
Changes in Subcellular Localization of Host Proteins Induced by Plant Viruses



Abstract
:1. Introduction
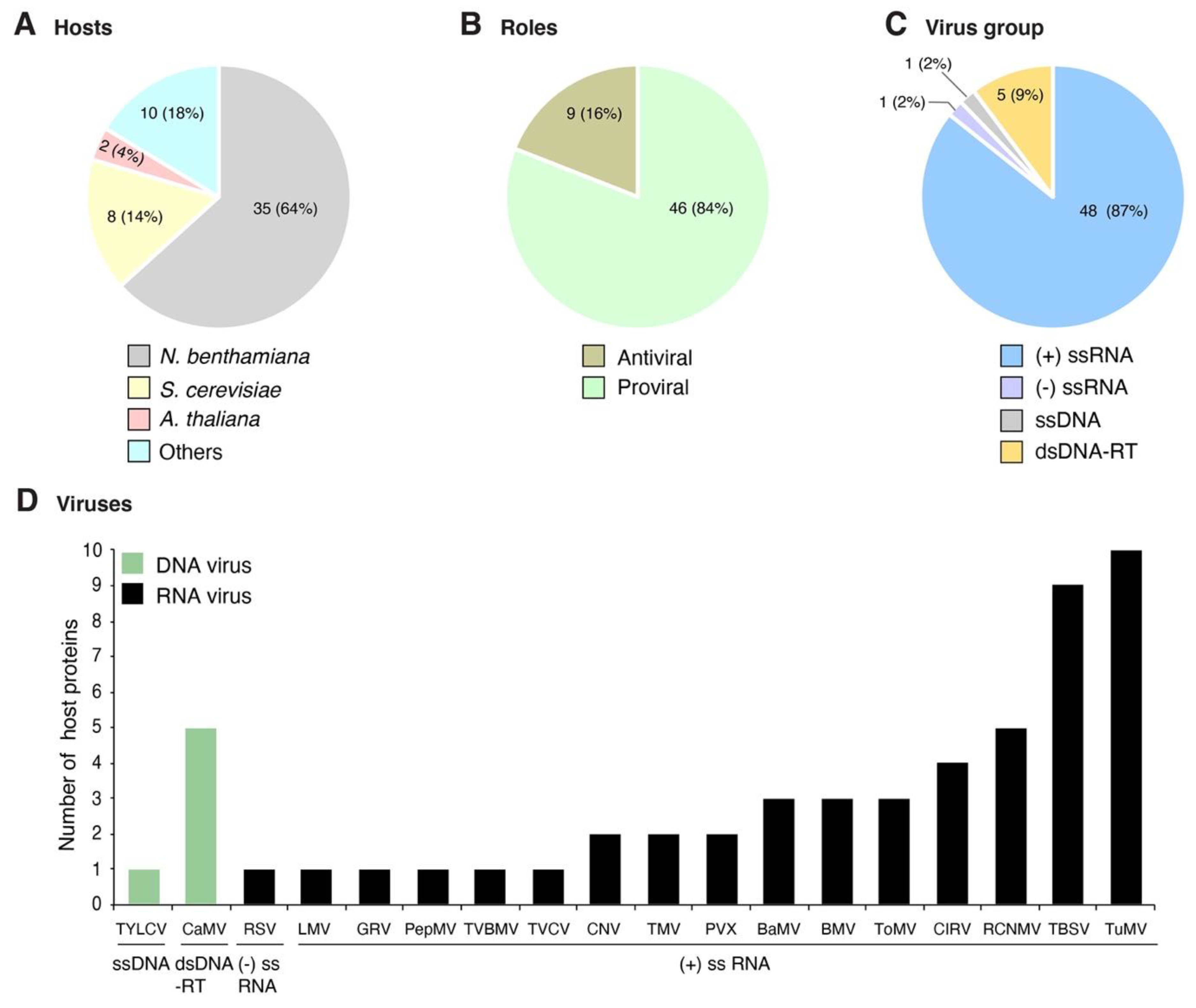
2. Profile of Host Proteins
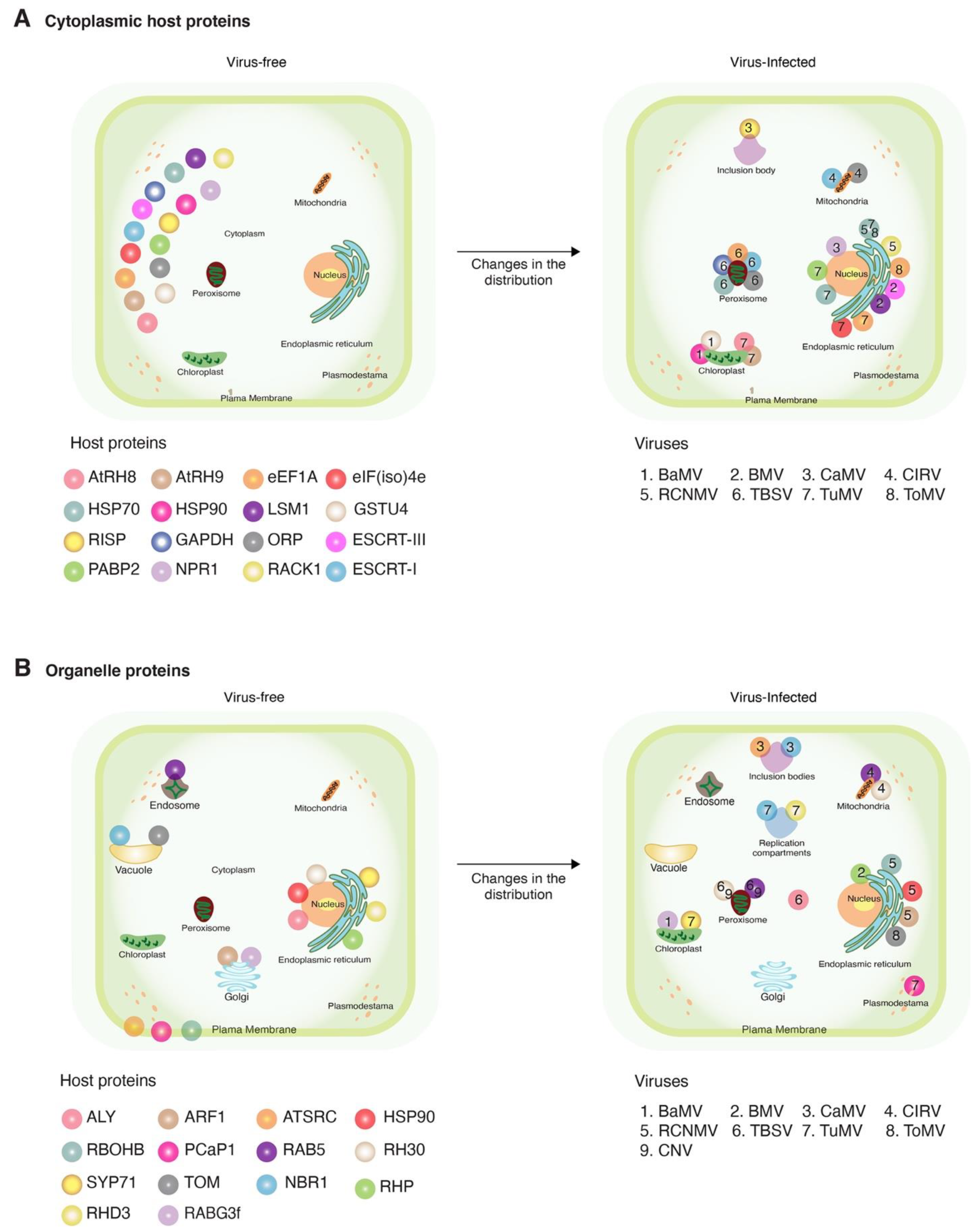
3. Cytoplasmic Host Proteins
3.1. Heat Shock Proteins (HSPs)
3.2. Endosomal Sorting Complexes Required for Transport (ESCRTs)
3.3. Translation Factors
3.4. Asp-Glu-Ala-Asp (DEAD)-Box RNA Helicases (RHAs)
3.5. Glyceraldehyde 3-Phosphate Dehydrogenase (GAPDH)
3.6. Glutathione Transferase U4 (GSTU4)
3.7. Other Cytoplasmic Proteins
4. Endosomal Proteins
5. Endoplasmic Reticulum Proteins
5.1. Soluble N-Ethyl Maleimide Sensitive Factor Adaptor Protein Receptors (SNAREs)
5.2. Reticulon Homology Domain Proteins (RHPs)
5.3. Root Hair Defective 3 (RHD3)
6. Golgi Apparatus Proteins
7. Plasma Membrane Proteins
8. Nuclear Proteins
9. Vacuolar Proteins
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Den Boon, J.A.; Diaz, A.; Ahlquist, P. Cytoplasmic viral replication complexes. Cell Host Microbe 2010, 8, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Ceniceros-Ojeda, E.A.; Rodriguez-Negrete, E.A.; Rivera-Bustamante, R.F. Two populations of viral minichromosomes are present in a geminivirus-infected plant showing symptom remission (recovery). J. Virol. 2016, 90, 3828–3838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ruiz, H. Susceptibility genes to plant viruses. Viruses 2018, 10, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, K.I.; Eskelin, K.; Lohmus, A.; Makinen, K. Molecular and cellular mechanisms underlying potyvirus infection. J. Gen. Virol. 2014, 95, 1415–1429. [Google Scholar] [CrossRef] [Green Version]
- Lellis, A.D.; Kasschau, K.D.; Whitham, S.A.; Carrington, J.C. Loss-of-susceptibility mutants of arabidopsis thaliana reveal an essential role for eif(iso)4e during potyvirus infection. Curr. Biol. 2002, 12, 1046–1051. [Google Scholar] [CrossRef] [Green Version]
- Hyodo, K.; Okuno, T. Pathogenesis mediated by proviral host factors involved in translation and replication of plant positive-strand rna viruses. Curr. Opin. Virol. 2016, 17, 11–18. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H. Host factors against plant viruses. Mol. Plant. Pathol. 2019, 20, 1588–1601. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Yeakley, J.M.; Garcia, E.W.; Holdridge, J.D.; Fan, J.B.; Whitham, S.A. Salicylic acid-dependent expression of host genes in compatible arabidopsis-virus interactions. Plant Physiol. 2005, 137, 1147–1159. [Google Scholar] [CrossRef] [Green Version]
- Ascencio-Ibanez, J.T.; Sozzani, R.; Lee, T.J.; Chu, T.M.; Wolfinger, R.D.; Cella, R.; Hanley-Bowdoin, L. Global analysis of arabidopsis gene expression uncovers a complex array of changes impacting pathogen response and cell cycle during geminivirus infection. Plant Physiol. 2008, 148, 436–454. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Jiang, G.; Wu, J.; Liu, Y.; Qian, Y.; Zhou, X. Characterization of a novel polerovirus infecting maize in china. Viruses 2016, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Fan, B.; Chen, C.; Chen, Z. An important role of an inducible rna-dependent rna polymerase in plant antiviral defense. Proc. Natl. Acad. Sci. USA 2001, 98, 6516–6521. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.; Edelbaum, O.; Sela, I. Tobacco mosaic virus regulates the expression of its own resistance gene n. Plant Physiol. 2004, 135, 2392–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgyan, J.; Havelda, Z. Viral suppressors of rna silencing. Trends Plant. Sci. 2011, 16, 265–272. [Google Scholar] [CrossRef] [PubMed]
- De Ronde, D.; Butterbach, P.; Kormelink, R. Dominant resistance against plant viruses. Front. Plant Sci. 2014, 5, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Wang, A. Rna decay is an antiviral defense in plants that is counteracted by viral rna silencing suppressors. PLoS Pathog. 2018, 14, e1007228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Wang, A. The potyvirus silencing suppressor protein vpg mediates degradation of sgs3 via ubiquitination and autophagy pathways. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Pollari, M.; De, S.; Wang, A.; Mäkinen, K. The potyviral silencing suppressor hcpro recruits and employs host argonaute1 in pro-viral functions. PLoS Pathog. 2020, 16, e1008965. [Google Scholar] [CrossRef]
- Kovalev, N.; de Castro, I.F.; Pogany, J.; Barajas, D.; Pathak, K.; Risco, C.; Nagy, P.D. Role of viral rna and co-opted cellular escrt-i and escrt-iii factors in formation of tombusvirus spherules harboring the tombusvirus replicase. J. Virol. 2016, 90, 3611–3626. [Google Scholar] [CrossRef] [Green Version]
- Hwang, Y.T.; McCartney, A.W.; Gidda, S.K.; Mullen, R.T. Localization of the carnation italian ringspot virus replication protein p36 to the mitochondrial outer membrane is mediated by an internal targeting signal and the tom complex. BMC Cell Biol. 2008, 9, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufresne, P.J.; Thivierge, K.; Cotton, S.; Beauchemin, C.; Ide, C.; Ubalijoro, E.; Laliberte, J.F.; Fortin, M.G. Heat shock 70 protein interaction with turnip mosaic virus rna-dependent rna polymerase within virus-induced membrane vesicles. Virology 2008, 374, 217–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mine, A.; Hyodo, K.; Tajima, Y.; Kusumanegara, K.; Taniguchi, T.; Kaido, M.; Mise, K.; Taniguchi, H.; Okuno, T. Differential roles of hsp70 and hsp90 in the assembly of the replicase complex of a positive-strand rna plant virus. J. Virol. 2012, 86, 12091–12104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Rojas, M.R.; Park, M.R.; Seo, Y.S.; Lucas, W.J.; Gilbertson, R.L. Histone h3 interacts and colocalizes with the nuclear shuttle protein and the movement protein of a geminivirus. J. Virol. 2011, 85, 11821–11832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, Y.-L.; Hung, Y.-J.; Tseng, Y.-H.; Hsu, H.-T.; Yang, J.-Y.; Wung, C.-H.; Lin, N.-S.; Meng, M.; Hsu, Y.-H.; Chang, B.-Y. The stable association of virion with the triple-gene-block protein 3-based complex of bamboo mosaic virus. PLoS Pathog. 2013, 9, e1003405. [Google Scholar] [CrossRef] [Green Version]
- Okinaka, Y.; Mise, K.; Suzuki, E.; Okuno, T.; Furusawa, I. The c terminus of brome mosaic virus coat protein controls viral cell-to-cell and long-distance movement. J. Virol. 2001, 75, 5385–5390. [Google Scholar] [CrossRef] [Green Version]
- Dalmay, T.; Rubino, L.; Burgyan, J.; Kollar, A.; Russo, M. Functional analysis of cymbidium ringspot virus genome. Virology 1993, 194, 697–704. [Google Scholar] [CrossRef]
- Xiang, Y.; Kakani, K.; Reade, R.; Hui, E.; Rochon, D. A 38-amino-acid sequence encompassing the arm domain of the cucumber necrosis virus coat protein functions as a chloroplast transit peptide in infected plants. J. Virol. 2006, 80, 7952–7964. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Macfarlane, S.; Kalinina, N.O.; Rakitina, D.V.; Ryabov, E.V.; Gillespie, T.; Haupt, S.; Brown, J.W.; Taliansky, M. Interaction of a plant virus-encoded protein with the major nucleolar protein fibrillarin is required for systemic virus infection. Proc. Natl. Acad. Sci. USA 2007, 104, 11115–11120. [Google Scholar] [CrossRef] [Green Version]
- Movahed, N.; Sun, J.; Vali, H.; Laliberte, J.F.; Zheng, H. A host er fusogen is recruited by turnip mosaic virus for maturation of viral replication vesicles. Plant Physiol. 2019, 179, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Mathioudakis, M.M.; Khechmar, S.; Owen, C.A.; Medina, V.; Ben Mansour, K.; Tomaszewska, W.; Spanos, T.; Sarris, P.F.; Livieratos, I.C. A thioredoxin domain-containing protein interacts with pepino mosaic virus triple gene block protein 1. Int. J. Mol. Sci. 2018, 19, 3747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verchot-Lubicz, J.; Torrance, L.; Solovyev, A.G.; Morozov, S.Y.; Jackson, A.O.; Gilmer, D. Varied movement strategies employed by triple gene block-encoding viruses. Mol. Plant Microbe Interact. 2010, 23, 1231–1247. [Google Scholar] [CrossRef] [Green Version]
- Turner, K.A.; Sit, T.L.; Callaway, A.S.; Allen, N.S.; Lommel, S.A. Red clover necrotic mosaic virus replication proteins accumulate at the endoplasmic reticulum. Virology 2004, 320, 276–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusumanegara, K.; Mine, A.; Hyodo, K.; Kaido, M.; Mise, K.; Okuno, T. Identification of domains in p27 auxiliary replicase protein essential for its association with the endoplasmic reticulum membranes in red clover necrotic mosaic virus. Virology 2012, 433, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Kaido, M.; Tsuno, Y.; Mise, K.; Okuno, T. Endoplasmic reticulum targeting of the red clover necrotic mosaic virus movement protein is associated with the replication of viral rna1 but not that of rna2. Virology 2009, 395, 232–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, A.; Zheng, J.Y.; Lazarowitz, S.G. Synaptotagmin syta forms er-plasma membrane junctions that are recruited to plasmodesmata for plant virus movement. Curr. Biol. 2015, 25, 2018–2025. [Google Scholar] [CrossRef] [Green Version]
- McCartney, A.W.; Greenwood, J.S.; Fabian, M.R.; White, K.A.; Mullen, R.T. Localization of the tomato bushy stunt virus replication protein p33 reveals a peroxisome-to-endoplasmic reticulum sorting pathway. Plant Cell 2005, 17, 3513–3531. [Google Scholar] [CrossRef] [Green Version]
- Scholthof, H.B.; Scholthof, K.B.; Kikkert, M.; Jackson, A.O. Tomato bushy stunt virus spread is regulated by two nested genes that function in cell-to-cell movement and host-dependent systemic invasion. Virology 1995, 213, 425–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, S.; Watanabe, Y.; Beachy, R.N. Tobacco mosaic virus infection spreads cell to cell as intact replication complexes. Proc. Natl. Acad. Sci. USA 2004, 101, 6291–6296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Liu, Y.; Sun, X.; Qian, W.; Zhang, D.; Qiu, B. Characterization of a specific interaction between ip-l, a tobacco protein localized in the thylakoid membranes, and tomato mosaic virus coat protein. Biochem. Biophys. Res. Commun. 2008, 374, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.; Yan, Z.Y.; Cheng, D.J.; Liu, J.; Tian, Y.P.; Zhu, C.X.; Wang, H.Y.; Li, X.D. Tobacco vein banding mosaic virus 6k2 protein hijacks nbpsbo1 for virus replication. Sci. Rep. 2017, 7, 43455. [Google Scholar] [CrossRef]
- Geng, C.; Cong, Q.Q.; Li, X.D.; Mou, A.L.; Gao, R.; Liu, J.L.; Tian, Y.P. Developmentally regulated plasma membrane protein of nicotiana benthamiana contributes to potyvirus movement and transports to plasmodesmata via the early secretory pathway and the actomyosin system. Plant Physiol. 2015, 167, 394–410. [Google Scholar] [CrossRef] [Green Version]
- Hiraguri, A.; Netsu, O.; Sasaki, N.; Nyunoya, H.; Sasaya, T. Recent progress in research on cell-to-cell movement of rice viruses. Front. Microbiol. 2014, 5, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Chen, H.; Chen, Q.; Omura, T.; Xie, L.; Wu, Z.; Wei, T. The early secretory pathway and an actin-myosin viii motility system are required for plasmodesmatal localization of the nsvc4 protein of rice stripe virus. Virus Res. 2011, 159, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.K.; Lian, S.; Kim, S.M.; Park, S.H.; Kim, K.H. Current insights into research on rice stripe virus. Plant Pathol. J. 2013, 29, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harries, P.A.; Palanichelvam, K.; Yu, W.C.; Schoelz, J.E.; Nelson, R.S. The cauliflower mosaic virus protein p6 forms motile inclusions that traffic along actin microfilaments and stabilize microtubules. Plant Physiol. 2009, 149, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Thiébeauld, O.; Schepetilnikov, M.; Park, H.-S.; Geldreich, A.; Kobayashi, K.; Keller, M.; Hohn, T.; Ryabova, L.A. A new plant protein interacts with eif3 and 60s to enhance virus-activated translation re-initiation. EMBO J. 2009, 28, 3171–3184. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.W.; Hu, C.C.; Liou, M.R.; Chang, B.Y.; Tsai, C.H.; Meng, M.; Lin, N.S.; Hsu, Y.H. Hsp90 interacts specifically with viral rna and differentially regulates replication initiation of bamboo mosaic virus and associated satellite rna. PLoS Pathog. 2012, 8, e1002726. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.H.; Chiu, M.H.; Cheng, S.F.; Hsu, Y.H.; Tsai, C.H. The glutathione transferase of nicotiana benthamiana nbgstu4 plays a role in regulating the early replication of bamboo mosaic virus. New Phytol. 2013, 199, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Diaz, A.; Zhang, J.; Ollwerther, A.; Wang, X.; Ahlquist, P. Host escrt proteins are required for bromovirus rna replication compartment assembly and function. PLoS Pathog. 2015, 11, e1004742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungfleisch, J.; Chowdhury, A.; Alves-Rodrigues, I.; Tharun, S.; Diez, J. The lsm1-7-pat1 complex promotes viral rna translation and replication by differential mechanisms. RNA 2015, 21, 1469–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diez, J.; Ishikawa, M.; Kaido, M.; Ahlquist, P. Identification and characterization of a host protein required for efficient template selection in viral rna replication. Proc. Natl. Acad. Sci. USA 2000, 97, 3913–3918. [Google Scholar] [CrossRef] [Green Version]
- Richardson, L.G.; Clendening, E.A.; Sheen, H.; Gidda, S.K.; White, K.A.; Mullen, R.T. A unique n-terminal sequence in the carnation italian ringspot virus p36 replicase-associated protein interacts with the host cell escrt-i component vps23. J. Virol. 2014, 88, 6329–6344. [Google Scholar] [CrossRef] [Green Version]
- Barajas, D.; Xu, K.; de Castro Martin, I.F.; Sasvari, Z.; Brandizzi, F.; Risco, C.; Nagy, P.D. Co-opted oxysterol-binding orp and vap proteins channel sterols to rna virus replication sites via membrane contact sites. PLoS Pathog. 2014, 10, e1004388. [Google Scholar] [CrossRef] [Green Version]
- Mathioudakis, M.; Rita, S.L.V.; Canto, T.; Medina, V.; Mossialos, D.; Makris Antonios, M.; Livieratos, I. Pepino mosaic virus triple gene block protein 1 (tgbp1) interacts with and increases tomato catalase 1 activity to enhance virus accumulation. Mol. Plant Pathol. 2013, 14, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Fridborg, I.; Grainger, J.; Page, A.; Coleman, M.; Findlay, K.; Angell, S. Tip, a novel host factor linking callose degradation with the cell-to-cell movement of potato virus x. Mol. Plant Microbe Interact. 2003, 16, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Hyodo, K.; Suzuki, N.; Okuno, T. Hijacking a host scaffold protein, rack1, for replication of a plant rna virus. New Phytol. 2019, 221, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xiang, C.Y.; Yang, J.; Chen, J.P.; Zhang, H.M. Interaction of hsp20 with a viral rdrp changes its sub-cellular localization and distribution pattern in plants. Sci. Rep. 2015, 5, 14016. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Pogany, J.; Tupman, S.; Esposito, A.M.; Kinzy, T.G.; Nagy, P.D. Translation elongation factor 1a facilitates the assembly of the tombusvirus replicase and stimulates minus-strand synthesis. PLoS Pathog. 2010, 6, e1001175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barajas, D.; Jiang, Y.; Nagy, P.D. A unique role for the host escrt proteins in replication of tomato bushy stunt virus. PLoS Pathog. 2009, 5, e1000705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.Y.; Nagy, P.D. Tomato bushy stunt virus co-opts the rna-binding function of a host metabolic enzyme for viral genomic rna synthesis. Cell Host Microbe 2008, 3, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Pogany, J.; Stork, J.; Li, Z.; Nagy, P.D. In vitro assembly of the tomato bushy stunt virus replicase requires the host heat shock protein 70. Proc. Natl. Acad. Sci. USA 2008, 105, 19956–19961. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.Y.-L.; Stork, J.; Nagy, P.D. A key role for heat shock protein 70 in the localization and insertion of tombusvirus replication proteins to intracellular membranes. J. Virol. 2009, 83, 3276–3287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, Y.; Kobayashi, T.; Hamada, K.; Sakurai, K.; Yoshii, A.; Suzuki, M.; Namba, S.; Hibi, T. In vivo interaction between tobacco mosaic virus rna-dependent rna polymerase and host translation elongation factor 1a. Virology 2006, 347, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.S.; Wei, T.; Laliberte, J.; Wang, A. A host rna helicase-like protein, atrh8, interacts with the potyviral genome-linked protein, vpg, associates with the virus accumulation complex, and is essential for infection. Plant Physiol. 2010, 152, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xiong, R.; Bernards, M.; Wang, A. Recruitment of arabidopsis rna helicase atrh9 to the viral replication complex by viral replicase to promote turnip mosaic virus replication. Sci. Rep. 2016, 6, 30297. [Google Scholar] [CrossRef] [Green Version]
- Thivierge, K.; Cotton, S.; Dufresne, P.J.; Mathieu, I.; Beauchemin, C.; Ide, C.; Fortin, M.G.; Laliberte, J.F. Eukaryotic elongation factor 1a interacts with turnip mosaic virus rna-dependent rna polymerase and vpg-pro in virus-induced vesicles. Virology 2008, 377, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, C.; Laliberte, J.F. The poly(a) binding protein is internalized in virus-induced vesicles or redistributed to the nucleolus during turnip mosaic virus infection. J. Virol. 2007, 81, 10905–10913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikiori, M.; Dohi, K.; Mori, M.; Meshi, T.; Naito, S.; Ishikawa, M. Membrane-bound tomato mosaic virus replication proteins participate in rna synthesis and are associated with host proteins in a pattern distinct from those that are not membrane bound. J. Virol. 2006, 80, 8459–8468. [Google Scholar] [CrossRef] [Green Version]
- Gorovits, R.; Moshe, A.; Ghanim, M.; Czosnek, H. Recruitment of the host plant heat shock protein 70 by tomato yellow leaf curl virus coat protein is required for virus infection. PLoS ONE 2013, 8, e70280. [Google Scholar] [CrossRef]
- Xu, K.; Nagy, P.D. Enrichment of phosphatidylethanolamine in viral replication compartments via co-opting the endosomal rab5 small gtpase by a positive-strand rna virus. PLoS Biol. 2016, 14, e2000128. [Google Scholar] [CrossRef] [Green Version]
- Diaz, A.; Wang, X.; Ahlquist, P. Membrane-shaping host reticulon proteins play crucial roles in viral rna replication compartment formation and function. Proc. Natl. Acad. Sci. USA 2010, 107, 16291–16296. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Zhang, C.; Hou, X.; Sanfaçon, H.; Wang, A. The snare protein syp71 is essential for turnip mosaic virus infection by mediating fusion of virus-induced vesicles with chloroplasts. PLoS Pathog. 2013, 9, e1003378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.P.; Jhuo, J.H.; Tsai, M.S.; Tsai, C.H.; Chen, H.C.; Lin, N.S.; Hsu, Y.H.; Cheng, C.P. Nbrabg3f, a member of rab gtpase, is involved in bamboo mosaic virus infection in nicotiana benthamiana. Mol. Plant Pathol. 2016, 17, 714–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carluccio, A.V.; Zicca, S.; Stavolone, L. Hitching a ride on vesicles: Cauliflower mosaic virus movement protein trafficking in the endomembrane system. Plant Physiol. 2014, 164, 1261. [Google Scholar] [CrossRef] [Green Version]
- Hyodo, K.; Mine, A.; Taniguchi, T.; Kaido, M.; Mise, K.; Taniguchi, H.; Okuno, T. Adp ribosylation factor 1 plays an essential role in the replication of a plant rna virus. J. Virol. 2013, 87, 163–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.; Angel, C.A.; Lutz, L.; Leisner, S.M.; Nelson, R.S.; Schoelz, J.E. Association of the p6 protein of cauliflower mosaic virus with plasmodesmata and plasmodesmal proteins. Plant Physiol. 2014, 166, 1345–1358. [Google Scholar] [CrossRef] [Green Version]
- Hyodo, K.; Hashimoto, K.; Kuchitsu, K.; Suzuki, N.; Okuno, T. Harnessing host ros-generating machinery for the robust genome replication of a plant rna virus. Proc. Natl. Acad. Sci. USA 2017, 114, E1282–E1290. [Google Scholar] [CrossRef] [Green Version]
- Vijayapalani, P.; Maeshima, M.; Nagasaki-Takekuchi, N.; Miller, W.A. Interaction of the trans-frame potyvirus protein p3n-pipo with host protein pcap1 facilitates potyvirus movement. PLoS Pathog. 2012, 8, e1002639. [Google Scholar] [CrossRef]
- Yoshii, A.; Shimizu, T.; Yoshida, A.; Hamada, K.; Sakurai, K.; Yamaji, Y.; Suzuki, M.; Namba, S.; Hibi, T. Nth201, a novel class ii knotted1-like protein, facilitates the cell-to-cell movement of tobacco mosaic virus in tobacco. Mol. Plant Microbe Interact. 2008, 21, 586–596. [Google Scholar] [CrossRef] [Green Version]
- Uhrig, J.F.; Canto, T.; Marshall, D.; MacFarlane, S.A. Relocalization of nuclear aly proteins to the cytoplasm by the tomato bushy stunt virus p19 pathogenicity protein. Plant Physiol. 2004, 135, 2411–2423. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara-Komoda, Y.; Hirai, K.; Mochizuki, A.; Nishiguchi, M.; Meshi, T.; Ishikawa, M. Overexpression of a host factor tom1 inhibits tomato mosaic virus propagation and suppression of rna silencing. Virology 2008, 376, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, T.; Imai, T.; Satoh, R.; Kawashima, A.; Takahashi, M.; Tomita, K.; Kubota, K.; Meshi, T.; Naito, S.; Ishikawa, M. Complete inhibition of tobamovirus multiplication by simultaneous mutations in two homologous host genes. J. Virol. 2002, 76, 2491–2497. [Google Scholar] [CrossRef] [Green Version]
- Hafren, A.; Macia, J.L.; Love, A.J.; Milner, J.J.; Drucker, M.; Hofius, D. Selective autophagy limits cauliflower mosaic virus infection by nbr1-mediated targeting of viral capsid protein and particles. Proc. Natl. Acad. Sci. USA 2017, 114, E2026–E2035. [Google Scholar] [CrossRef] [Green Version]
- Ballut, L.; Drucker, M.; Pugniere, M.; Cambon, F.; Blanc, S.; Roquet, F.; Candresse, T.; Schmid, H.P.; Nicolas, P.; Gall, O.L.; et al. Hcpro, a multifunctional protein encoded by a plant rna virus, targets the 20s proteasome and affects its enzymic activities. J. Gen. Virol. 2005, 86, 2595–2603. [Google Scholar] [CrossRef]
- Dielen, A.S.; Sassaki, F.T.; Walter, J.; Michon, T.; Menard, G.; Pagny, G.; Krause-Sakate, R.; Ide, G.M.; Badaoui, S.; Le Gall, O.; et al. The 20s proteasome alpha5 subunit of arabidopsis thaliana carries an rnase activity and interacts in planta with the lettuce mosaic potyvirus hcpro protein. Mol. Plant Pathol. 2011, 12, 137–150. [Google Scholar] [CrossRef]
- Cho, S.Y.; Cho, W.K.; Choi, H.S.; Kim, K.H. Cis-acting element (sl1) of potato virus x controls viral movement by interacting with the nbmpb2cb and viral proteins. Virology 2012, 427, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.Y.; Nagy, P.D. Blocking tombusvirus replication through the antiviral functions of ddx17-like rh30 dead-box helicase. PLoS Pathog. 2019, 15, e1007771. [Google Scholar] [CrossRef] [PubMed]
- Hafren, A.; Ustun, S.; Hochmuth, A.; Svenning, S.; Johansen, T.; Hofius, D. Turnip mosaic virus counteracts selective autophagy of the viral silencing suppressor hcpro. Plant Physiol. 2018, 176, 649–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltazar, B.M.; Espinoza, L.C.; Banda, A.E.; de la Fuente Martinez, J.M.; Tiznado, J.A.G.; Garcia, J.G.; Gutierrez, M.A.; Rodriguez, J.L.G.; Diaz, O.H.; Horak, M.J.; et al. Pollen-mediated gene flow in maize: Implications for isolation requirements and coexistence in mexico, the center of origin of maize. PLoS ONE 2015, 10, e0131549. [Google Scholar] [CrossRef]
- Timperio, A.; Egidi, M.; Zolla, L. Proteomics applied on plant abiotic stresses: Role of heat shock proteins (hsp). Elsevier 2008, 71, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.P.; Chen, I.H.; Tsai, C.H. Host factors in the infection cycle of bamboo mosaic virus. Front. Microbiol. 2017, 8, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wollert, T.; Hurley, J. Molecular mechanism of multivesicular body biogenesis by escrt complexes. Nature 2010, 464, 864–869. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Paredes, C.A.; Silva-Rosales, L.; Daros, J.A.; Alejandri-Ramirez, N.D.; Dinkova, T.D. The absence of eukaryotic initiation factor eif(iso)4e affects the systemic spread of a tobacco etch virus isolate in arabidopsis thaliana. Mol. Plant Microbe Interact. 2013, 26, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Morris, D.R.; Geballe, A.P. Upstream open reading frames as regulators of mrna translation. Mol. Cell. Biol. 2000, 20, 8635–8642. [Google Scholar] [CrossRef] [Green Version]
- Cordin, O.; Banroques, J.; Tanner, N.K.; Linder, P. The dead-box protein family of rna helicases. Gene 2006, 367, 17–37. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, K.A.; Oh, R.T.; Park, C.M.; Kang, H. Functional characterization of dead-box rna helicases in arabidopsis thaliana under abiotic stress conditions. Plant. Cell Physiol. 2008, 49, 1563–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noueiry, A.O.; Chen, J.; Ahlquist, P. A mutant allele of essential, general translation initiation factor ded1 selectively inhibits translation of a viral mrna. Proc. Natl. Acad. Sci. USA 2000, 97, 12985–12990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Serviene, E.; Gal, J.; Panavas, T.; Nagy, P.D. Identification of essential host factors affecting tombusvirus rna replication based on the yeast tet promoters hughes collection. J. Virol. 2006, 80, 7394–7404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasanth, K.R.; Huang, Y.W.; Liou, M.R.; Wang, R.Y.; Hu, C.C.; Tsai, C.H.; Meng, M.; Lin, N.S.; Hsu, Y.H. Glyceraldehyde 3-phosphate dehydrogenase negatively regulates the replication of bamboo mosaic virus and its associated satellite rna. J. Virol. 2011, 85, 8829–8840. [Google Scholar] [CrossRef] [Green Version]
- Dixon, D.; Lapthorn, A.; Edwards, R. Protein family review: Plant glutathione transferases. Genome Biol. 2002, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skopelitou, K.; Muleta, A.W.; Papageorgiou, A.C.; Chronopoulou, E.; Labrou, N.E. Catalytic features and crystal structure of a tau class glutathione transferase from glycine max specifically upregulated in response to soybean mosaic virus infections. Biochim. Biophys. Acta 2015, 1854, 166–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber-Boyvat, M.; Zhong, W.; Yan, D.; Olkkonen, V.M. Oxysterol-binding proteins: Functions in cell regulation beyond lipid metabolism. Biochem. Pharmacol. 2013, 86, 89–95. [Google Scholar] [CrossRef]
- Islas-Flores, T.; Rahman, A.; Ullah, H.; Villanueva, M.A. The receptor for activated c kinase in plant signaling: Tale of a promiscuous little molecule. Front. Plant Sci. 2015, 6, 1090. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, A.; Chen, L.; Thao, N.P.; Fujiwara, M.; Wong, H.L.; Kuwano, M.; Umemura, K.; Shirasu, K.; Kawasaki, T.; Shimamoto, K. Rack1 functions in rice innate immunity by interacting with the rac1 immune complex. Plant Cell 2008, 20, 2265–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Li, J.F.; Niu, Y.; Zhang, X.C.; Woody, O.Z.; Xiong, Y.; Djonovic, S.; Millet, Y.; Bush, J.; McConkey, B.J.; et al. Pathogen-secreted proteases activate a novel plant immune pathway. Nature 2015, 521, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.R.; Ron, D.; Kiely, P.A. Rack1, a multifaceted scaffolding protein: Structure and function. Cell Commun. Signal. 2011, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Otegui, M.S.; Spitzer, C. Endosomal functions in plants. Traffic 2008, 9, 1589–1598. [Google Scholar] [CrossRef]
- Lundquist, E. Small GTPases. WormBook 2006, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Brey, I.; Bartenschlager, R. Endoplasmic reticulum: The favorite intracellular niche for viral replication and assembly. Viruses 2016, 8, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanderfoot, A.; Kovaleva, V.; Bassham, D.; Raikhel, N. Interactions between syntaxins identify at least five snare complexes within the golgi/prevacuolar system of the arabidopsis cell. Mol. Biol. Cell 2001, 12, 3733–3743. [Google Scholar] [CrossRef]
- Suwastika, N.; Uemura, T.; Shiina, T.; Sato, M.; Takeyasu, K. Suwastika in, uemura t, shiina t, sato mh, takeyasu. Cell Struct. Funct. 2008, 33, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Nziengui, H.; Schoefs, B. Functions of reticulons in plants: What we can learn from animals and yeasts. Cell Mol. Life Sci. 2009, 66, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Stefano, G.; Brandizzi, F.; Zheng, H. Arabidopsis rhd3 mediates the generation of the tubular er network and is required for golgi distribution and motility in plant cells. J. Cell Sci. 2011, 124, 2241–2252. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.-F.; Huang, Y.-P.; Wu, Z.-R.; Hu, C.-C.; Hsu, Y.-H.; Tsai, C.-H. Identification of differentially expressed genes induced by bamboo mosaic virus infection in nicotiana benthamianaby cdna-amplified fragment length polymorphism. BMC Plant Biol. 2010, 10, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memon, A.R. The role of adp-ribosylation factor and sar1 in vesicular trafficking in plants. Biochim. Biophys. Acta 2004, 1664, 9–30. [Google Scholar] [CrossRef] [Green Version]
- Harries, P.; Ding, B. Cellular factors in plant virus movement: At the leading edge of macromolecular trafficking in plants. Virology 2011, 411, 237–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory burst oxidases: The engines of ros signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [Google Scholar] [CrossRef]
- Deng, X.G.; Zhu, T.; Zou, L.J.; Han, X.Y.; Zhou, X.; Xi, D.H.; Zhang, D.W.; Lin, H.H. Orchestration of hydrogen peroxide and nitric oxide in brassinosteroid-mediated systemic virus resistance in nicotiana benthamiana. Plant J. 2016, 85, 478–493. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.-G.; Zhu, T.; Peng, X.-J.; Xi, D.-H.; Guo, H.; Yin, Y.; Zhang, D.-W.; Lin, H.-H. Role of brassinosteroid signaling in modulating tobacco mosaic virus resistance in nicotiana benthamiana. Sci. Rep. 2016, 6, 20579. [Google Scholar] [CrossRef] [PubMed]
- Pfaff, C.; Ehrnsberger, H.; Flores-Tornero, M.; Sorensen, B.; Schubert, T.; Längst, G.; Griesenbeck, L.; Sprunck, S.; Grasser, M.; Grasser, K.D. Aly rna-binding proteins are required for nucleocytosolic mrna transport and modulate plant growth and development. Plant Physiol. 2018, 177, 2226–2240. [Google Scholar] [CrossRef]
- Canto, T.; Uhrig, J.F.; Swanson, M.; Wright, K.M.; MacFarlane, S.A. Translocation of tomato bushy stunt virus p19 protein into the nucleus by aly proteins compromises its silencing suppressor activity. J. Virol. 2006, 80, 9064–9072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavel, M.; Michaeli, S.; Genschik, P. Autophagy: A double-edged sword to fight plant viruses. Trends Plant Sci. 2017, 22, 646–648. [Google Scholar] [CrossRef] [PubMed]
- Haxim, Y.; Ismayil, A.; Jia, Q.; Wang, Y.; Zheng, X.; Chen, T.; Qian, L.; Liu, N.; Wang, Y.; Han, S.; et al. Autophagy functions as an antiviral mechanism against geminiviruses in plants. eLife 2017, 6, e23897. [Google Scholar] [CrossRef] [PubMed]
- Hafren, A.; Lohmus, A.; Makinen, K. Formation of potato virus a-induced rna granules and viral translation are interrelated processes required for optimal virus accumulation. PLoS Pathog. 2015, 11, e1005314. [Google Scholar] [CrossRef] [Green Version]
- Pitzalis, N.; Amari, K.; Graindorge, S.; Pflieger, D.; Donaire, L.; Wassenegger, M.; Llave, C.; Heinlein, M. Turnip mosaic virus in oilseed rape activates networks of srna-mediated interactions between viral and host genomes. Commun. Biol. 2020, 3, 702. [Google Scholar] [CrossRef]
- Beauchemin, C.; Boutet, N.; Laliberte, J.F. Visualization of the interaction between the precursors of vpg, the viral protein linked to the genome of turnip mosaic virus, and the translation eukaryotic initiation factor iso 4e in planta. J. Virol. 2007, 81, 775–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigam, D.; LaTourrette, K.; Souza, P.F.N.; Garcia-Ruiz, H. Genome-wide variation in potyviruses. Front. Plant Sci. 2019, 10, 1439. [Google Scholar] [CrossRef] [Green Version]
- Charron, C.; Nicolaï, M.; Gallois, J.L.; Robaglia, C.; Moury, B.; Palloix, A.; Caranta, C. Natural variation and functional analyses provide evidence for co-evolution between plant eif4e and potyviral vpg. Plant J. 2008, 54, 56–68. [Google Scholar] [CrossRef]
- Ahlquist, P.; Noueiry, A.O.; Lee, W.M.; Kushner, D.B.; Dye, B.T. Host factors in positive-strand rna virus genome replication. J. Virol. 2003, 77, 8181–8186. [Google Scholar] [CrossRef] [Green Version]
- Panavas, T.; Serviene, E.; Brasher, J.; Nagy, P.D. Yeast genome-wide screen reveals dissimilar sets of host genes affecting replication of rna viruses. Proc. Natl. Acad. Sci. USA 2005, 102, 7326–7331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, Y.J.; Venkatesh, J.; Lee, J.H.; Kim, J.; Lee, H.E.; Kim, D.S.; Kang, B.C. Genome editing of eif4e1 in tomato confers resistance to pepper mottle virus. Front. Plant Sci. 2020, 11, 1098. [Google Scholar] [CrossRef] [PubMed]
- Mahuku, G.; Lockhart, B.E.; Wanjala, B.; Jones, M.W.; Kimunye, J.N.; Stewart, L.R.; Cassone, B.J.; Sevgan, S.; Nyasani, J.O.; Kusia, E.; et al. Maize lethal necrosis (mln), an emerging threat to maize-based food security in sub-saharan africa. Phytopathology 2015, 105, 956–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Virus | RdRp | Site of Replication | Intracellular Movement of the Replication Compartments | Cell-To-Cell Movement Form | Reference |
|---|---|---|---|---|---|
| Group II: Single-strand DNA | |||||
| TYLCV | Rep protein | Nucleus | From nucleus to plasmodesmata | Minichromosome | [22] |
| Group IV: Single positive-strand RNA | |||||
| BaMV | 155 kDa | Chloroplast | From chloroplast to plasmodesmata | Virions or ribonucleoprotein particles | [23] |
| BMV | 2a | Endoplasmic reticulum | Non-motile | Virions or ribonucleoprotein particles | [24] |
| CIRV | p36 | Mitochondria | Non-motile | Ribonucleoprotein particles | [25] |
| CNV | p33 | Peroxisome | Non-motile | Ribonucleoprotein particles | [26] |
| GRV | RdRp | Cytoplasm | NA | Ribonucleoprotein particles | [27] |
| LMV | NIb | Endoplasmic reticulum | From ER to plasmodesmata | Replication vesicles | [28] |
| PepMV | 164 kDa | Cytoplasm (membrane association with ER is unclear) | From cytoplasm to plasmodesmata | Ribonucleoprotein particles | [29] |
| PVX | RdRp | Endoplasmic reticulum | NA | Virions or ribonucleoprotein particles | [30] |
| RCNMV | p27 and p88 | Endoplasmic reticulum | From ER to plasmodesmata | Virions | [31,32,33] |
| TVCV | RdRp | Endoplasmic reticulum | From ER to plasmodesmata | Virions or ribonucleoprotein particles | [34] |
| TBSV | p92pol | Peroxisomes | Non-motile | Ribonucleoprotein particles | [35,36] |
| TMV | RdRp | Endoplasmic reticulum | From ER to plasmodesmata | Replication complexes or ribonucleoprotein particles | [37] |
| ToMV | 130K and 180K | Endoplasmic reticulum | From ER to plasmodesmata | Virions or ribonucleoprotein particles | [38] |
| TuMV | NIb | ER and chloroplasts | From ER to chloroplast and/or to Golgi apparatus and to plasmodesmata | Replication vesicles | [28] |
| TVBMV | NIb | Chloroplasts | From ER to chloroplast and/or to Golgi apparatus and to plasmodesmata | Replication vesicles | [39,40] |
| Group V: Single negative-strand RNA | |||||
| RSV | 337 kDa | Cytoplasm (membrane association is unknown) | From ER to Golgi to plasmodesmata | Virion–protein complexes | [41,42,43] |
| Group VII: Double-strand DNA-RT | |||||
| CaMV | Rep protein | Nucleus | From nucleus to ER and/or directly to plasmodesmata | Virions | [44] |
| Virus | Viral Protein or RNA | Host Protein | Host | Movement of Host Protein into | Role | Initial Detection | Mechanism of Interaction | Experimental System for Detecting of New Localization Sites * | Method of Observation: Time | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Cytoplasmic proteins | ||||||||||
| CaMV | TAV | RISP | Arabidopsis thaliana | Inclusion bodies (cytoplasmic and nuclear) | Stimulates translation re-initiation | Yeast two hybrid | Protein–protein | Brassica rapa leaves | Immunofluorescence and confocal microscopy: 15 dpi | [45] |
| BaMV | 155 kDa and 3′ UTR | HSP90 | Nicotiana benthamiana | Chloroplast | Formation of replication compartments | Partially purified replicase | Protein–protein and RNA–protein | Saccharomyces cerevisiae and Escherichia coli | Yeast two hybrid, GST-pull down | [46] |
| 3′ UTR | NbGSTU4 | N. benthamiana | Chloroplast | Binds to the 3′ UTR and stimulates negative-strand RNA synthesis | Partially purified replicase | RNA–protein | E. coli | UV crosslink | [47] | |
| BMV | 1a | ESCRT- III | S. cerevisiae | Perinuclear ER | Formation of replication compartments | Yeast genetic analysis | Protein–protein | S. cerevisiae | Immunofluorescence and confocal microscopy: 48 h | [48] |
| 1a and 2b | LSM1 | S. cerevisiae | ER | Promotes viral RNA translation | Yeast mutagenesis | Protein–protein | S. cerevisiae | Immunofluorescence and confocal microscopy: 48 h | [49,50] | |
| CIRV | p36 | ESCRT-I | N. benthamiana | Mitochondria | Formation of replication compartments | Split ubiquitin assay | Protein–protein | S. cerevisiae | Immunofluorescence and confocal microscopy: 15-45 min | [51] |
| p36 | ORP | N. benthamiana and S. cerevisiae | Mitochondria and ER | Formation of replication compartments | Yeast two hybrid | Protein–protein | N. benthamiana leaves | BiFC: 48 h | [52] | |
| PepMV | p26 | Catalase 1 | Solanum lycopersicum | Cytoplasm and nucleus | Antagonist to antiviral response | Yeast two hybrid | Protein–protein | N. benthamiana leaves | BiFC, immunolabeling, and electron microscopy: 3–4 dpi | [53] |
| PVX | TGB12K | TIP | Nicotiana tabacum | Peripheral bodies | Regulates plasmodesmata opening | Yeast two hybrid | Protein–protein | N. benthamiana leaves | Confocal microscopy: 3 dpi | [54] |
| RCNMV | p27 | HSP70 | N. benthamiana | ER | Formation of replication compartments | Affinity purification | Protein–protein | N. benthamiana leaves | Confocal microscopy: 3 dpi | [21] |
| p27 | NbRACK1 | N. benthamiana | ER-derived aggregates | Increases ROS to benefit the virus | Co-immunoprecipitation | Protein–protein | N. benthamiana leaves | BiFC: 4 dpi | [55] | |
| RSV | 337 kDa | HSP20 | N. benthamiana and Oryza sativa | Nucleus | Antagonist to antiviral response | Yeast two hybrid | Protein–protein | N. benthamiana leaves | BiFC: 48 h | [56] |
| TBSV | p33 | eEF1A | S. cerevisiae | Peroxisomal membrane | Stabilization of p33 | Purified replicase proteomics | Protein–protein | S. cerevisiae | Co-purification | [35,57] |
| p33 | ESCRT-I | N. benthamiana | Peroxisomal membrane | Formation of replication compartments | Split ubiquitin assay | Protein–protein | S. cerevisiae | Confocal microscopy: 15–45 min | [58] | |
| p33 | GAPDH | N. benthamiana and S. cerevisiae | Peroxisomal membrane | Viral genomic RNA synthesis | Purified replicase proteomics | Indirect: mediated by p92pol | S. cerevisiae | Confocal microscopy: 16 h | [59] | |
| p33 and p92pol | HSP70 | S. cerevisiae | Peroxisomal membrane | Formation of replication compartments | Reconstitution assay | Protein–protein | S. cerevisiae | Confocal microscopy: 16 and 24 h | [60,61] | |
| p33 | ORP | S. cerevisiae | Peroxisome and ER | Formation of replication compartments | Affinity purification | Protein–protein | S. cerevisiae and N. benthamiana leaves | BiFC: 2 dpi | [52] | |
| TMV | RdRp and3′ UTR | eEF1A | N. tabacum | Replication compartment | Formation of replication compartments and cell-to-cell movement | Pull-down assay | Protein–protein | N. tabacum | Immunoprecipitation: 4 dpi | [62] |
| TuMV | VPg | AtRH8 | Prunus persica and A. thaliana | Chloroplast membrane | Formation of replication compartments | Yeast two hybrid | Protein–protein | N. benthamiana leaves | BiFC: 2 and 10 dpi | [63] |
| 6K2 | AtRH9 | A. thaliana | Chloroplast membrane | Formation of replication compartments | Confocal microcopy | Protein–protein | N. benthamiana leaves | Confocal microscopy: 72 h | [64] | |
| VPg and NIb | eEF1A | A. thaliana | ER-derived replication compartments | Viral RNA translation, formation of replication compartments | Tandem affinity purification | Protein–protein | N. benthamiana leaves | Immunofluorescence and confocal microscopy: 4–5 dpi | [65] | |
| VPg | eIF(iso)4e | A. thaliana | ER and chloroplasts | Viral RNA translation, formation of replication compartments | Pull-down assay | Protein–protein | N. benthamiana leaves | Immunofluorescence and confocal microscopy: 2–4 dpi | [65] | |
| TuMV | NIb | HSP70 | A. thaliana | Nucleus and replication compartments in the ER | Formation of replication compartments, regulation of NIb activity | Tandem affinity purification | Indirect: mediated by RdRp | N. benthamiana leaves | Confocal microscopy: 2–4 dpi | [20] |
| VPg | PABP2 | Brassica perviridis | Nucleus and ER | Formation of replication compartments | Subcellular fractionation | Protein–protein | N. benthamiana leaves | Confocal microscopy: 4–5 dpi | [66] | |
| ToMV | 130K and 180K | eEF1A | N. tabacum | ER membranes | Viral RNA translation, formation of replication compartments | Subcellular fractionation | Protein–protein | Transgenic N. tabacum BY-2 protoplast | Affinity purification | [67] |
| 130K and 180K | HSP70 | N. tabacum | ER membranes | Formation of replication compartments | Subcellular fractionation | Protein–protein | Transgenic N. tabacum BY-2 protoplast | Affinity purification | [67] | |
| TYLCV | CP | HSP70 | S. lycopersicum | Cytoplasm and nucleus aggregates | Movement of virions | Subcellular fractionation | Protein–protein | S. lycopersicum leaves | Immunodetection and confocal microscopy: 28 or 49 dpi | [68] |
| Endosomal proteins | ||||||||||
| CIRV | p36 | RAB5-GTPase | A. thaliana | Mitochondria | Formation of replication compartments | Yeast two hybrid | Protein–protein | N. benthamiana leaves | BiFC: 2 dpi | [69] |
| CNV | p33 | RAB5-GTPase | A. thaliana | Peroxisome | Formation of replication compartments | Yeast two hybrid | Protein–protein | N. benthamiana leaves | BiFC: 2 dpi | [69] |
| TBSV | p33 | RAB5-GTPase | A. thaliana | Peroxisome | Formation of replication compartments | Yeast two hybrid | Protein–protein | N. benthamiana leaves | BiFC: 2 dpi | [69] |
| Endoplasmic reticulum proteins | ||||||||||
| BMV | 1a | RHP | S. cerevisiae | Perinuclear ER membrane | Formation of replication compartments | Immunoprecipitation | Protein–protein | S. cerevisiae | Co-Ip and confocal microscopy: 12 dpi | [70] |
| TuMV | 6K2 | SNARE -SYP71 | A. thaliana | Chloroplast | Fusion replication compartments in chloroplast | Confocal microscopy | Indirect: mediated by Vap27-1 | N. benthamiana leaves | Confocal microscopy: 48 h | [71] |
| 6K2 | RHD3 | A. thaliana | Replication compartments | Maturation of replication compartments | Yeast two hybrid | Protein–protein | N. tabacum leaves | Confocal microscopy: 7 dpi | [28] | |
| Golgi apparatus proteins | ||||||||||
| BaMV | NA | RABG3f | N. benthamiana | Replication compartments | Formation and movement of replication compartments | Immunofluorescence | Unknown | N. benthamiana leaves | Confocal microscopy: 5 dpi | [72] |
| CaMV | MP | µA-adaptin | A. thaliana | Plasma membrane | MP trafficking | GST pull-down | Protein–protein | Escherichia coli and A. thaliana | GST-pull down | [73] |
| RCNMV | p27 | ARF1 | N. benthamiana and N. tabacum | ER | Formation of replication compartments | Affinity purification | Protein–protein | N. tabacum protoplast | Confocal microscopy: 16 h | [74] |
| Plasma membrane proteins | ||||||||||
| CaMV | p6 | AtSRC2.2 | A. thaliana | Inclusion bodies (cytoplasmic and nuclear) | Cell-to-cell movement | Yeast two hybrid | Protein–protein | N. benthamiana leaves | Co-immunoprecipitation and confocal microscopy: 3 dpi | [75] |
| RCNMV | p27 | RBOHB | N. benthamiana | Perinuclear ER-derived aggregates | ROS synthesis | Immunoprecipitation | Protein–protein | N. benthamiana leaves | Confocal microscopy andBiFC : 4 dpi | [76] |
| TVBMV | P3N-PIPO and CI | DREPP | N. benthamiana | Plasmodesmata | Cell-to-cell movement | Yeast two hybrid | Protein–protein | N. benthamiana leaves | BiFC: 2 and 5 dpi | [40] |
| Plasma membrane proteins | ||||||||||
| TVCV | MP | SYTA | A. thaliana | Plasmodesmata | Alters plasmodesmata permeability | Confocal microscopy | Protein–protein | N. benthamiana leaves | Confocal microscopy and BiFC | [34] |
| TuMV | P3N-PIPO | PCaP1 | N. benthamiana | Plasmodesmata | Cell-to-cell movement | Yeast two hybrid | Protein–protein | N. benthamiana leaves | BiFC: 38 h | [77] |
| Nuclear proteins | ||||||||||
| GRV | ORF3 | Fibrillarin | N. benthamiana and A. thaliana | Cytoplasm | Systemic movement | Affinity purification and chromatography | Protein–protein | N. benthamiana leaves and E. coli | Far Western blotting | [27] |
| RCNMV | p27 | HSP90 | N. benthamiana | ER | Formation of replication compartments | Partially purified replicase | Protein–protein | N. benthamiana leaves | BiFC: 3 and 4 dpi | [21] |
| TMV | MP | NTH201 | N. tabacum | Cytoplasm and plasmodesmata | Enhances replication compartment formation | Confocal microscopy | Indirect | N. benthamiana leaves | Confocal microscopy: 24 h | [78] |
| TBSV | p19 | ALY | N. benthamiana and A. thaliana | Cytoplasm | Co-factor | Yeast two hybrid | Protein–protein | N. benthamiana leaves | Confocal microscopy: 3 dpi | [79] |
| Vacuolar proteins | ||||||||||
| ToMV | 130K and 180K | TOM1 TOM3 | A. thaliana and N. tabacum | ER | Formation and anchoring of replication compartments | Membrane flotation | Protein–protein | S. cerevisiae and N. tabacum leaves | Yeast two hybrid and subcellular fractionation at 2 dpi | [80,81] |
| Virus | Viral Protein or RNA | Host Protein | Host | Movement of Host Protein into | Role | Initial Detection Experiment | Mechanism of Interaction | Experimental System for Detecting New Localization Sites * | Method of Observation: Time | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Cytoplasmic proteins | ||||||||||
| CaMV | P6 | NBR1 | A. thaliana | Nucleus | Inhibits salicylic acid-dependent defense responses | Confocal microscopy | Enhanced jasmonic acid signaling | Transgenic A. thaliana expressing 35S:NPR1-GFP leaves | Confocal microscopy: 5 to 40 min | [82] |
| LMV | HC-Pro | 20S α5 | A. thaliana | HC-Pro aggregates | Reduces RNase activity on viral RNA | Subcellular fractionation | Protein–protein | Lactuca sativa leaves | BiFC: 4 dpi | [83,84] |
| PVX | CP, TGBp1, or TGBp2 | MPB2Cb | N. benthamiana | ER | Blocks formation of replication compartments | Yeast two hybrid | Protein–protein | N. benthamiana leaves | Confocal microscopy: 2 dpi | [85] |
| Nuclear proteins | ||||||||||
| CIRV | p36 and p95pol | RH30 | N. benthamiana and A. thaliana | Mitochondria | Blocks assembly of the sites of replication | Confocal microscopy | Protein–protein | N. benthamiana leaves | Confocal microscopy: 84 h | [86] |
| CNV | p33 and p92pol | RH30 | N. benthamiana and A. thaliana | Peroxisome | Blocks assembly of the sites of replication | Confocal microscopy | Protein–protein | N. benthamiana leaves | Confocal microscopy: 84 h | [86] |
| TBSV | p33 and p92pol | RH30 | N. benthamiana and A. thaliana | Peroxisome | Blocks assembly of the sites of replication | Confocal microscopy | Protein–protein | N. benthamiana leaves | Confocal microscopy: 84 h | [86] |
| TBSV | p19 | ALY1 ALY3 | A. thaliana | Cytoplasm | Unknown | Yeast two hybrid | Protein–protein | N. benthamiana leaves | Confocal microscopy: 3 dpi | [79] |
| Vacuolar proteins | ||||||||||
| CaMV | p4 | NBR1 | A. thaliana | Inclusion bodies | NBR1-dependent degradation of p4 | Yeast two hybrid | Protein–protein | N. benthamiana leaves | Confocal microscopy: 2 dpi | [82] |
| TuMV | HC-Pro | NBR1 | A. thaliana | Granule-like cytoplasmic structures | NBR1-dependent degradation of HC-Pro | Confocal microscopy | Protein–protein | Transgenic A. thaliana expressing NBR1-RFP leaves | Confocal microscopy on systemically infected leaves: 14 dpi | [87] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez-Peña, R.; Mounadi, K.E.; Garcia-Ruiz, H. Changes in Subcellular Localization of Host Proteins Induced by Plant Viruses. Viruses 2021, 13, 677. https://doi.org/10.3390/v13040677
Rodriguez-Peña R, Mounadi KE, Garcia-Ruiz H. Changes in Subcellular Localization of Host Proteins Induced by Plant Viruses. Viruses. 2021; 13(4):677. https://doi.org/10.3390/v13040677
Chicago/Turabian StyleRodriguez-Peña, Rosalba, Kaoutar El Mounadi, and Hernan Garcia-Ruiz. 2021. "Changes in Subcellular Localization of Host Proteins Induced by Plant Viruses" Viruses 13, no. 4: 677. https://doi.org/10.3390/v13040677
APA StyleRodriguez-Peña, R., Mounadi, K. E., & Garcia-Ruiz, H. (2021). Changes in Subcellular Localization of Host Proteins Induced by Plant Viruses. Viruses, 13(4), 677. https://doi.org/10.3390/v13040677