
Characterization of a Novel Group of Listeria Phages That Target Serotype 4b Listeria monocytogenes



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Bacteriophages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Serotype | Reference or Original |
|---|---|---|
| 10403S | 1/2a | Bishop and Hinrichs, 1987 [32] |
| MACK | 1/2a | Hodgson, 2000 [33] |
| F2365 | 4b | Nelson, 2004 [34] |
| FSL J1-175 | 1/2b | Bergholz, 2010 [35] |
| FSL J1-208 | 4a | Roberts, 2006 [36] |
| FSL C1-115 | 3a | Fugett, 2006 [37] |
| FSL J1-094 | 1/2c | Fugett, 2006 [37] |
| FSL F2-695 | 4a | Roberts, 2006 [36] |
| FSL F2-501 | 4b | Roberts, 2006 [36] |
| FSL J2-071 | 4c | Roberts, 2006 [36] |
| FSL W1-110 | 4b | De Jesus and Whiting, 2003 [38] |
| FSL J1-158 | 4b | De Jesus and Whiting, 2003 [38] |
| FSL J1-169 | 3b | Fugett, 2006 [37] |
| FSL J1-049 | 3c | Fugett, 2006 [37] |
| FSL D4-0014 | 1/2a | Denes, 2015 [15] |
| FSL D4-0119 | 3 | Denes, 2015 [15] |
| FSL R9-0915 | 7 | Denes, 2015 [15] |
2.2. Transmission Electron Microscopy
2.3. DNA Extraction and Genomic Analysis
2.4. Efficiencies of Plaquing and Relative Phage Activity
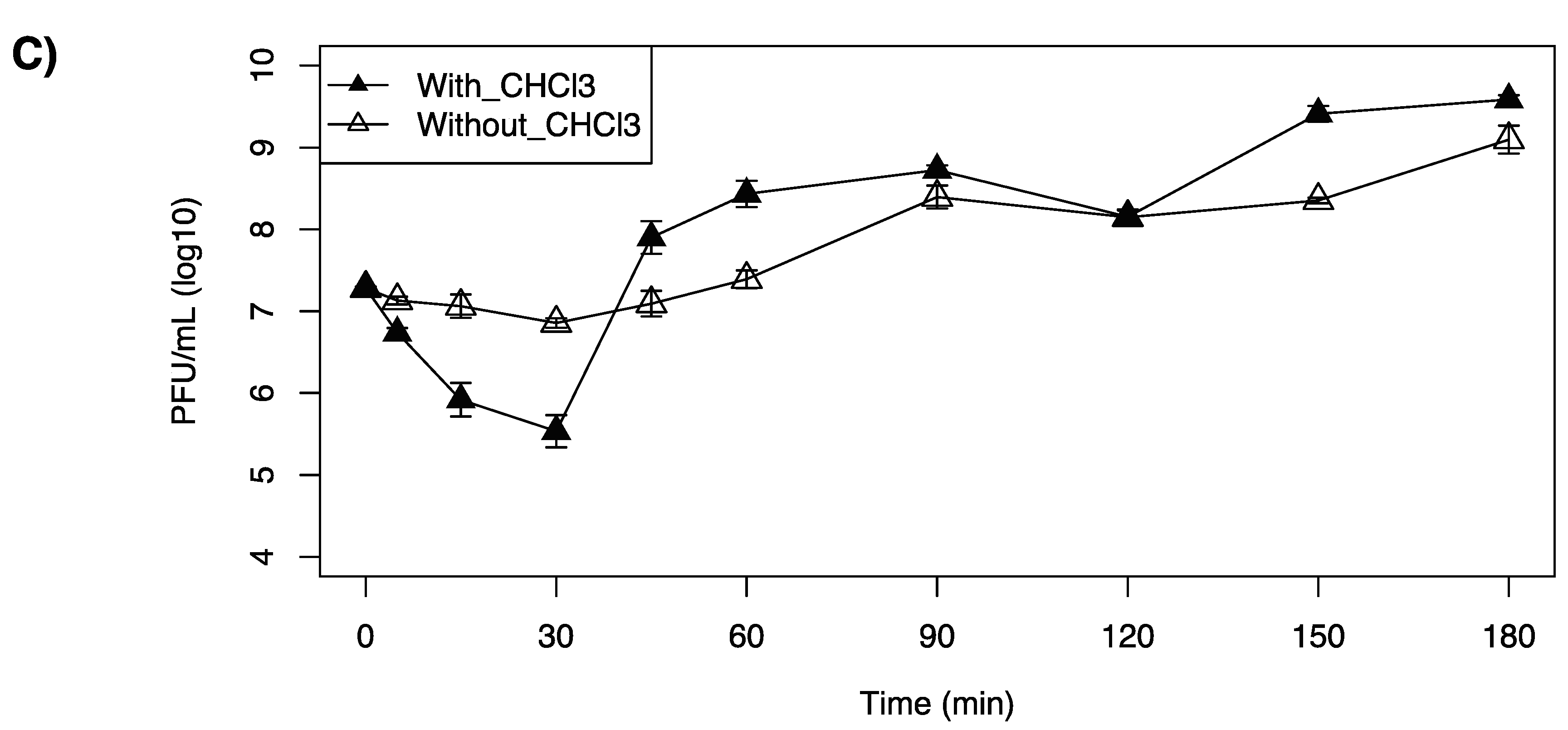
2.5. One-Step Growth Curve
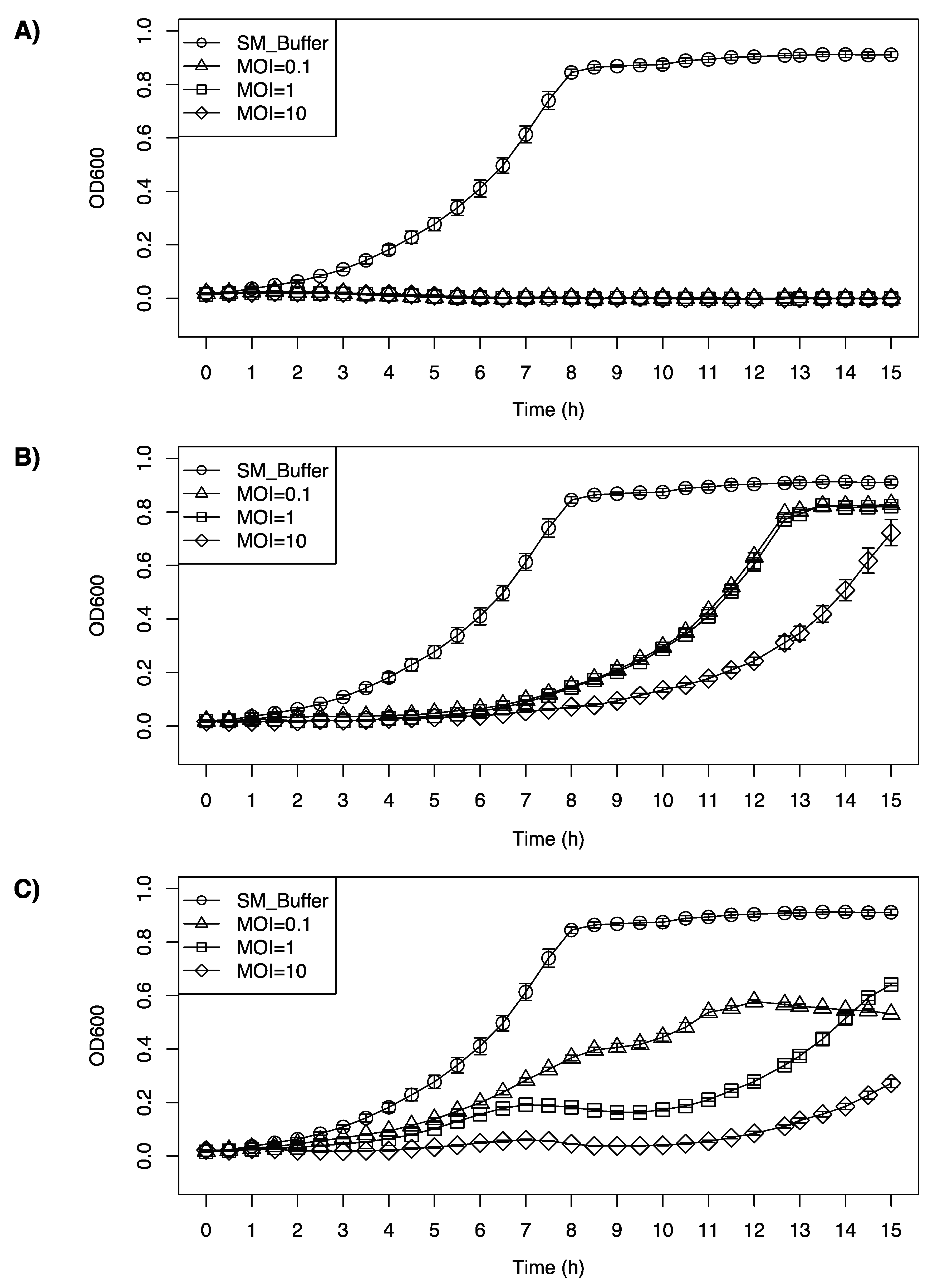
2.6. Inhibition Growth Curve of Listeria monocytogenes F2365 by LP-020, LP-027, and LP-094
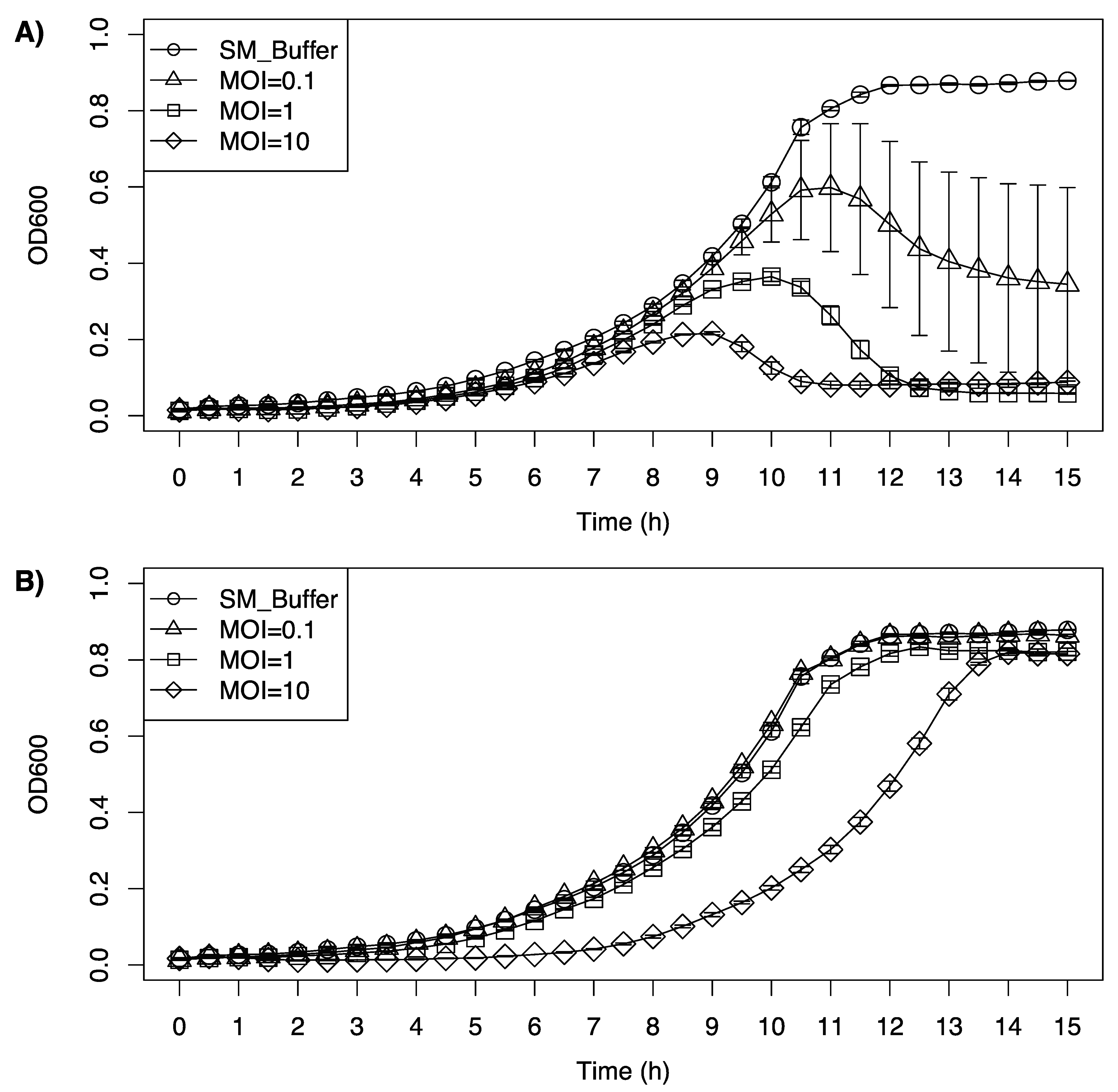
2.7. Inhibition Growth Curve of Listeria monocytogenes Cocktail by LP-020 and LP-094
3. Results and Discussion
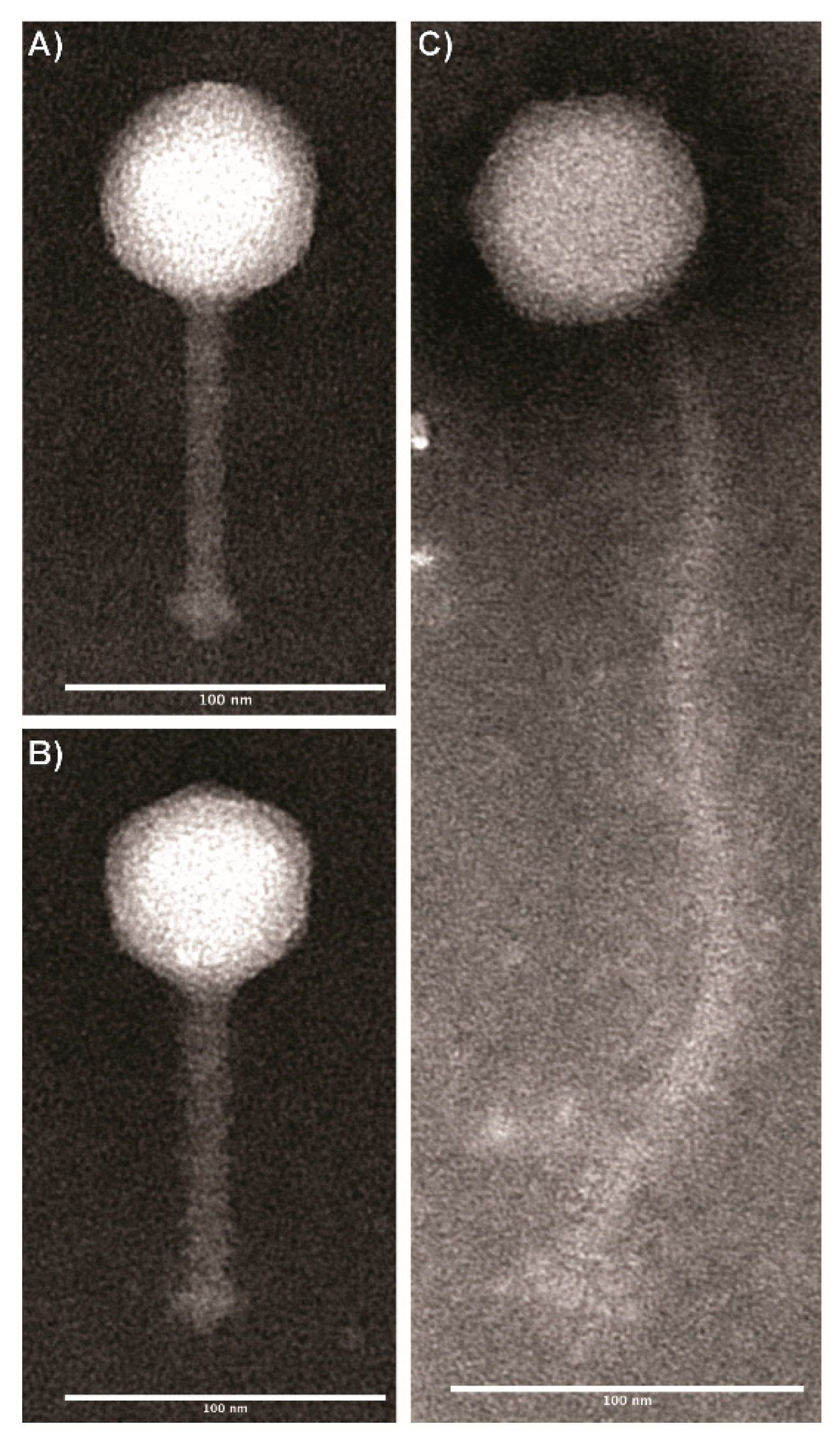
3.1. Transmission Electron Microscopy Imaging of Wild Type Listeria Phages Revealed Two Distinct Morphologies
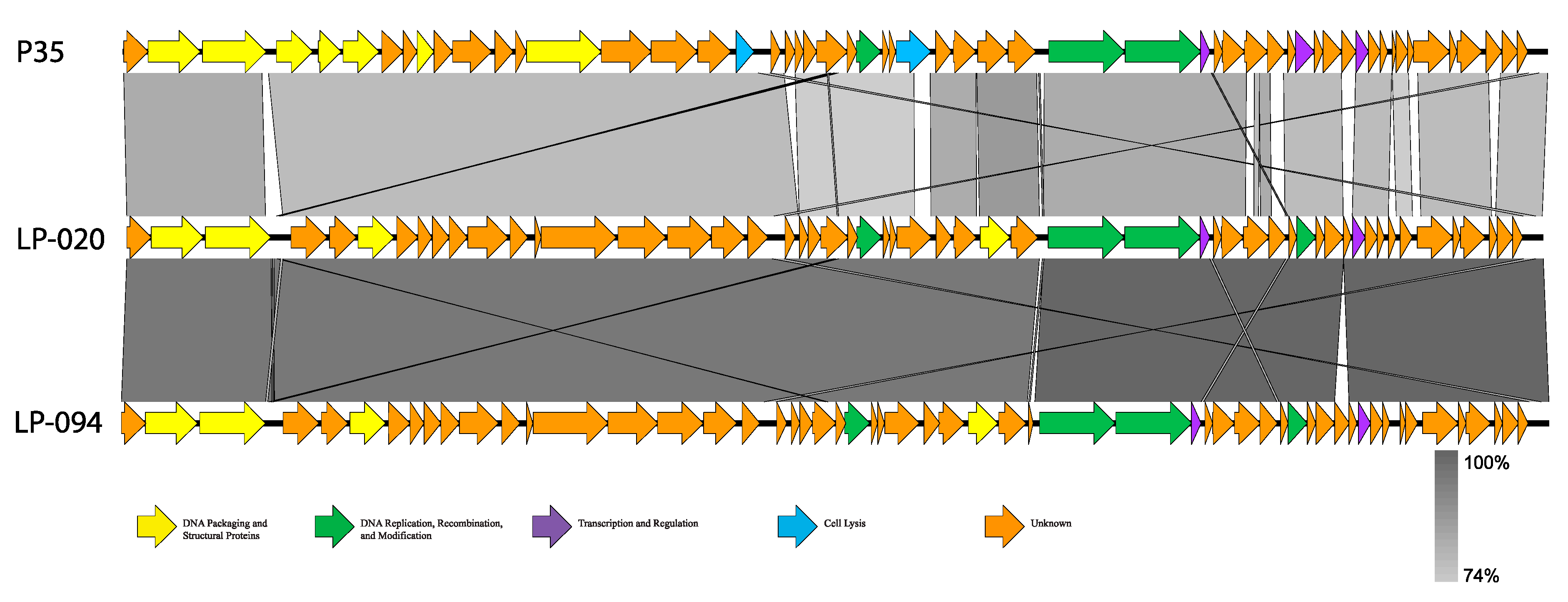
3.2. Genomic Analysis
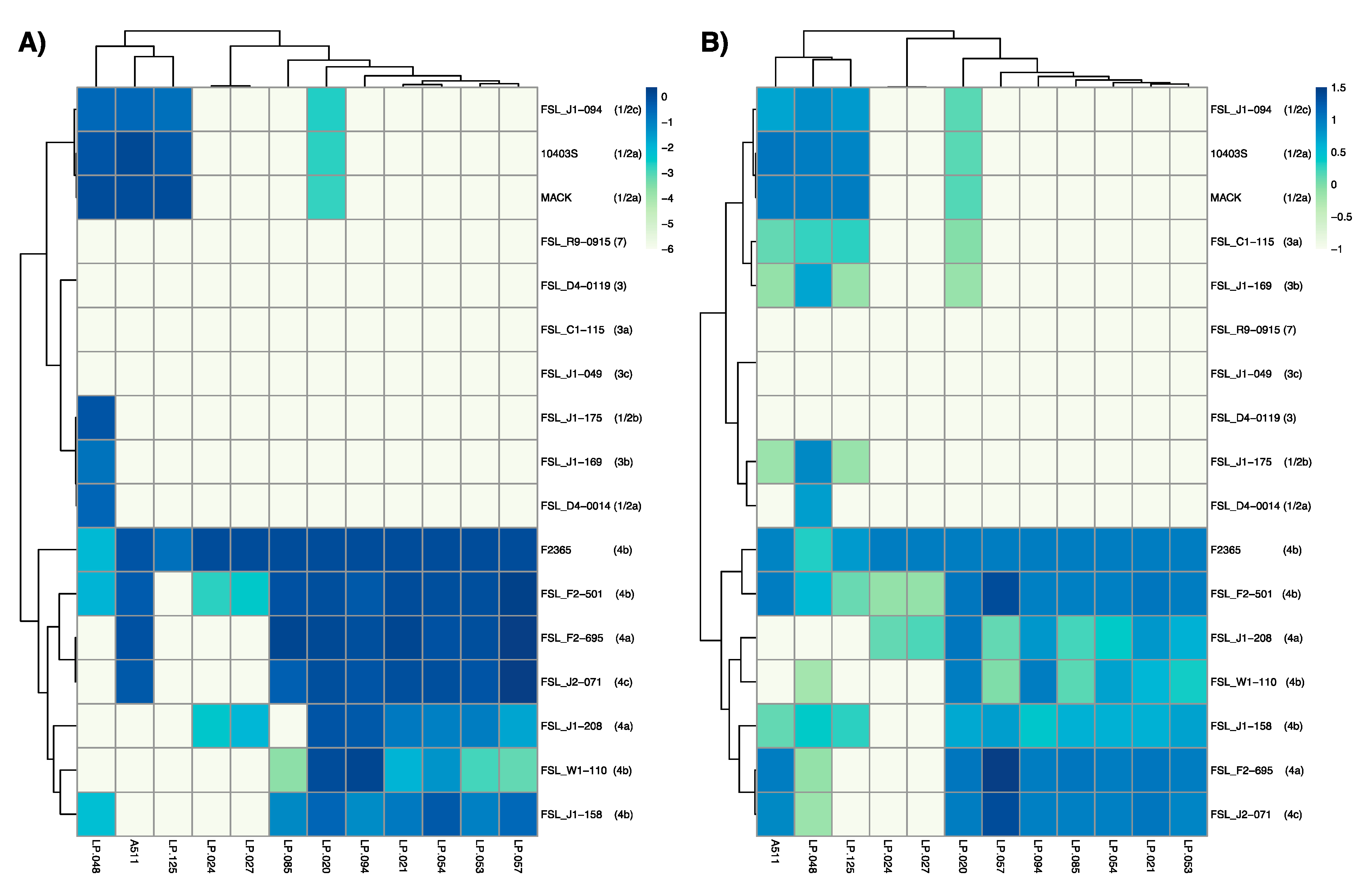
3.3. Host Range Analysis
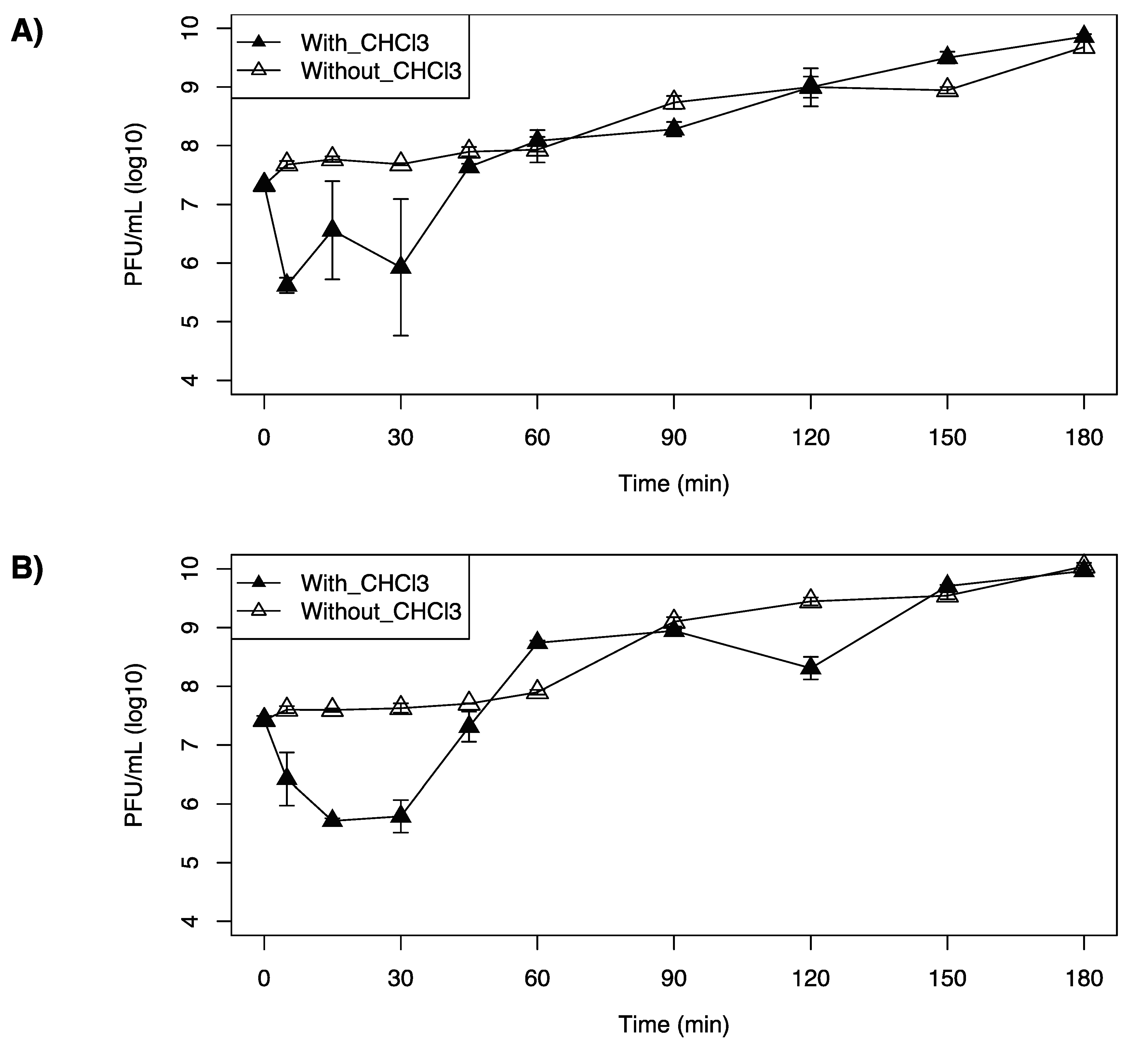
3.4. One-Step Growth Curves
3.5. Inhibition Growth Curve of Listeria monocytogenes F2365 by LP-020, LP-027, and LP-094
3.6. Inhibition Growth Curve of Listeria monocytogenes Cocktail by LP-020 and LP-094
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Karthikeyan, R.; Gayathri, P.; Gunasekaran, P.; Jagannadham, M.V.; Rajendhran, J. Comprehensive proteomic analysis and pathogenic role of membrane vesicles of Listeria monocytogenes serotype 4b reveals proteins associated with virulence and their possible interaction with host. Int. J. Med. Microbiol. 2019, 309, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Vivant, A.L.; Garmyn, D.; Piveteau, P. Listeria monocytogenes, a down-to-earth pathogen. Front. Cell. Infect. Microbiol. 2013, 3, 87. [Google Scholar] [CrossRef] [Green Version]
- Sauders, B.D.; Overdevest, J.; Fortes, E.; Windham, K.; Schukken, Y.; Lembo, A.; Wiedmann, M. Diversity of Listeria Species in Urban and Natural Environments. Appl. Environ. Microbiol. 2012, 78, 4420–4433. [Google Scholar] [CrossRef] [Green Version]
- Fenlon, D. Wild birds and silage as reservoirs of Listeria in the agricultural environment. J. Appl. Bacteriol. 1985, 59, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Lawrence, M.L.; Ainsworth, A.J.; Austin, F.W. Comparative assessment of acid, alkali and salt tolerance in Listeria monocytogenes virulent and avirulent strains. FEMS Microbiol. Lett. 2005, 243, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Malakar, D.; Borah, P.; Das, L.; Kumar, N.S. A Comprehensive Review on Molecular Characteristics and Food-Borne Outbreaks of Listeria Monocytogenes. Sci. Technol. J. 2019, 7. [Google Scholar] [CrossRef]
- De Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7. [Google Scholar] [CrossRef]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607. [Google Scholar] [CrossRef] [PubMed]
- Cost Estimates of Foodborne Illnesses. 2021. Available online: https://www.ers.usda.gov/data-products/cost-estimates-of-foodborne-illnesses.aspx (accessed on 4 March 2021).
- Carlton, R.; Noordman, W.; Biswas, B.; de Meester, E.; Loessner, M. Bacteriophage P100 for control of Listeria monocytogenes in foods: Genome sequence, bioinformatic analyses, oral toxicity study, and application. Regul. Toxicol. Pharmacol. 2005, 43, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Jurczak-Kurek, A.; Gąsior, T.; Nejman-Faleńczyk, B.; Bloch, S.; Dydecka, A.; Topka, G.; Necel, A.; Jakubowska-Deredas, M.; Narajczyk, M.; Richert, M.; et al. Biodiversity of bacteriophages: Morphological and biological properties of a large group of phages isolated from urban sewage. Sci. Rep. 2016, 6, 34338. [Google Scholar] [CrossRef]
- Sillankorva, S.M.; Oliveira, H.; Azeredo, J. Bacteriophages and Their Role in Food Safety. Int. J. Microbiol. 2012, 2012, 863945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibeu, A.; Agius, L.; Gao, A.; Sabour, P.M.; Kropinski, A.M.; Balamurugan, S. Efficacy of bacteriophage LISTEX™P100 combined with chemical antimicrobials in reducing Listeria monocytogenes in cooked turkey and roast beef. Int. J. Food Microbiol. 2013, 167, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Denes, T.; Bakker, H.C.D.; Tokman, J.I.; Guldimann, C.; Wiedmann, M. Selection and Characterization of Phage-Resistant Mutant Strains of Listeria monocytogenes Reveal Host Genes Linked to Phage Adsorption. Appl. Environ. Microbiol. 2015, 81, 4295–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumrall, E.T.; Keller, A.P.; Shen, Y.; Loessner, M.J. Structure and function of Listeria teichoic acids and their implications. Mol. Microbiol. 2020, 113, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Vongkamjan, K.; Switt, A.M.; Bakker, H.C.D.; Fortes, E.D.; Wiedmann, M. Silage Collected from Dairy Farms Harbors an Abundance of Listeriaphages with Considerable Host Range and Genome Size Diversity. Appl. Environ. Microbiol. 2012, 78, 8666–8675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasebe, R.; Nakao, R.; Ohnuma, A.; Yamasaki, T.; Sawa, H.; Takai, S.; Horiuchi, M. Listeria monocytogenes serotype 4b strains replicate in monocytes/macrophages more than the other serotypes. J. Vet. Med. Sci. 2017, 79, 962–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontello, M.; Guaita, A.; Sala, G.; Cipolla, M.; Gattuso, A.; Sonnessa, M.; Gianfranceschi, M.V. Listeria monocytogenes serotypes in human infections (Italy, 2000–2010). Ann. dell’Istituto Super. Sanità 2012, 48, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Aarnisalo, K.; Autio, T.; Sjöberg, A.M.; Lundén, J.; Korkeala, H.; Suihko, M.L. Typing of Listeria monocytogenes isolates originating from the food processing industry with automated ribotyping and pulsed-field gel electrophoresis. J. Food Prot. 2003, 66, 249–255. [Google Scholar] [CrossRef]
- Liu, D. Handbook of Listeria Monocytogenes; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Huang, Y.; Morvay, A.A.; Shi, X.; Suo, Y.; Shi, C.; Knøchel, S. Comparison of oxidative stress response and biofilm formation of Listeria monocytogenes serotypes 4b and 1/2a. Food Control 2018, 85, 416–422. [Google Scholar] [CrossRef]
- Nucera, D.; Lomonaco, S.; Bianchi, D.M.; Decastelli, L.; Grassi, M.A.; Bottero, M.T.; Civera, T. A five year surveillance report on PFGE types of Listeria monocytogenes isolated in Italy from food and food related environments. Int. J. Food Microbiol. 2010, 140, 271–276. [Google Scholar] [CrossRef]
- Leong, D.; Alvarez-Ordóñez, A.; Jordan, K. Monitoring occurrence and persistence of Listeria monocytogenes in foods and food processing environments in the Republic of Ireland. Front. Microbiol. 2014, 5, 436. [Google Scholar] [CrossRef]
- Peters, T.L.; Song, Y.; Bryan, D.W.; Hudson, L.K.; Denes, T.G. Mutant and Recombinant Phages Selected from In Vitro Coevolution Conditions Overcome Phage-Resistant Listeria monocytogenes. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Song, Y.; Peters, T.L.; Bryan, D.W.; Hudson, L.K.; Denes, T.G. Homburgvirus LP-018 Has a Unique Ability to Infect Phage-Resistant Listeria monocytogenes. Viruses 2019, 11, 1166. [Google Scholar] [CrossRef] [Green Version]
- Tokman, J.I.; Kent, D.J.; Wiedmann, M.; Denes, T. Temperature Significantly Affects the Plaquing and Adsorption Efficiencies of Listeria Phages. Front. Microbiol. 2016, 7, 631. [Google Scholar] [CrossRef]
- Kim, J.-W.; Kathariou, S. Temperature-Dependent Phage Resistance of Listeria monocytogenes Epidemic Clone II. Appl. Environ. Microbiol. 2009, 75, 2433–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, H.-W.; Tremblay, D.; Moineau, S. Long-Term Bacteriophage Preservation. 2004. Available online: https://www.researchgate.net/profile/Sylvain-Moineau/publication/285783875_Long-term_bacteriophage_preservation/links/56965e4b08ae34f3cf1dbb34/Long-term-bacteriophage-preservation.pdf (accessed on 12 April 2021).
- Clark, W.A. Comparison of several methods for preserving bacteriophages. Appl. Microbiol. 1962, 10, 466–471. [Google Scholar] [CrossRef]
- Cooper, C.; Denyer, S.; Maillard, J.-Y. Stability and purity of a bacteriophage cocktail preparation for nebulizer delivery. Lett. Appl. Microbiol. 2013, 58, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.; Hinrichs, D. Adoptive transfer of immunity to Listeria monocytogenes. The influence of in vitro stimulation on lymphocyte subset requirements. J. Immunol. 1987, 139, 2005–2009. [Google Scholar] [PubMed]
- Hodgson, D.A. Generalized transduction of serotype 1/2 and serotype 4b strains of Listeria monocytogenes. Mol. Microbiol. 2000, 35, 312–323. [Google Scholar] [CrossRef]
- Nelson, K.E.; Fouts, D.E.; Mongodin, E.F.; Ravel, J.; DeBoy, R.T.; Kolonay, J.F.; Rasko, D.A.; Angiuoli, S.V.; Gill, S.R.; Paulsen, I.T.; et al. Whole genome comparisons of serotype 4b and 1/2a strains of the food-borne pathogen Listeria monocytogenes reveal new insights into the core genome components of this species. Nucleic Acids Res. 2004, 32, 2386–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergholz, T.M.; Bakker, H.C.D.; Fortes, E.D.; Boor, K.J.; Wiedmann, M. Salt Stress Phenotypes in Listeria monocytogenes Vary by Genetic Lineage and Temperature. Foodborne Pathog. Dis. 2010, 7, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Nightingale, K.; Jeffers, G.; Fortes, E.; Kongo, J.M.; Wiedmann, M. Genetic and phenotypic characterization of Listeria monocytogenes lineage III. Microbiology 2006, 152, 685–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fugett, E.; Fortes, E.; Nnoka, C.; Wiedmann, M. International Life Sciences Institute North America Listeria monocytogenes strain collection: Development of standard Listeria monocytogenes strain sets for research and validation studies. J. Food Prot. 2006, 69, 2929–2938. [Google Scholar] [CrossRef]
- De Jesus, A.J.; Whiting, R.C. Thermal inactivation, growth, and survival studies of Listeria monocytogenes strains belonging to three distinct genotypic lineages. J. Food Prot. 2003, 66, 1611–1617. [Google Scholar] [CrossRef]
- Loessner, M.J.; Busse, M. Bacteriophage typing of Listeria species. Appl. Environ. Microbiol. 1990, 56, 1912–1918. [Google Scholar] [CrossRef] [Green Version]
- Klumpp, J.; Dorscht, J.; Lurz, R.; Bielmann, R.; Wieland, M.; Zimmer, M.; Calendar, R.; Loessner, M.J. The Terminally Redundant, Nonpermuted Genome of Listeria Bacteriophage A511: A Model for the SPO1-Like Myoviruses of Gram-Positive Bacteria. J. Bacteriol. 2008, 190, 5753–5765. [Google Scholar] [CrossRef] [Green Version]
- Denes, T.; Vongkamjan, K.; Ackermann, H.-W.; Switt, A.I.M.; Wiedmann, M.; Bakker, H.C.D. Comparative Genomic and Morphological Analyses of Listeria Phages Isolated from Farm Environments. Appl. Environ. Microbiol. 2014, 80, 4616–4625. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with multiplex MinION sequencing. Microb. Genom. 2017, 3, e000132. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Bushnell, B. BBTools: A Suite of Fast, Multithreaded Bioinformatics Tools Designed for Analysis of DNA and RNA Sequence Data. Joint Genome Institute. 2018. Available online: https://jgi.doe.gov/data-and-tools/bbtools (accessed on 29 December 2020).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2015, 32, 929–931. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- McNair, K.; Bailey, B.A.; Edwards, R.A. PHACTS, a computational approach to classifying the lifestyle of phages. Bioinformatics 2012, 28, 614–618. [Google Scholar] [CrossRef] [Green Version]
- Trudelle, D.M.; Bryan, D.W.; Hudson, L.K.; Denes, T.G. Cross-resistance to phage infection in Listeria monocytogenes serotype 1/2a mutants. Food Microbiol. 2019, 84, 103239. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps (Version 1.0.12). 2019. Available online: https://rdrr.io/cran/pheatmap/ (accessed on 1 October 2020).
- Dorscht, J.; Klumpp, J.; Bielmann, R.; Schmelcher, M.; Born, Y.; Zimmer, M.; Calendar, R.; Loessner, M.J. Comparative Genome Analysis of Listeria Bacteriophages Reveals Extensive Mosaicism, Programmed Translational Frameshifting, and a Novel Prophage Insertion Site. J. Bacteriol. 2009, 191, 7206–7215. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, E.M.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, M.D.; Klumpp, J.; Barrios, M.E.; Mbayed, V.A. Gene gain and loss and recombination shape evolution of Listeria bacteriophages of the genus Pecentumvirus. Genomics 2020, 113, 411–419. [Google Scholar] [CrossRef]
- Peters, T.L.; Hudson, L.K.; Song, Y.; Denes, T.G. Complete Genome Sequences of Two Listeria Phages of the Genus Pecentumvirus. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, L.K.; Peters, T.L.; Song, Y.; Denes, T.G. Complete Genome Sequences and Transmission Electron Micrographs of Listeria Phages of the Genus Homburgvirus. Microbiol. Resour. Announc. 2019, 8, e00825-19. [Google Scholar] [CrossRef] [Green Version]
- Schmuki, M.M.; Erne, D.; Loessner, M.J.; Klumpp, J. Bacteriophage P70: Unique Morphology and Unrelatedness to Other Listeria Bacteriophages. J. Virol. 2012, 86, 13099–13102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilcher, S.; Studer, P.; Muessner, C.; Klumpp, J.; Loessner, M.J. Cross-genus rebooting of custommade, synthetic bacteriophage genomes in L-form bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, 567–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]







| Phages | Description | Reference or Original |
|---|---|---|
| A511 | Loessner and Busse, 1990 [39]; Klumpp, Jochen, et al., 2008 [40] | |
| LP-020 | Vongkamjan et al., 2012 | |
| LP-021 | Vongkamjan et al., 2012 [17] | |
| LP-024 | Vongkamjan et al., 2012 [17] | |
| LP-027 | Vongkamjan et al., 2012 [17] | |
| LP-048 | P100-like phage | Vongkamjan et al., 2012; Denes et al., 2014 [17,41] |
| LP-053 | Vongkamjan et al., 2012 [17,41] | |
| LP-054 | Vongkamjan et al., 2012 [17] | |
| LP-057 | Vongkamjan et al., 2012 [17] | |
| LP-085 | Vongkamjan et al., 2012 [17] | |
| LP-094 | Vongkamjan et al., 2012 [17] | |
| LP-125 | P100-like phage | Vongkamjan et al., 2012; Denes et al., 2014 [17,41] |
| Listeria monocytogenes Phages | Capsid Diameter (nm) | Tail Length (nm) | Tail Width (nm) |
|---|---|---|---|
| LP-020 | 73.40 ± 0.61 | 104.80 ± 1.45 | 11.54 ± 0.09 |
| LP-021 | 69.92 ± 4.62 | 102.45 ± 2.32 | 11.01 ± 0.61 |
| LP-024 | 66.10 ± 5.07 | 291.29 ± 18.56 | 8.25 ± 2.54 |
| LP-027 | 70.35 ± 2.40 | 295.38 ± 10.42 | 7.95 ± 2.09 |
| LP-053 | 68.03 ± 4.76 | 104.01 ± 3.00 | 11.88 ± 1.91 |
| LP-054 | 73.70 ± 0.50 | 100.71 ± 3.55 | 11.43 ± 0.33 |
| LP-057 | 73.60 ± 0.37 | 102.45 ± 1.62 | 13.46 ± 0.90 |
| LP-085 | 72.22 ± 0.62 | 104.02 ± 3.19 | 12.61 ± 0.77 |
| LP-094 | 71.75 ± 2.98 | 100.49 ± 2.87 | 12.81 ± 0.97 |
| Phage | BioSample ID | Length (bp) | Avg. Illumina Read Coverage (X) | G+C (%) | No. CDS | No. RNAs |
|---|---|---|---|---|---|---|
| LP-020 | SAMN17217625 | 35,609 | 326.6 | 40.0 | 54 | 0 |
| LP-021 | SAMN17217626 | 35,610 | 344.1 | 40.0 | 54 | 0 |
| LP-024 | SAMN17217627 | 40,964 | 153.7 | 36.5 | 74 | 0 |
| LP-027 | SAMN17217628 | 41,120 | 89.6 | 36.6 | 74 | 0 |
| LP-053 | SAMN17217629 | 35,951 | 143.5 | 40.0 | 57 | 0 |
| LP-054 | SAMN17217630 | 35,951 | 524.9 | 40.0 | 57 | 0 |
| LP-057 | SAMN17217631 | 35,608 | 1001.1 | 40.0 | 54 | 0 |
| LP-085 | SAMN17217632 | 35,951 | 650.4 | 39.9 | 57 | 0 |
| LP-094 | SAMN17217633 | 35,885 | 3397.6 | 40.0 | 56 | 0 |
| Average Nucleotide Identity (ANI; %) [Aligned Nucleotides (%)] | ||||||
|---|---|---|---|---|---|---|
| Phage | LP-024 | LP-027 | LP-030-3 | A500 | A118 | A006 |
| LP-024 | 100.00 [96.63] | 100.00 [96.63] | 92.81 [63.99] | 86.34 [52.10] | 92.00 [6.82] | |
| LP-027 | 100.00 [95.51] | 100.00 [95.92] | 92.77 [63.41] | 86.10 [50.80] | 91.76 [5.70] | |
| LP-030-3 | 99.99 [96.21] | 100.00 [96.57] | 92.07 [62.93] | 84.54 [47.14] | 88.41 [6.80] | |
| A500 | 92.74 [68.96] | 92.74 [68.96] | 92.74 [69.01] | 84.52 [49.43] | 89.93 [6.42] | |
| A118 | 84.79 [53.25] | 84.78 [53.48] | 84.78 [53.51] | 84.00 [47.63] | 95.38 [23.01] | |
| A006 | 89.85 [7.30] | 89.54 [9.18] | 89.54 [9.18] | 84.93 [4.69] | 94.91 [27.18] | |
| Average Nucleotide Identity (ANI; %) [Aligned Nucleotides (%)] | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Phage | LP-020 | LP-021 | LP-053 | LP-054 | LP-057 | LP-085 | LP-094 | P35 | P40 |
| LP-020 | 99.99 [97.36] | 97.68 [94.41] | 97.68 [94.41] | 98.71 [97.35] | 97.86 [94.41] | 97.66 [93.39] | 79.51 [85.61] | 61.76 [32.56] | |
| LP-021 | 99.99 [97.36] | 97.69 [94.41] | 97.69 [94.41] | 98.72 [97.34] | 97.87 [94.41] | 97.68 [93.39] | 79.52 [85.63] | 61.76 [32.55] | |
| LP-053 | 97.77 [90.63] | 97.78 [90.63] | 100.00 [99.24] | 99.15 [90.64] | 99.74 [99.24] | 99.98 [98.21] | 79.85 [87.67] | 61.47 [33.98] | |
| LP-054 | 97.77 [90.63] | 97.78 [90.63] | 100.00 [99.24] | 99.15 [90.64] | 99.74 [99.24] | 99.98 [98.21] | 79.85 [87.67] | 61.47 [33.98] | |
| LP-057 | 98.71 [97.34] | 98.72 [97.35] | 98.97 [94.42] | 98.97 [94.42] | 99.02 [94.42] | 98.96 [93.40] | 79.62 [85.79] | 61.80 [32.41] | |
| LP-085 | 97.97 [90.63] | 97.98 [90.63] | 99.74 [99.24] | 99.74 [99.24] | 99.22 [90.64] | 99.73 [98.21] | 79.87 [87.67] | 61.51 [33.94] | |
| LP-094 | 97.75 [91.58] | 97.76 [91.58] | 99.98 [98.24] | 99.98 [98.24] | 99.08 [91.59] | 99.73 [98.24] | 79.98 [86.27] | 61.54 [32.97] | |
| P35 | 79.85 [83.06] | 79.86 [83.07] | 80.35 [84.12] | 80.35 [84.12] | 80.02 [83.03] | 80.36 [84.12] | 80.33 [84.12] | 63.10 [23.90] | |
| P40 | 61.52 [25.48] | 61.81 [24.84] | 62.26 [22.30] | 62.26 [22.30] | 62.37 [22.14] | 62.36 [22.26] | 62.29 [22.30] | 62.80 [24.06] | |
| Listeria monocytogenes Phages | LP-020 | LP-027 | LP-094 |
|---|---|---|---|
| Adsorption Time(min) | 5 | 30 | 30 |
| Adsorption Rate(%) | 97.9 ± 0.5 | 96.4 ± 2.3 | 97.8 ± 1.0 |
| Latent Period(min) | 60~90 | 45~60 | 30~45 |
| Eclipse Period(min) | 5~15 | 30~45 | 30~45 |
| Burst Size(PFU/cell) | 9.7 ± 2.9 | 34.4 ± 5.8 | 28.3 ± 4.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Peters, T.L.; Bryan, D.W.; Hudson, L.K.; Denes, T.G. Characterization of a Novel Group of Listeria Phages That Target Serotype 4b Listeria monocytogenes. Viruses 2021, 13, 671. https://doi.org/10.3390/v13040671
Song Y, Peters TL, Bryan DW, Hudson LK, Denes TG. Characterization of a Novel Group of Listeria Phages That Target Serotype 4b Listeria monocytogenes. Viruses. 2021; 13(4):671. https://doi.org/10.3390/v13040671
Chicago/Turabian StyleSong, Yaxiong, Tracey L. Peters, Daniel W. Bryan, Lauren K. Hudson, and Thomas G. Denes. 2021. "Characterization of a Novel Group of Listeria Phages That Target Serotype 4b Listeria monocytogenes" Viruses 13, no. 4: 671. https://doi.org/10.3390/v13040671
APA StyleSong, Y., Peters, T. L., Bryan, D. W., Hudson, L. K., & Denes, T. G. (2021). Characterization of a Novel Group of Listeria Phages That Target Serotype 4b Listeria monocytogenes. Viruses, 13(4), 671. https://doi.org/10.3390/v13040671