glycoproteins of Influenza Virus: Mechanisms of Action and Resistance. Viruses. 2021; 13(4):624. https://doi.org/10.3390/v13040624
Chicago/Turabian StyleBai, Yaqin, Jeremy C. Jones, Sook-San Wong, and Mark Zanin. 2021. "Antivirals Targeting the Surface Glycoproteins of Influenza Virus: Mechanisms of Action and Resistance" Viruses 13, no. 4: 624. https://doi.org/10.3390/v13040624
APA StyleBai, Y., Jones, J. C., Wong, S. -S., & Zanin, M. (2021). Antivirals Targeting the Surface Glycoproteins of Influenza Virus: Mechanisms of Action and Resistance. Viruses, 13(4), 624. https://doi.org/10.3390/v13040624