
Isolation and Characterization of Streptococcus mutans Phage as a Possible Treatment Agent for Caries

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Growth Conditions
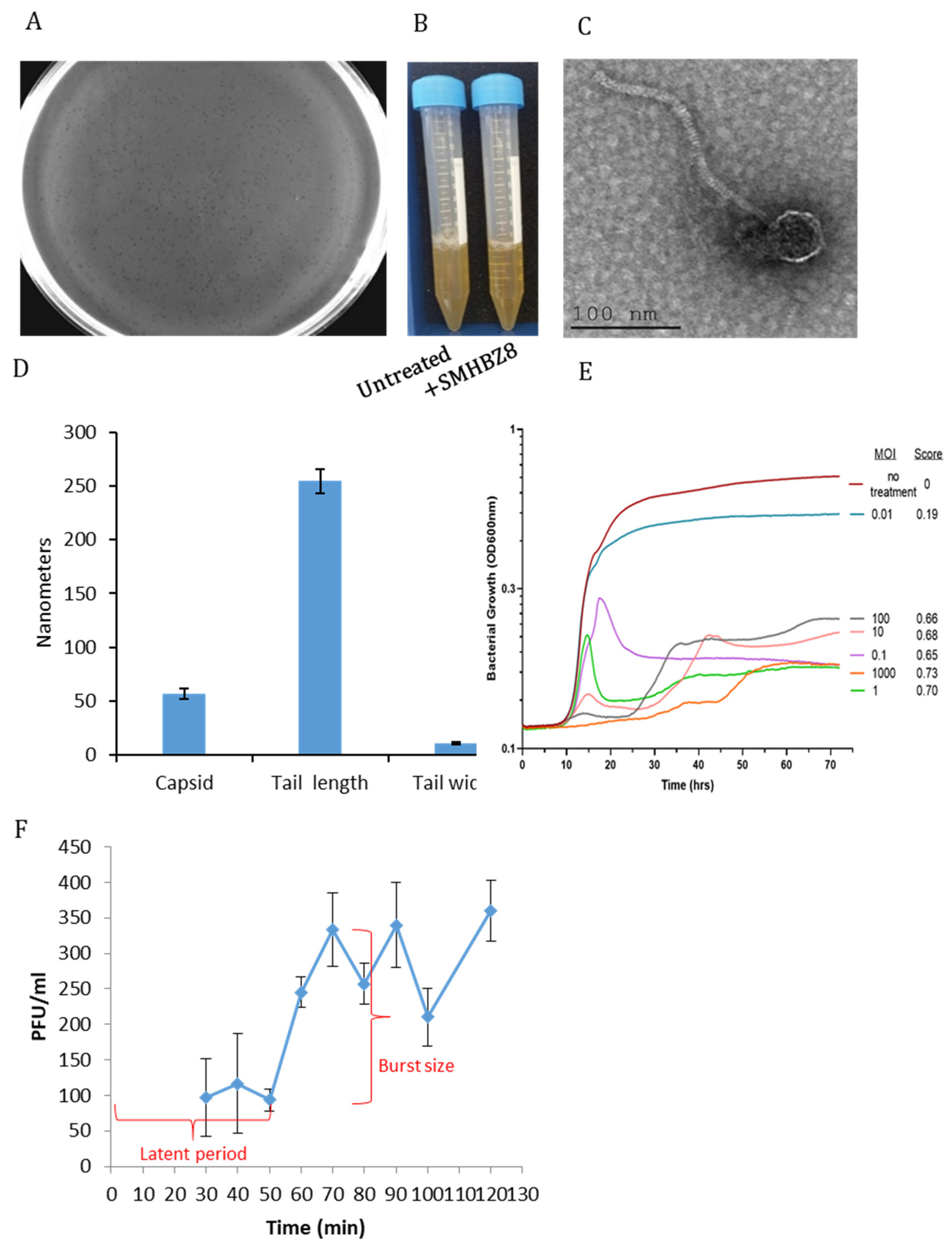
2.2. One Step Growth
2.3. Sample Collection
2.4. Phage Isolation and Propagation
2.5. TEM Analysis of the Phage
2.6. DNA Isolation and Sequencing
2.7. Determination of the Phage Lytic Activity in Planktonic Culture
2.8. Determination of Phage Activity on Biofilm Formation
2.9. Host Range Specificity Tests
2.10. Effect of pH on Phage Activity
2.11. SMHBZ8 Effect on Cariogenic Dentin
2.12. Statistical Analysis
3. Results
3.1. Isolation of SMHBZ8 and Testing Its Efficiency against S. mutans
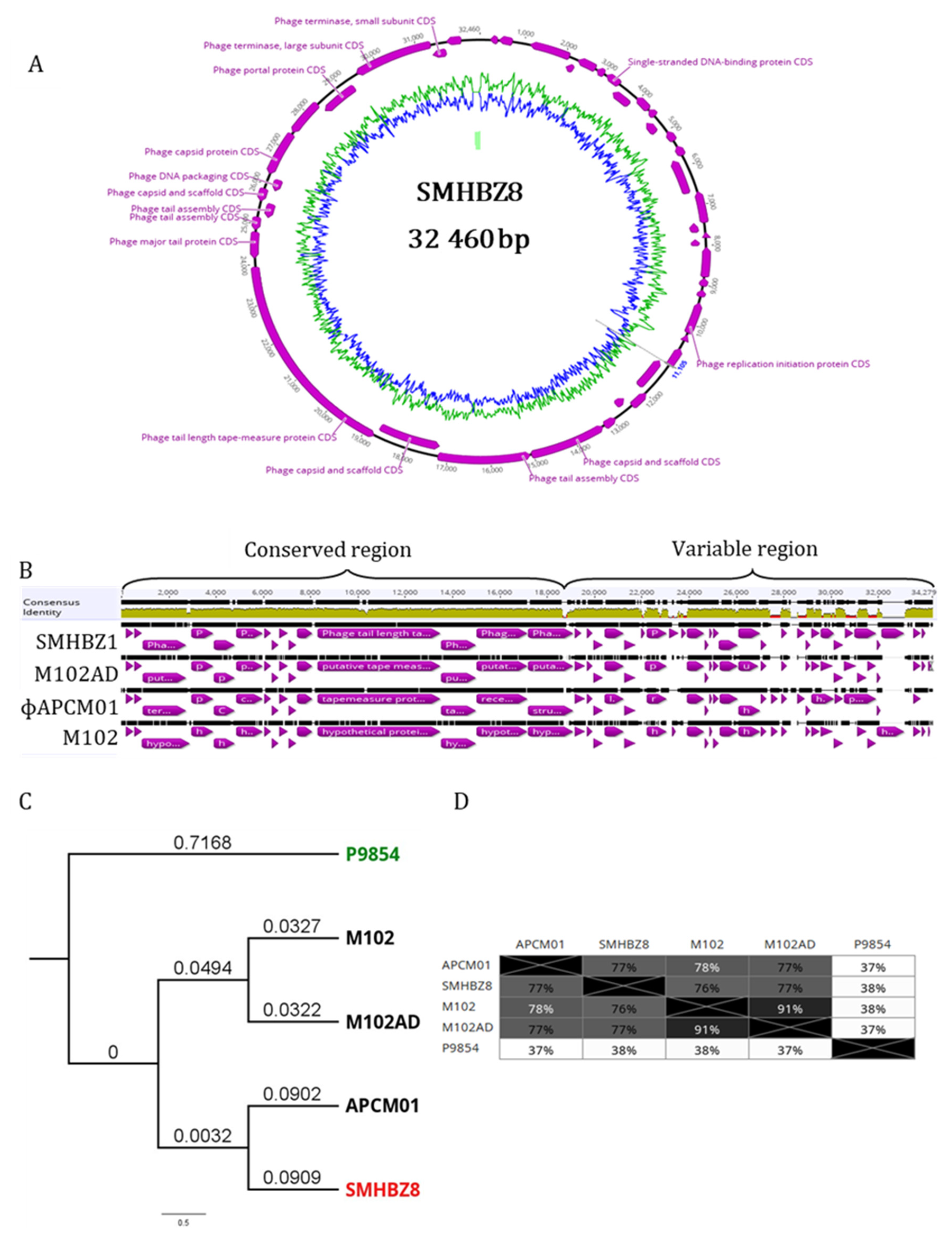
3.2. Whole-Genome Sequencing, and Phylogeny of SMHBZ8
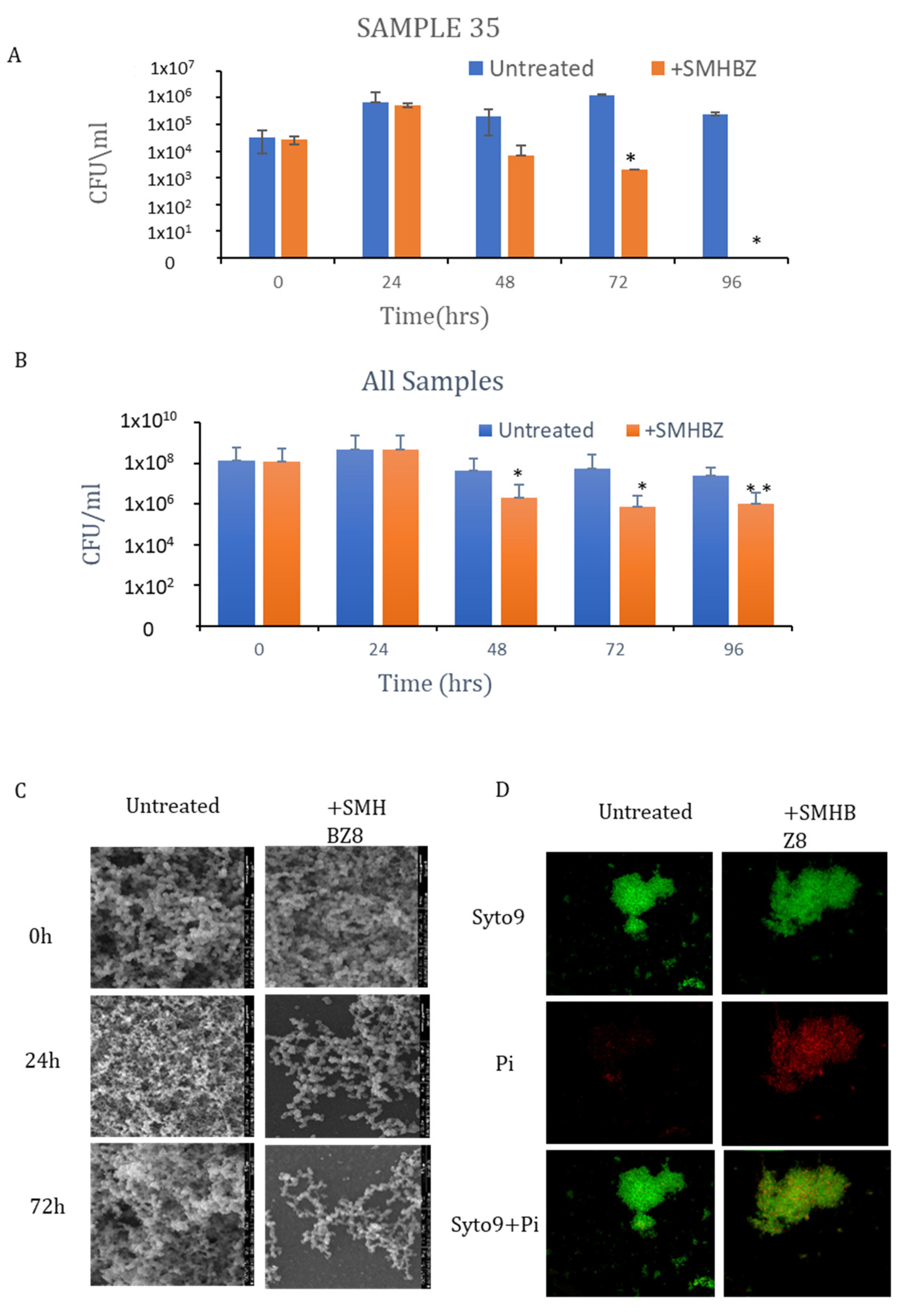
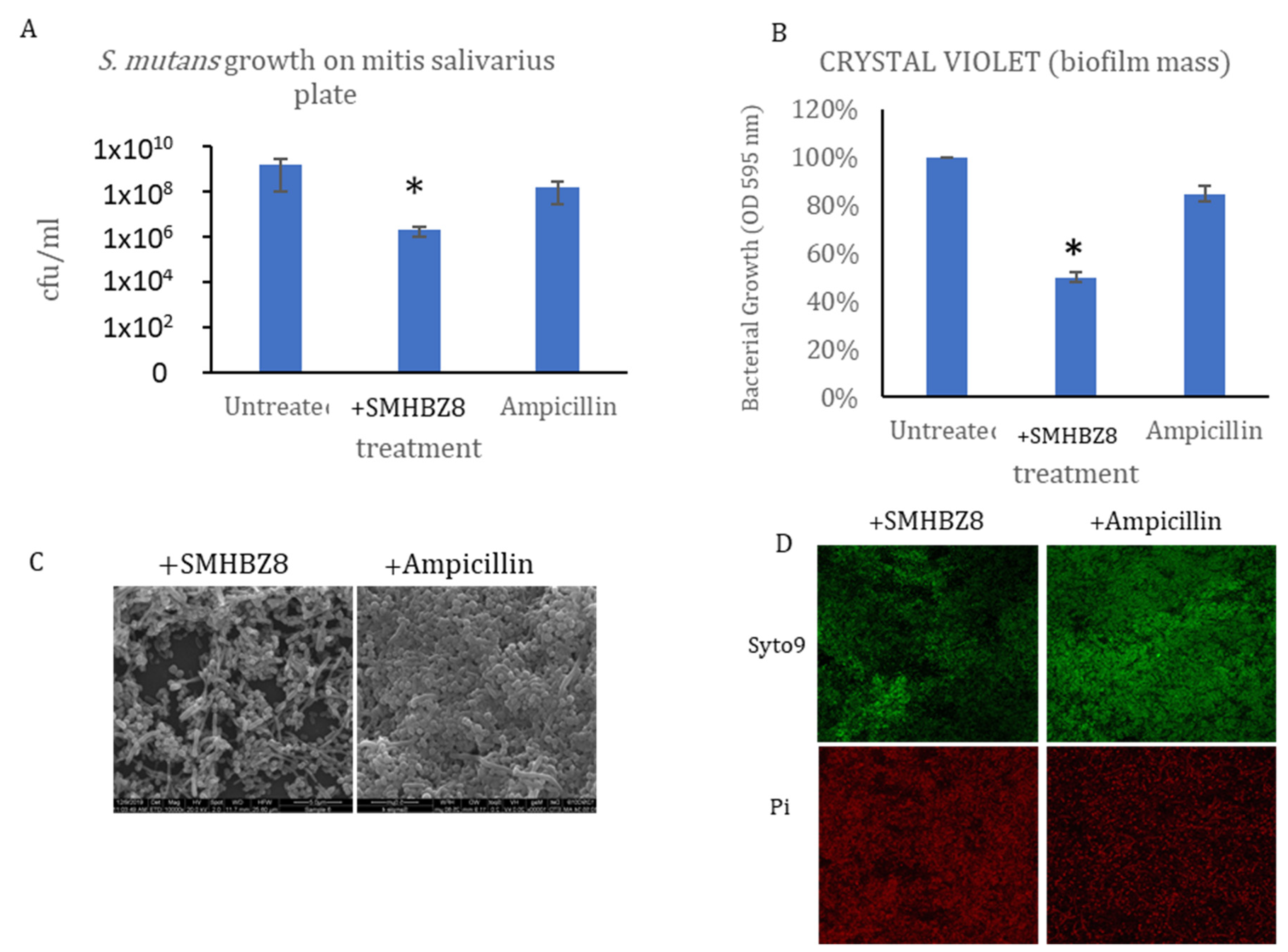
3.3. SMHBZ8 Activity on S. mutans Biofilm Formation
3.4. SMHBZ8 Range of Infectivity Tests
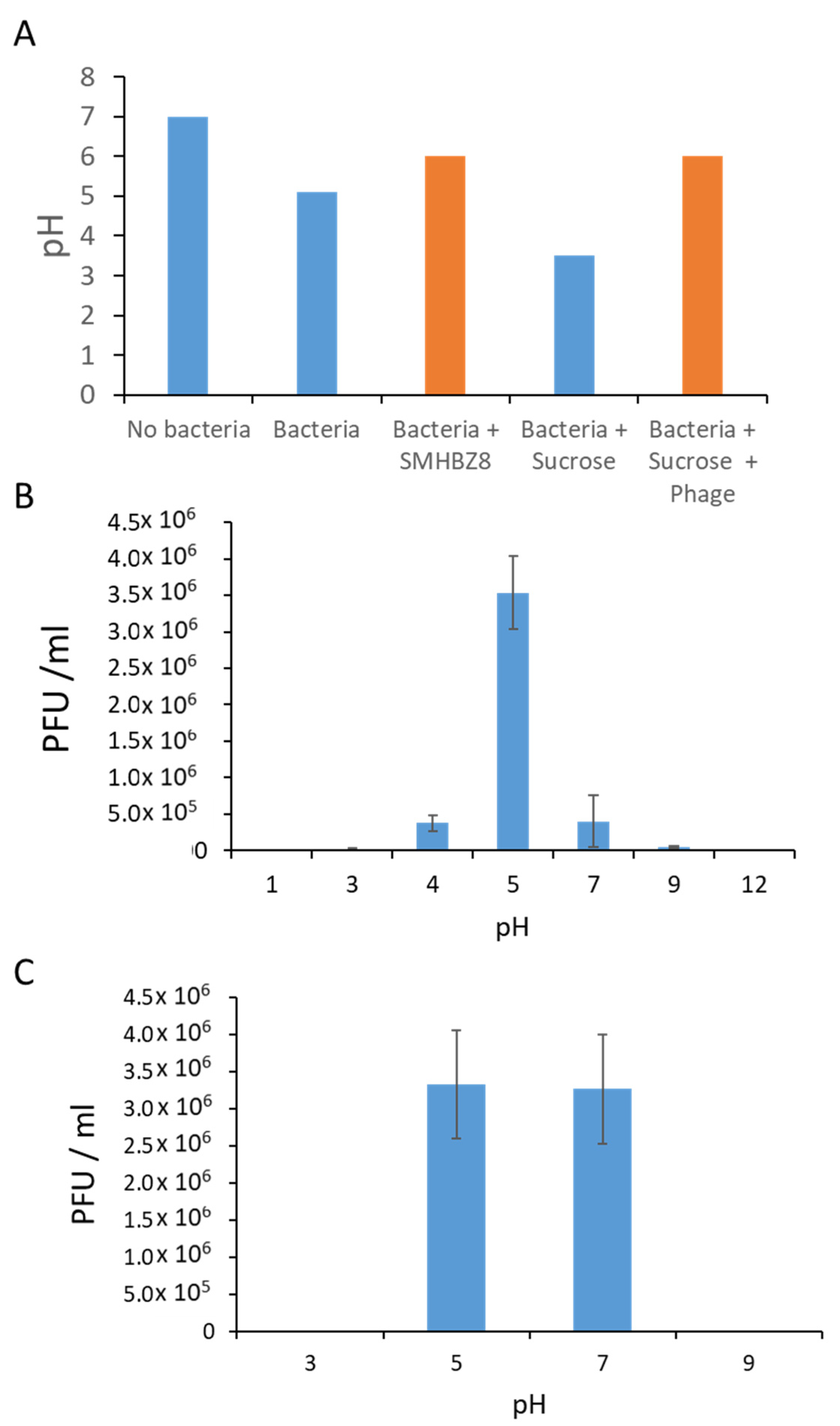
3.5. Phage pH Properties
3.6. SMHBZ8 Effect on Cariogenic Dentin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kutsch, V.K. Dental caries: An updated medical model of risk assessment. J. Prosthet. Dent. 2014, 111, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Kassebaum, N.J.; Bernabe, E.; Dahiya, M.; Bhandari, B.; Murray, C.J.; Marcenes, W. Global Burden of Untreated Caries: A Systematic Review and Metaregression. J. Dent. Res. 2015. [Google Scholar] [CrossRef]
- Yon, M.J.Y.; Gao, S.S.; Chen, K.J.; Duangthip, D.; Lo, E.C.M.; Chu, C.H. Medical Model in Caries Management. Dent. J. 2019, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpiński, T.M.; Szkaradkiewicz, A.K. Microbiology of dental caries. J. Biol. Earth Sci. 2013, 3, M21–M24. [Google Scholar]
- Yip, K.; Smales, R. Oral diagnosis and treatment planning: Part 2. Dental caries and assessment of risk. Br. Dent. J. 2012, 213, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, S.B. Oral Streptococci Bacteriophages as a Potential Agent for Dental Caries Therapy. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 512–515. [Google Scholar]
- Motegi, M.; Takagi, Y.; Yonezawa, H.; Hanada, N.; Terajima, J.; Watanabe, H.; Senpuku, H. Assessment of genes associated with Streptococcus mutans biofilm morphology. Appl Environ. Microbiol. 2006, 72, 6277–6287. [Google Scholar] [CrossRef] [Green Version]
- Nakano, K.; Ooshima, T. Serotype classification of Streptococcus mutans and its detection outside the oral cavity. Future Microbiol. 2009, 4, 891–902. [Google Scholar] [CrossRef]
- Shibata, Y.; Ozaki, K.; Seki, M.; Kawato, T.; Tanaka, H.; Nakano, Y.; Yamashita, Y. Analysis of loci required for determination of serotype antigenicity in Streptococcus mutans and its clinical utilization. J. Clin. Microbiol. 2003, 41, 4107–4112. [Google Scholar] [CrossRef] [Green Version]
- National Institutes of Health. Diagnosis and management of dental caries throughout life. NIH Consens. Statement 2001, 18, 1–23. [Google Scholar]
- Mathur, V.P.; Dhillon, J.K. Dental Caries: A Disease Which Needs Attention. Indian J. Pediatrics 2018, 85, 202–206. [Google Scholar] [CrossRef]
- Salli, K.M.; Ouwehand, A.C. The use of in vitro model systems to study dental biofilms associated with caries: A short review. J. Oral Microbiol. 2015, 7, 26149. [Google Scholar] [CrossRef]
- Khalifa, L.; Shlezinger, M.; Beyth, S.; Houri-Haddad, Y.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Phage therapy against Enterococcus faecalis in dental root canals. J. Oral Microbiol. 2016, 8, 32157. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ren, Z.; Hwang, G.; Koo, H. Therapeutic Strategies Targeting Cariogenic Biofilm Microenvironment. Adv. Dent. Res. 2018, 29, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Rugg-Gunn, A. Dental caries: Strategies to control this preventable disease. Acta Med. Acad. 2013, 42, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Forssten, S.D.; Bjorklund, M.; Ouwehand, A.C. Streptococcus mutans, caries and simulation models. Nutrients 2010, 2, 290–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karygianni, L.; Al-Ahmad, A.; Argyropoulou, A.; Hellwig, E.; Anderson, A.C.; Skaltsounis, A.L. Natural Antimicrobials and Oral Microorganisms: A Systematic Review on Herbal Interventions for the Eradication of Multispecies Oral Biofilms. Front. Microbiol. 2015, 6, 1529. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.I.; Hasson, H.; Sohn, W. Dental caries in the second millennium. J. Dent. Educ. 2001, 65, 953–959. [Google Scholar] [CrossRef]
- Bowen, W.H. Dental caries—Not just holes in teeth! A perspective. Mol. Oral Microbiol. 2016, 31, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Poorni, S.; Srinivasan, M.R.; Nivedhitha, M.S. Probiotic Streptococcus strains in caries prevention: A systematic review. J. Conserv. Dent. 2019, 22, 123–128. [Google Scholar] [CrossRef]
- Wittebole, X.; De Roock, S.; Opal, S.M. A historical overview of bacteriophage therapy as an alternative to antibiotics for the treatment of bacterial pathogens. Virulence 2014, 5, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, S.B. Bacteriophage Therapy: A possible new alternative for oral diseases. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 437–442. [Google Scholar]
- Rehman, S.; Ali, Z.; Khan, M.; Bostan, N.; Naseem, S. The dawn of phage therapy. Rev. Med Virol. 2019, e2041. [Google Scholar] [CrossRef]
- Dublanchet, A.; Bourne, S. The epic of phage therapy. Can. J. Infect. Dis. Med Microbiol. J. Can. Des Mal. Infect. Microbiol. Med. 2007, 18, 15–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, L.; Brosh, Y.; Gelman, D.; Coppenhagen-Glazer, S.; Beyth, S.; Poradosu-Cohen, R.; Que, Y.A.; Beyth, N.; Hazan, R. Targeting Enterococcus faecalis biofilms with phage therapy. Appl Environ. Microbiol. 2015, 81, 2696–2705. [Google Scholar] [CrossRef] [Green Version]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Pinto, G.; Silva, M.D.; Peddey, M.; Sillankorva, S.; Azeredo, J. The role of bacteriophages in periodontal health and disease. Future Microbiol. 2016, 11, 1359–1369. [Google Scholar] [CrossRef]
- Fruciano, D.E.; Bourne, S. Phage as an antimicrobial agent: D’Herelle’s heretical theories and their role in the decline of phage prophylaxis in the West. Can. J. Infect. Dis. Med Microbiol. J. Can. Des Mal. Infect. Microbiol. Med. 2007, 18, 19–26. [Google Scholar] [CrossRef]
- Hyman, P.; Abedon, S.T. Chapter 7—Bacteriophage Host Range and Bacterial Resistance. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2010; Volume 70, pp. 217–248. [Google Scholar]
- Chen, Y.; Batra, H.; Dong, J.; Chen, C.; Rao, V.B.; Tao, P. Genetic Engineering of Bacteriophages Against Infectious Diseases. Front. Microbiol. 2019, 10, 954. [Google Scholar] [CrossRef]
- Fauconnier, A. Phage Therapy Regulation: From Night to Dawn. Viruses 2019, 11, 352. [Google Scholar] [CrossRef] [Green Version]
- Nir-Paz, R.; Gelman, D.; Khouri, A.; Sisson, B.M.; Fackler, J.; Alkalay-Oren, S.; Khalifa, L.; Rimon, A.; Yerushalmy, O.; Bader, R.; et al. Successful Treatment of Antibiotic-resistant, Poly-microbial Bone Infection With Bacteriophages and Antibiotics Combination. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2019, 69, 2015–2018. [Google Scholar] [CrossRef]
- van der Ploeg, J.R. Analysis of CRISPR in Streptococcus mutans suggests frequent occurrence of acquired immunity against infection by M102-like bacteriophages. Microbiology 2009, 155, 1966–1976. [Google Scholar] [CrossRef] [Green Version]
- Maal, K.; Bouzari, M.; Arbabzadeh, F. Identification of Streptococcus Salivarius Bacteriophage Isolated from Persian Gulf as a Potential Agent for Dental Caries Phage Therapy. Afr. J. Microbiol. Res. 2010, 4, 2127–2132. [Google Scholar]
- Hitch, G.; Pratten, J.; Taylor, P.W. Isolation of bacteriophages from the oral cavity. Lett. Appl. Microbiol. 2004, 39, 215–219. [Google Scholar] [CrossRef]
- Bachrach, G.; Leizerovici-Zigmond, M.; Zlotkin, A.; Naor, R.; Steinberg, D. Bacteriophage isolation from human saliva. Lett. Appl. Microbiol. 2003, 36, 50–53. [Google Scholar] [CrossRef]
- van der Ploeg, J.R. Genome sequence of Streptococcus mutans bacteriophage M102. FEMS Microbiol. Lett. 2007, 275, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delisle, A.L.; Guo, M.; Chalmers, N.I.; Barcak, G.J.; Rousseau, G.M.; Moineau, S. Biology and genome sequence of Streptococcus mutans phage M102AD. Appl. Environ. Microbiol. 2012, 78, 2264–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmasso, M.; de Haas, E.; Neve, H.; Strain, R.; Cousin, F.J.; Stockdale, S.R.; Ross, R.P.; Hill, C. Isolation of a Novel Phage with Activity against Streptococcus mutans Biofilms. PLoS ONE 2015, 10, e0138651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, O.G.; Jordan, H.V.; van Houte, J. A selective medium for Streptococcus mutans. Arch. Oral Biol. 1973, 18, 1357–1364. [Google Scholar] [CrossRef]
- Ajdic, D.; McShan, W.M.; McLaughlin, R.E.; Savic, G.; Chang, J.; Carson, M.B.; Primeaux, C.; Tian, R.; Kenton, S.; Jia, H.; et al. Genome sequence of Streptococcus mutans UA159, a cariogenic dental pathogen. Proc. Natl. Acad. Sci. USA 2002, 99, 14434–14439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtonen, O.-P.; Gråhn, E.; Ståhlberg, T.; Laitinen, L.A. Amount and avidity of salivary and serum antibodies against Streprococcus mutans in two groups of human subjects with different dental susceptibility. Infect. Immun. 1984, 43, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, L.; Gelman, D.; Shlezinger, M.; Dessal, A.L.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Defeating Antibiotic- and Phage-Resistant Enterococcus faecalis Using a Phage Cocktail in Vitro and in a Clot Model. Front. Microbiol. 2018, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Alkalay, S.; Sternberg, S.; Coppenhagen-Glazer, S.; Hazan, R. Complete Genome Sequences of Three Bacillus anthracis Bacteriophages. Genome Announc. 2018, 6, e01164-17. [Google Scholar] [CrossRef] [Green Version]
- OpenWetWare. Gill: Preparing Phage Specimens for TEM. Available online: https://openwetware.org/wiki/Gill:Preparing_phage_specimens_for_TEM (accessed on 22 September 2019).
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Kot, W.; Vogensen, F.K.; Sorensen, S.J.; Hansen, L.H. DPS—A rapid method for genome sequencing of DNA-containing bacteriophages directly from a single plaque. J. Virol. Methods 2014, 196, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Eini, A.; Sol, A.; Coppenhagen-Glazer, S.; Skvirsky, Y.; Zini, A.; Bachrach, G. Oxygen deprivation affects the antimicrobial action of LL-37 as determined by microplate real-time kinetic measurements under anaerobic conditions. Anaerobe 2013, 22, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Storms, Z.J.; Teel, M.R.; Mercurio, K.; Sauvageau, D. The virulence index: A metric for quantitative analysis of phage virulence. Phage 2020, 1, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Emilson, C.G.; Bratthall, D. Growth of Streptococcus mutans on various selective media. J. Clin. Microbiol. 1976, 4, 95–98. [Google Scholar] [PubMed]
- Baker, C.N.; Thornsberry, C. Antimicrobial susceptibility of Streptococcus mutans isolated from patients with endocarditis. Antimicrob. Agents Chemother. 1974, 5, 268–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, H.-W. Tailed Bacteriophages: The Order Caudovirales. In Advances in Virus Research; Maramorosch, K., Murphy, F.A., Shatkin, A.J., Eds.; Academic Press: Cambridge, MA, USA, 1998; Volume 51, pp. 135–201. [Google Scholar]
- Fokine, A.; Rossmann, M.G. Molecular architecture of tailed double-stranded DNA phages. Bacteriophage 2014, 4, e28281. [Google Scholar] [CrossRef] [Green Version]
- Elbreki, M.; Ross, R.P.; Hill, C.; O’Mahony, J.; McAuliffe, O.; Coffey, A. Bacteriophages and Their Derivatives as Biotherapeutic Agents in Disease Prevention and Treatment. J. Viruses 2014, 2014, 382539. [Google Scholar] [CrossRef] [Green Version]
- Sillankorva, S.; Oliveira, R.; Vieira, M.J.; Sutherland, I.; Azeredo, J. Bacteriophage Φ S1 Infection of Pseudomonas fluorescens Planktonic Cells versus Biofilms. Biofouling 2004, 20, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Abedon, S.T. Ecology of Anti-Biofilm Agents II: Bacteriophage Exploitation and Biocontrol of Biofilm Bacteria. Pharmaceuticals 2015, 8, 559–589. [Google Scholar] [CrossRef] [Green Version]
- Stephan, R.M.; Miller, B.F. A Quantitative Method for Evaluating Physical and Chemical Agents which Modify Production of Acids in Bacterial Plaques on Human Teeth. J. Dent. Res. 1943, 22, 45–51. [Google Scholar] [CrossRef]
- Shlezinger, M.; Khalifa, L.; Houri-Haddad, Y.; Coppenhagen-Glazer, S.; Resch, G.; Que, Y.A.; Beyth, S.; Dorfman, E.; Hazan, R.; Beyth, N. Phage Therapy: A New Horizon in the Antibacterial Treatment of Oral Pathogens. Curr. Top. Med. Chem. 2017, 17, 1199–1211. [Google Scholar] [CrossRef]
- Steier, L.; de Oliveira, S.D.; de Figueiredo, J.A.P. Bacteriophages in Dentistry-State of the Art and Perspectives. Dent. J. 2019, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.; Abedon, S.T. Phage therapy pharmacology phage cocktails. Adv. Appl. Microbiol. 2012, 78, 1–23. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Broth | Incubation Conditions |
|---|---|---|
| Streptococcus strains | ||
| Streptococcus sobrinus (lsb013) | BHI | 37 °C |
| Streptococcus salivarius | BHI | 37 °C |
| Streptococcus gordonii | BHI | 37 °C |
| Other strains: | ||
| E. faecalis V583 | BHI | 37 °C, 200 rpm shaking |
| Pseudomonas aeruginosa PA14 R | LB | 37 °C, 200 rpm shaking |
| Klebsiella pneumonia (bkp016) R | LB | 37 °C, 200 rpm shaking |
| Actinomyces viscosus | BHI | 37 °C |
| Fusobacterium nucleatum (fs014) | LB | 37 °C, anaerobic |
| Type | Number of Genes |
|---|---|
| Core | 7 |
| Soft core including HMBZ genes | 1 |
| Soft core without HMBZ | 3 |
| Conserved in APCM01 and M102 | 1 |
| Conserved in M102 and M102AD | 13 |
| SMHBZ8 unique | 32 |
| APCM01 unique | 25 |
| M102 unique | 14 |
| M102AD unique | 14 |
| Bacterial Strain | Origin | SMHBZ1 | Serotype |
|---|---|---|---|
| S. mutans strains | |||
| S. mutans (700610) | ATCC | S | c |
| S. mutans (27351) | ATCC | S | c |
| S. mutans (ES1) | Clinically isolated from saliva | S | c |
| S. mutans (ES2) | Clinically isolated from saliva | S | c |
| S. mutans (ES3) | Clinically isolated from saliva | S | c |
| S. mutans (ES4) | Clinically isolated from saliva | S | c |
| S. mutans (ES5) | Clinically isolated from saliva | S | c |
| S. mutans (ES6) | Clinically isolated from saliva | S | c |
| S. mutans (ES7) | Clinically isolated from saliva | S | c |
| S. mutans (ES8) | Clinically isolated from saliva | S | c |
| S. mutans (ES9) | Clinically isolated from saliva | S | c |
| S. mutans (ES10) | Clinically isolated from saliva | S | c |
| S. mutans (MT8148) | Kyushu University | S | c |
| S. mutans (MT703) | Kyushu University | R | e |
| S. mutans (LM7) | Kyushu University | R | e |
| S. mutans (OMZ175) | Kyushu University | R | f |
| S. mutans (M76219) | Kyushu University | R | f |
| Other Streptococcus strains | |||
| Streptococcus sobrinus (lsb013) | ATCC | R | |
| Streptococcus salivarius | ATCC | R | |
| Streptococcus gordonii | ATCC | R | |
| Other strains | |||
| E. faecalis V583 | ATCC | R | |
| Pseudomonas aeruginosa PA14 R | ATCC | R | |
| Klebsiella pneumonia (bkp016) R | ATCC | R | |
| Actinomyces viscosus | Clinically isolated | R | |
| Fusobacterium nucleatum (fs014) | ATCC | R | |
| Staphylococcus aureus | Clinically isolated | R | |
| Escherichia coli | ATCC | R | |
| Streptococcus salivarius | ATCC | R | |
| Streptococcus gordonii | ATCC | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben-Zaken, H.; Kraitman, R.; Coppenhagen-Glazer, S.; Khalifa, L.; Alkalay-Oren, S.; Gelman, D.; Ben-Gal, G.; Beyth, N.; Hazan, R. Isolation and Characterization of Streptococcus mutans Phage as a Possible Treatment Agent for Caries. Viruses 2021, 13, 825. https://doi.org/10.3390/v13050825
Ben-Zaken H, Kraitman R, Coppenhagen-Glazer S, Khalifa L, Alkalay-Oren S, Gelman D, Ben-Gal G, Beyth N, Hazan R. Isolation and Characterization of Streptococcus mutans Phage as a Possible Treatment Agent for Caries. Viruses. 2021; 13(5):825. https://doi.org/10.3390/v13050825
Chicago/Turabian StyleBen-Zaken, Hadar, Reut Kraitman, Shunit Coppenhagen-Glazer, Leron Khalifa, Sivan Alkalay-Oren, Daniel Gelman, Gilad Ben-Gal, Nurit Beyth, and Ronen Hazan. 2021. "Isolation and Characterization of Streptococcus mutans Phage as a Possible Treatment Agent for Caries" Viruses 13, no. 5: 825. https://doi.org/10.3390/v13050825
APA StyleBen-Zaken, H., Kraitman, R., Coppenhagen-Glazer, S., Khalifa, L., Alkalay-Oren, S., Gelman, D., Ben-Gal, G., Beyth, N., & Hazan, R. (2021). Isolation and Characterization of Streptococcus mutans Phage as a Possible Treatment Agent for Caries. Viruses, 13(5), 825. https://doi.org/10.3390/v13050825