
Common Polymorphisms in the Glycoproteins of Human Cytomegalovirus and Associated Strain-Specific Immunity

 , , ,
, , ,
Abstract
:1. Introduction
2. Complexity of HCMV Genomes and Viral Populations In Vivo
3. Common Polymorphisms in the Glycoproteins of HCMV
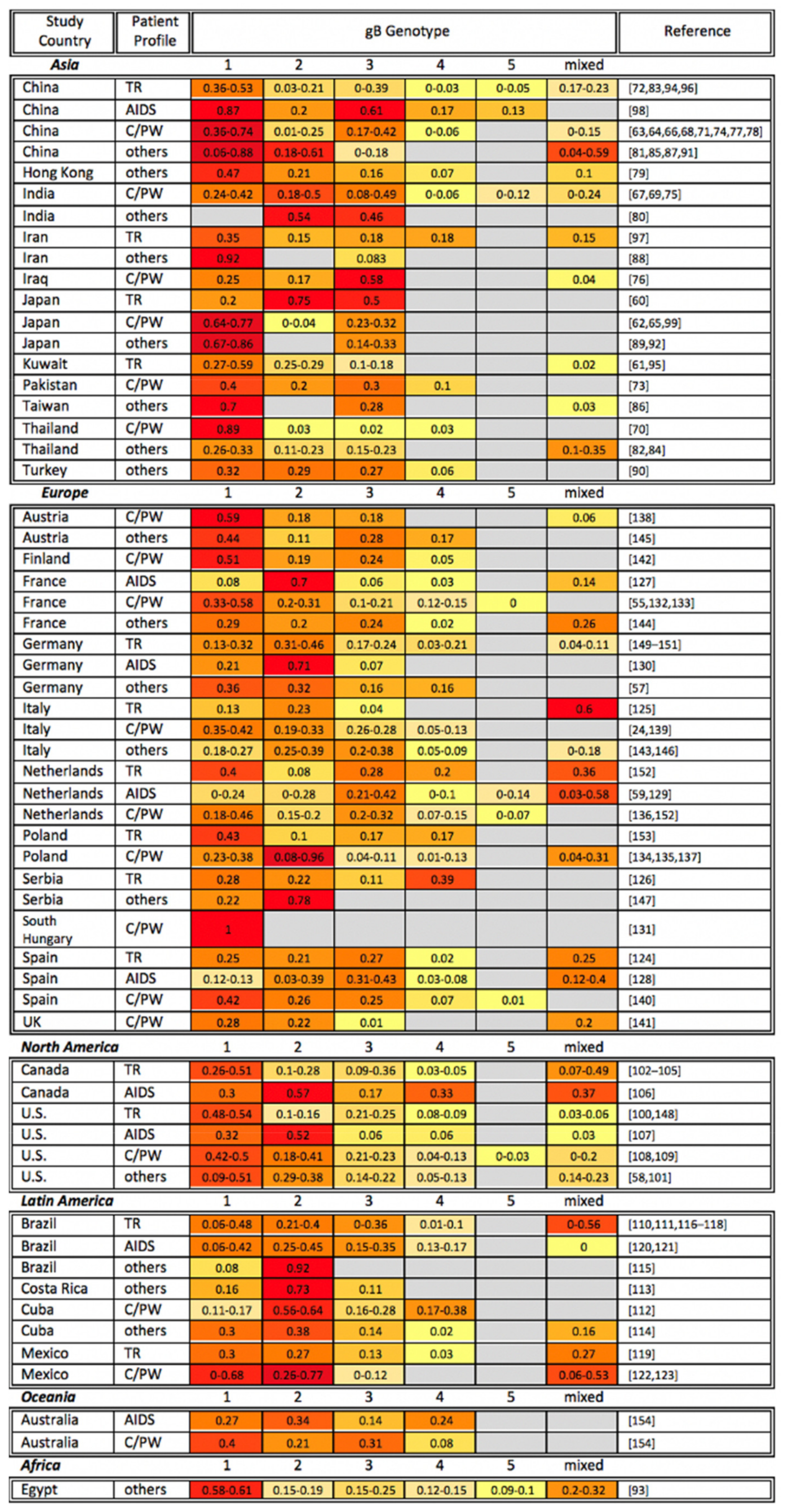
4. gB

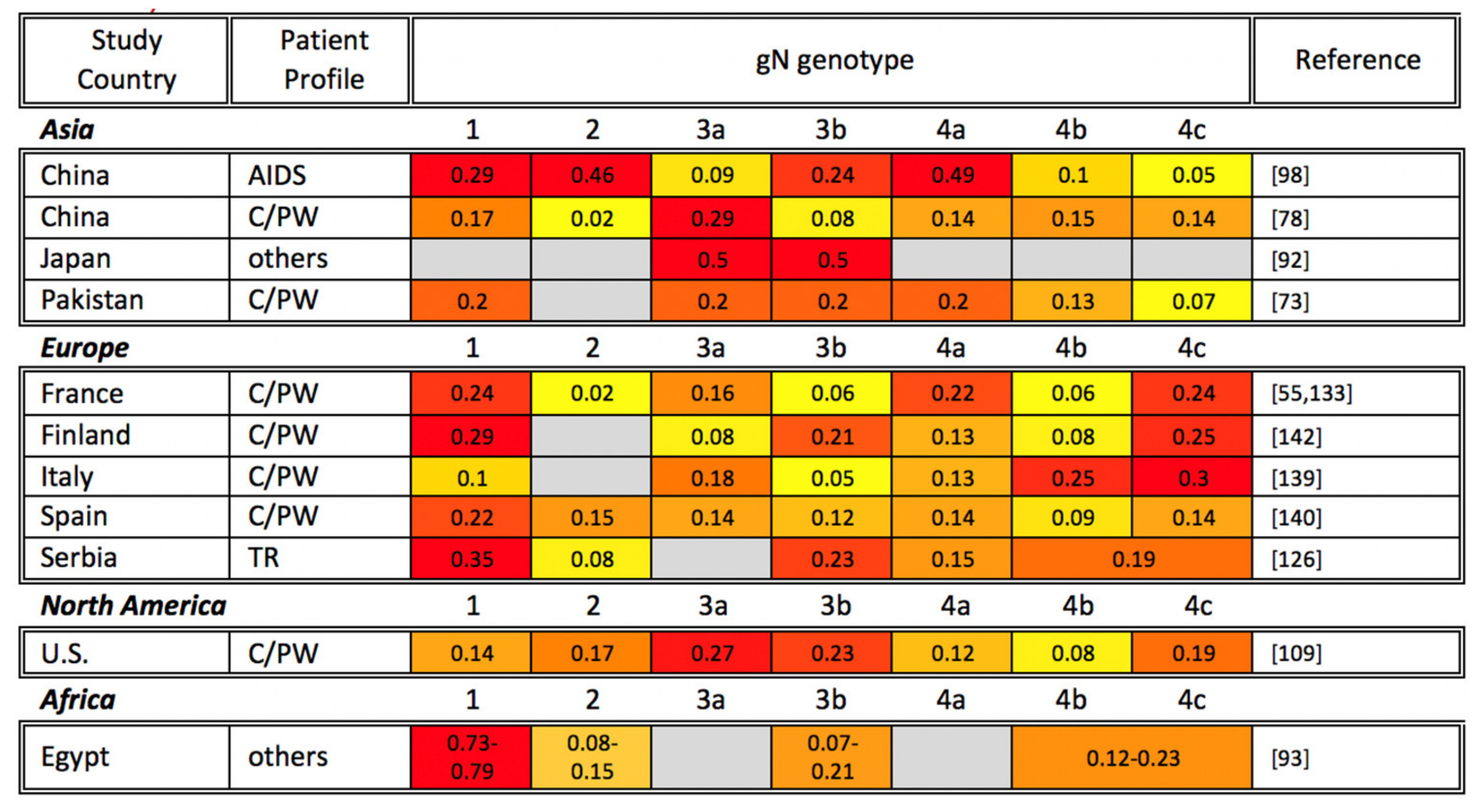
5. gN
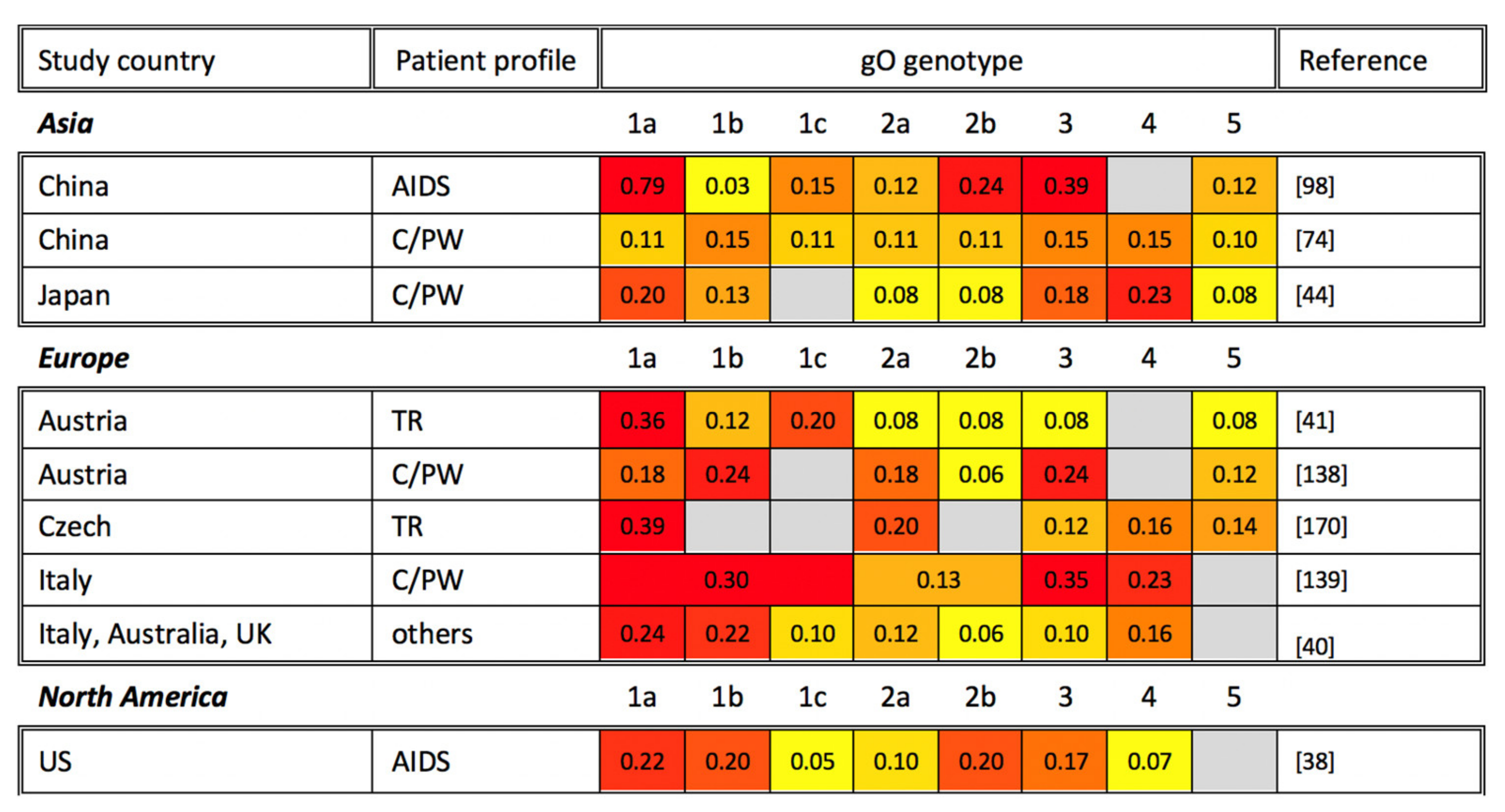
6. gO
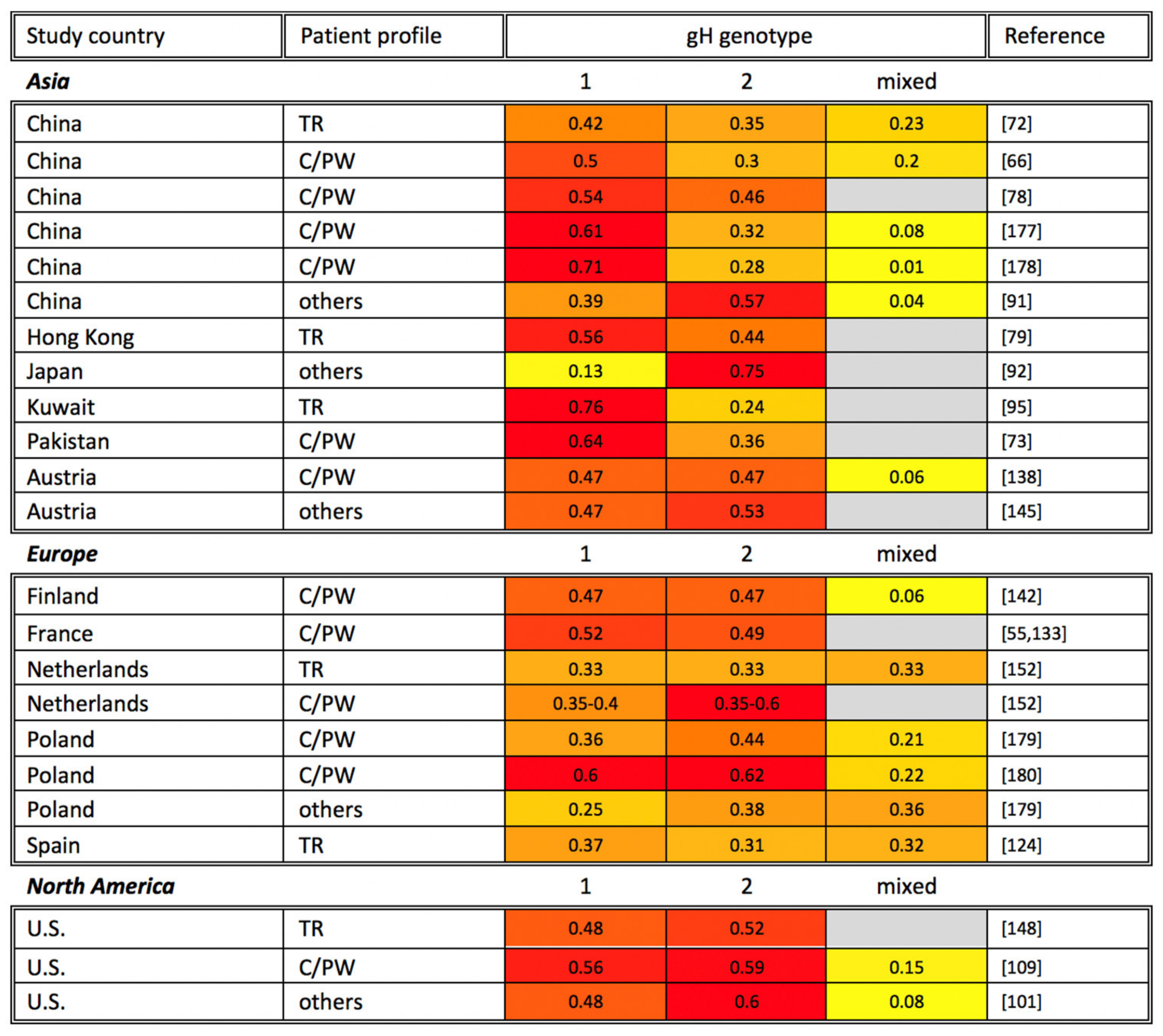
7. gH
8. Concluding Remarks
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuhair, M.; Smit, G.S.A.; Wallis, G.; Jabbar, F.; Smith, C.; Devleesschauwer, B.; Griffiths, P. Estimation of the worldwide seroprevalence of cytomegalovirus: A systematic review and meta-analysis. Rev. Med. Virol. 2019, 29, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Emery, V.C. Investigation of CMV disease in immunocompromised patients. J. Clin. Pathol. 2001, 54, 84–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, S.W. Reactivation and recombination of multiple cytomegalovirus strains from individual organ donors. J. Infect. Dis. 1989, 160, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Sackman, A.M.; Pfeifer, S.P.; Kowalik, T.F.; Jensen, J.D. On the demographic and selective forces shaping patterns of human cytomegalovirus variation within hosts. Pathogens 2018, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Huang, E.S.; Huong, S.M.; Tegtmeier, G.E.; Alford, C. Cytomegalovirus: Genetic variation of viral genomes. Ann. N. Y. Acad. Sci. 1980, 354, 332–346. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.; Geissler, A.; Winters, M. Inter- and Intragenic Variations Complicate the Molecular Epidemiology of Human Cytomegalovirus. J. Infect. Dis. 2003, 187, 809–819. [Google Scholar] [CrossRef]
- Plotkin, S.A.; Furukawa, T.; Zygraich, N.; Huygelen, C. Candidate cytomegalovirus strain for human vaccination. Infect. Immun. 1975, 12, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, S.A.; Friedman, H.M.; Fleisher, G.R.; Dafoe, D.C.; Grossman, R.A.; Lynn Smiley, M.; Starr, S.E.; Wlodaver, C.; Friedman, A.D.; Barker, C.F. Towne-vaccine-induced prevention of cytomegalovirus disease after renal transplants. Lancet 1984, 323, 528–530. [Google Scholar] [CrossRef]
- Chee, M.S.; Bankier, A.T.; Beck, S.; Bohni, R.; Brown, C.M.; Cerny, R.; Horsnell, T.; Hutchison, C.A.; Kouzarides, T.; Martignetti, J.A.; et al. Analysis of the protein-coding content of the sequence of human cytomegalovirus strain AD169. In Proceedings of the Cytomegaloviruses; McDougall, J.K., Ed.; Springer: Berlin/Heidelberg, Germany, 1990; pp. 125–169. [Google Scholar]
- Renzette, N.; Pfeifer, S.P.; Matuszewski, S.; Kowalik, T.F.; Jensen, J.D. On the analysis of intrahost and interhost viral populations: Human cytomegalovirus as a case study of pitfalls and expectations. J. Virol. 2017, 91, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sinzger, C.; Hahn, G.; Digel, M.; Katona, R.; Sampaio, K.L.; Messerle, M.; Hengel, H.; Koszinowski, U.; Brune, W.; Adler, B. Cloning and sequencing of a highly productive, endotheliotropic virus strain derived from human cytomegalovirus TB40/E. J. Gen. Virol. 2008, 89, 359–368. [Google Scholar] [CrossRef]
- Murphy, E.; Yu, D.; Grimwood, J.; Schmutz, J.; Dickson, M.; Jarvis, M.A.; Hahn, G.; Nelson, J.A.; Myers, R.M.; Shenk, T.E. Coding potential of laboratory and clinical strains of human cytomegalovirus. Proc. Natl. Acad. Sci. USA 2003, 100, 14976–14981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, C.; Gatherer, D.; Hilfrich, B.; Baluchova, K.; Dargan, D.J.; Thomson, M.; Griffiths, P.D.; Wilkinson, G.W.G.; Schulz, T.F.; Davison, A.J. Sequences of complete human cytomegalovirus genomes from infected cell cultures and clinical specimens. J. Gen. Virol. 2010, 91, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Renzette, N.; Bhattacharjee, B.; Jensen, J.D.; Gibson, L.; Kowalik, T.F. Extensive genome-wide variability of human cytomegalovirus in congenitally infected infants. PLoS Pathog. 2011, 7, e1001344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renzette, N.; Gibson, L.; Bhattacharjee, B.; Fisher, D.; Schleiss, M.R.; Jensen, J.D.; Kowalik, T.F. Rapid intrahost evolution of human cytomegalovirus is shaped by demography and positive selection. PLoS Genet. 2013, 9, e1003735. [Google Scholar] [CrossRef] [Green Version]
- Sijmons, S.; Thys, K.; Corthout, M.; Van Damme, E.; Van Loock, M.; Bollen, S.; Baguet, S.; Aerssens, J.; Van Ranst, M.; Maes, P. A method enabling high-throughput sequencing of human cytomegalovirus complete genomes from clinical isolates. PLoS ONE 2014, 9, e95501. [Google Scholar] [CrossRef] [Green Version]
- Renzette, N.; Pokalyuk, C.; Gibson, L.; Bhattacharjee, B.; Schleiss, M.R.; Hamprecht, K.; Yamamoto, A.Y.; Mussi-Pinhata, M.M.; Britt, W.J.; Jensen, J.D.; et al. Limits and patterns of cytomegalovirus genomic diversity in humans. Proc. Natl. Acad. Sci. USA 2015, 112, E4120–E4128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, N.M.; Wilkie, G.S.; Hage, E.; Camiolo, S.; Holton, M.; Hughes, J.; Maabar, M.; Vattipally, S.B.; Dhingra, A.; Gompels, U.A.; et al. Human cytomegalovirus genomes sequenced directly from clinical material: Variation, multiple-strain infection, recombination, and gene loss. J. Infect. Dis. 2019, 220, 781–791. [Google Scholar] [CrossRef] [Green Version]
- Cudini, J.; Roy, S.; Houldcroft, C.J.; Bryant, J.M.; Depledge, D.P.; Tutill, H.; Veys, P.; Williams, R.; Worth, A.J.J.; Tamuri, A.U.; et al. Human cytomegalovirus haplotype reconstruction reveals high diversity due to superinfection and evidence of within-host recombination. Proc. Natl. Acad. Sci. USA 2019, 116, 5693–5698. [Google Scholar] [CrossRef] [Green Version]
- Renzette, N.; Gibson, L.; Jensen, J.D.; Kowalik, T.F. Human cytomegalovirus intrahost evolution—A new avenue for understanding and controlling herpesvirus infections. Curr. Opin. Virol. 2014, 8, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Renzette, N.; Kowalik, T.F.; Jensen, J.D. On the relative roles of background selection and genetic hitchhiking in shaping human cytomegalovirus genetic diversity. Mol. Ecol. 2016, 25, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, A.Y.; Mussi-Pinhata, M.M.; Boppana, S.B.; Novak, Z.; Wagatsuma, V.M.; de Frizzo Oliveira, P.; Duarte, G.; Britt, W.J. Human cytomegalovirus reinfection is associated with intrauterine transmission in a highly cytomegalovirus-immune maternal population. Am. J. Obstet. Gynecol. 2010, 202, 297.e1–297.e8. [Google Scholar] [CrossRef]
- Pokalyuk, C.; Renzette, N.; Irwin, K.K.; Pfeifer, S.P.; Gibson, L.; Britt, W.J.; Yamamoto, A.Y.; Mussi-Pinhata, M.M.; Kowalik, T.F.; Jensen, J.D. Characterizing human cytomegalovirus reinfection in congenitally infected infants: An evolutionary perspective. Mol. Ecol. 2017, 26, 1980–1990. [Google Scholar] [CrossRef]
- Barbi, M.; Binda, S.; Caroppo, S.; Primache, V.; Didò, P.; Guidotti, P.; Corbetta, C.; Melotti, D. CMV gB genotypes and outcome of vertical transmission: Study on dried blood spots of congenitally infected babies. J. Clin. Virol. 2001, 21, 75–79. [Google Scholar] [CrossRef]
- Murphy, E.; Shenk, T.E. Human Cytomegalovirus Genome; Springer: Berlin/Heidelberg, Germany, 2008; Volume 325, ISBN 9783540773481. [Google Scholar]
- Britt, W.J.; Mach, M. Human cytomegalovirus glycoproteins. Intervirology 1996, 39, 401–412. [Google Scholar] [CrossRef]
- Isaacson, M.K.; Compton, T. Human cytomegalovirus glycoprotein B is required for virus entry and cell-to-cell spread but not for virion attachment, assembly, or egress. J. Virol. 2009, 83, 3891–3903. [Google Scholar] [CrossRef] [Green Version]
- Mach, M.; Kropff, B.; Dal Monte, P.; Britt, W. Complex Formation by Human Cytomegalovirus Glycoproteins M (gpUL100) and N (gpUL73). J. Virol. 2000, 74, 11881–11892. [Google Scholar] [CrossRef] [Green Version]
- Gretch, D.R.; Kari, B.; Rasmussen, L.; Gehrz, R.C.; Stinski, M.F. Identification and characterization of three distinct families of glycoprotein complexes in the envelopes of human cytomegalovirus. J. Virol. 1988, 62, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Shenk, T. Human cytomegalovirus virion protein complex required for epithelial and endothelial cell tropism. Proc. Natl. Acad. Sci. USA 2005, 102, 18153–18158. [Google Scholar] [CrossRef] [Green Version]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Chou, S.; Dennison, K.M. Analysis of interstrain variation in cytomegalovirus glycoprotein B sequences encoding neutralization-related epitopes. J. Infect. Dis. 1991, 163, 1229–1234. [Google Scholar] [CrossRef]
- Chou, S. Comparative analysis of sequence variation in gp116 and gp55 components of glycoprotein B of human cytomegalovirus. Virology 1992, 188, 388–390. [Google Scholar] [CrossRef]
- Shepp, D.H.; Match, M.E.; Lipson, S.M.; Pergolizzi, R.G. A fifth human cytomegalovirus glycoprotein B genotype. Res. Virol. 1998, 149, 109–114. [Google Scholar] [CrossRef]
- Pignatelli, S.; Dal Monte, P.; Landini, M.P. gpUL73 (gN) genomic variants of human cytomegalovirus isolates are clustered into four distinct genotypes. J. Gen. Virol. 2001, 82, 2777–2784. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, S.; Dal Monte, P.; Rossini, G.; Chou, S.; Gojobori, T.; Hanada, K.; Guo, J.J.; Rawlinson, W.; Britt, W.; Mach, M.; et al. Human cytomegalovirus glycoprotein N (gpUL73-gN) genomic variants: Identification of a novel subgroup, geographical distribution and evidence of positive selective pressure. J. Gen. Virol. 2003, 84, 647–655. [Google Scholar] [CrossRef]
- Paterson, D.A.; Dyer, A.P.; Milne, R.S.B.; Sevilla-Reyes, E.; Gompels, U.A. A role for human cytomegalovirus glycoprotein O (gO) in cell fusion and a new hypervariable locus. Virology 2002, 293, 281–294. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, L.; Geissler, A.; Cowan, C.; Chase, A.; Winters, M. The genes encoding the gCIII complex of human cytomegalovirus exist in highly diverse combinations in clinical isolates. J. Virol. 2002, 76, 10841–10848. [Google Scholar] [CrossRef] [Green Version]
- Stanton, R.; Westmoreland, D.; Fox, J.D.; Davison, A.J.; Wilkinson, G.W.G. Stability of Human cytomegalovirus genotypes in persistently infected renal transplant recipients. J. Med. Virol. 2005, 75, 42–46. [Google Scholar] [CrossRef]
- Mattick, C.; Dewin, D.; Polley, S.; Sevilla-Reyes, E.; Pignatelli, S.; Rawlinson, W.; Wilkinson, G.; Dal Monte, P.; Gompels, U.A. Linkage of human cytomegalovirus glycoprotein gO variant groups identified from worldwide clinical isolates with gN genotypes, implications for disease associations and evidence for N-terminal sites of positive selection. Virology 2004, 318, 582–597. [Google Scholar] [CrossRef] [Green Version]
- Görzer, I.; Guelly, C.; Trajanoski, S.; Puchhammer-Stöckl, E. Deep sequencing reveals highly complex dynamics of human cytomegalovirus genotypes in transplant patients over time. J. Virol. 2010, 84, 7195–7203. [Google Scholar] [CrossRef] [Green Version]
- Chou, S. Molecular epidemiology of envelope glycoprotein H of human cytomegalovirus. J. Infect. Dis. 1992, 166, 604–607. [Google Scholar] [CrossRef]
- Murthy, S.; Hayward, G.S.; Wheelan, S.; Forman, M.S.; Ahn, J.H.; Pass, R.F.; Arav-Boger, R. Detection of a single identical cytomegalovirus (CMV) strain in recently seroconverted young women. PLoS ONE 2011, 6, e15949. [Google Scholar] [CrossRef]
- Yan, H.; Koyano, S.; Inami, Y.; Yamamoto, Y.; Suzutani, T.; Mizuguchi, M.; Ushijima, H.; Kurane, I.; Inoue, N. Genetic linkage among human cytomegalovirus glycoprotein N (gN) and gO genes, with evidence for recombination from congenitally and post-natally infected Japanese infants. J. Gen. Virol. 2008, 89, 2275–2279. [Google Scholar] [CrossRef]
- Arav-Boger, R. Strain variation and disease severity in congenital cytomegalovirus infection: In search of a viral marker. Infect. Dis. Clin. N. Am. 2015, 29, 401–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, M.; Schoppel, K.; Amvrossiadis, N.; Mach, M. Strain-specific neutralization of human cytomegalovirus isolates by human sera. J. Virol. 1999, 73, 878–886. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Freed, D.C.; Wang, D.; Qiu, P.; Li, F.; Fu, T.-M.; Kauvar, L.M.; McVoy, M.A. Impact of antibodies and strain polymorphisms on cytomegalovirus entry and spread in fibroblasts and epithelial cells. J. Virol. 2017, 91, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Beninga, J.; Kalbacher, H.; Mach, M. Analysis of T helper cell response to glycoprotein H (gpUL75) of human cytomegalovirus: Evidence for strain-specific T cell determinants. J. Infect. Dis. 1996, 173, 1051–1061. [Google Scholar] [CrossRef] [Green Version]
- Burke, H.G.; Heldwein, E.E. Crystal structure of the human cytomegalovirus glycoprotein B. PLoS Pathog. 2015, 11, e1005227. [Google Scholar] [CrossRef] [Green Version]
- Chandramouli, S.; Ciferri, C.; Nikitin, P.A.; Caló, S.; Gerrein, R.; Balabanis, K.; Monroe, J.; Hebner, C.; Lilja, A.E.; Settembre, E.C.; et al. Structure of HCMV glycoprotein B in the postfusion conformation bound to a neutralizing human antibody. Nat. Commun. 2015, 6, 8176. [Google Scholar] [CrossRef] [Green Version]
- Bagdonaite, I.; Nordén, R.; Joshi, H.J.; King, S.L.; Vakhrushev, S.Y.; Olofsson, S.; Wandall, H.H. Global mapping of o-glycosylation of varicella zoster virus, human cytomegalovirus, and Epstein-Barr virus. J. Biol. Chem. 2016, 291, 12014–12028. [Google Scholar] [CrossRef] [Green Version]
- Spaete, R.R.; Thayer, R.M.; Probert, W.S.; Masiarz, F.R.; Chamberlain, S.H.; Rasmussen, L.; Merigan, T.C.; Pachl, C. Human cytomegalovirus strain Towne glycoprotein B is processed by proteolytic cleavage. Virology 1988, 167, 207–225. [Google Scholar] [CrossRef]
- Haberland, M.; Meyer-König, U.; Hufert, F.T. Variation within the glycoprotein B gene of human cytomegalovirus is due to homologous recombination. J. Gen. Virol. 1999, 80, 1495–1500. [Google Scholar] [CrossRef]
- Fan, J.; Zhang, X.; Chen, X.M.; Gao, H.N.; Yang, M.F.; Zhao, H.; Hu, J.H.; Ma, W.H. Monitoring of human cytomegalovirus glycoprotein B genotypes using real-time quantitative PCR in immunocompromised Chinese patients. J. Virol. Methods 2009, 160, 74–77. [Google Scholar] [CrossRef]
- Grosjean, J.; Hantz, S.; Cotin, S.; Baclet, M.C.; Mengelle, C.; Trapes, L.; Virey, B.; Undreiner, F.; Brosset, P.; Pasquier, C.; et al. Direct genotyping of cytomegalovirus envelope glycoproteins from toddler’s saliva samples. J. Clin. Virol. 2009, 46, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, Y.P.; Gao, H.N.; Yang, M.F.; Zhao, H.; Hu, J.H.; Chen, X.M.; Ma, W.H.; Fan, J. Quantification of cytomegalovirus glycoprotein Bn DNA in hematopoietic stem cell transplant recipients by real-time PCR. PLoS ONE 2012, 7, e51224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer-König, U.; Haberland, M.; Von Laer, D.; Haller, O.; Hufert, F.T. Intragenic variability of human cytomegalovirus glycoprotein B in clinical strains. J. Infect. Dis. 1998, 177, 1162–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, L.; Hong, C.; Zipeto, D.; Morris, S.; Sherman, D.; Chou, S.; Miner, R.; Drew, W.L.; Wolitz, R.; Dowling, A.; et al. Cytomegalovirus gB genotype distribution differs in human immunodeficiency virus-infected patients and immunocompromised allograft recipients. J. Infect. Dis. 1997, 175, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Peek, R.; Verbraak, F.; Bruinenberg, M.; Van Der Lelij, A.; Van Den Horn, G.; Kijlstra, A. Cytomegalovirus glycoprotein B genotyping in ocular fluids and blood of AIDS patients with cytomegalovirus retinitis. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1183–1187. [Google Scholar]
- Wada, K.; Mizuno, S.; Kato, K.; Kamiya, T.; Ozawa, K. Cytomegalovirus glycoprotein b sequence variation among japanese bone marrow transplant recipients. Intervirology 1997, 40, 215–219. [Google Scholar] [CrossRef]
- Pacsa, A.S.; Essa, S.; Voevodin, A.; El-Shazly, A.; Kazak, H.; Nampoory, M.R.N.; Johny, K.V.; Said, T.; Al-Nakib, W. Correlation between CMV genotypes, multiple infections with herpesviruses (HHV-6, 7) and development of CMV disease in kidney recipients in Kuwait. FEMS Immunol. Med. Microbiol. 2003, 35, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Numazaki, K.; Tsutsumi, H. Human cytomegalovirus genetic variability in strains isolated from Japanese children during 1983–2003. J. Med. Virol. 2005, 76, 356–360. [Google Scholar] [CrossRef]
- Yu, Z.S.; Zou, C.C.; Zheng, J.Y.; Zhao, Z.Y. Cytomegalovirus gB genotype and clinical features in Chinese infants with congenital infections. Intervirology 2006, 49, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Wang, X.; Li, S. Human cytomegalovirus glycoprotein B genotype correlates with different symptoms of infected infants. Intervirology 2007, 50, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Koyano, S.; Inami, Y.; Yamamoto, Y.; Suzutani, T.; Mizuguchi, M.; Ushijima, H.; Kurane, I.; Inoue, N. Genetic variations in the gB, UL144 and UL149 genes of human cytomegalovirus strains collected from congenitally and postnatally infected Japanese children. Arch. Virol. 2008, 153, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.L.; Cai, T.; Jin, H.M. Human cytomegalovirus glycoprotein genotypes in the genital tract tissue of tubal pregnancy patients. J. Int. Med. Res. 2009, 37, 385–391. [Google Scholar] [CrossRef]
- Mewara, A.; Mishra, B.; Ratho, R.K.; Kumar, P. Cytomegalovirus glycoprotein b gene polymorphism and its association with clinical presentations in infants. Southeast Asian J. Trop. Med. Public Health 2009, 40, 759–764. [Google Scholar]
- Shen, Z.; Shang, S.Q.; Zou, C.C.; Zheng, J.Y.; Yu, Z.S. The detection and clinical features of human cytomegalovirus infection in infants. Fetal Pediatr. Pathol. 2010, 29, 393–400. [Google Scholar] [CrossRef]
- Gandhoke, I.; Hussain, S.A.; Pasha, S.T.; Chauhan, L.S.; Khare, S. Glycoprotein B genotyping in congenital/perinatal cytomegalovirus infection in symptomatic infants. Indian Pediatr. 2013, 50, 663–667. [Google Scholar] [CrossRef]
- Paca-Uccaralertkun, S.; Hiatt, R.; Leecharoen, R.; Tan-Ariya, P.; Mungthin, M.; Pongphong, S. Human cytomegalovirus gB1 genotypes among children who live at the Phayathai Babies’ home in Nonthaburi, Thailand. Southeast Asian J. Trop. Med. Public Health 2013, 44, 636–640. [Google Scholar]
- Ding, Z.Y.; Xu, F.; Chen, D.Z.; Meng, X.N.; Xu, T.S.; Lu, M.D.; Zhuge, H.X. A multifactorial analysis of the pregnancy outcomes in cytomegalovirus-infected women. Gynecol. Obstet. Investig. 2015, 80, 106–112. [Google Scholar] [CrossRef]
- Zhou, L.; Fan, J.; Zheng, S.S.; Ma, W.H. Genetic variation within the glycoprotein B and H genes of human cytomegalovirus in solid organ transplant recipients. Transpl. Infect. Dis. 2007, 9, 73–77. [Google Scholar] [CrossRef]
- Mujtaba, G.; Khurshid, A.; Sharif, S.; Alam, M.M.; Aamir, U.B.; Shaukat, S.; Angez, M.; Rana, M.S.; Umair, M.; Shah, A.A.; et al. Distribution of cytomegalovirus genotypes among neonates born to infected mothers in Islamabad, Pakistan. PLoS ONE 2016, 11, e156049. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Y.; Zheng, T.L.; Zhou, T.; Hu, P.W.; Huang, M.J.; Xu, X.; Pei, X.F. Human cytomegalovirus prevalence and distribution of glycoprotein B, O genotypes among hospitalized children with respiratory infections in West China, 2009–2014. Trop. Med. Int. Health 2016, 21, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Das, D.; Ansari, S.; Chatterjee, R.P.; Mishra, L.; Basu, B.; Ghosh, S.K.; Bhattacharyay, M.; Chakraborty, N. Genotypes of glycoprotein B gene among the Indian symptomatic neonates with congenital CMV infection. BMC Pediatr. 2019, 19, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Alwan, S.N.; Shamran, H.A.; Ghaib, A.H.; Kadhim, H.S.; Al-Mayah, Q.S.; AL-Saffar, A.J.; Bayati, A.H.; Arif, H.S.; Fu, J.; Wickes, B.L. Genotyping of cytomegalovirus from symptomatic infected neonates in Iraq. Am. J. Trop. Med. Hyg. 2019, 100, 957–963. [Google Scholar] [CrossRef]
- Min, X.; Wang, L.; Cui, A.; Zhang, C.; Wang, D.; Liu, Y.; Li, Z.; Xu, W. The nucleic acid positive rate and genotype distribution of human cytomegalovirus in human milk banks in China. Arch. Virol. 2020, 165, 1099–1107. [Google Scholar] [CrossRef]
- Dong, N.; Cao, L.; Su, L.; Lu, L.; Dong, Z.; Xu, M.; Xu, J. Human cytomegalovirus envelope glycoprotein B, H, and N polymorphisms among infants of Shanghai area in China. J. Med. Virol. 2020, 92, 3674–3681. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lo, C.Y.; Lo, S.K.F.; Siau, H.; Peiris, J.S.M.; Wong, S.S.Y.; Luk, W.K.; Chan, T.M.; Lim, W.W.; Yuen, K.Y. Distinct genotypic distributions of cytomegalovirus (CMV) envelope glycoprotein in bone marrow and renal transplant recipients with CMV disease. Clin. Diagn. Lab. Immunol. 1997, 4, 515–518. [Google Scholar] [CrossRef] [Green Version]
- Madhaven, H.N.; Priya, K. Polymerase chain reaction based restriction fragment length polymorphism for the genotyping of cytomegalovirus (CMV) from patients with CMV disease in Chennai. Indian J. Med. Res. 2002, 130, 556. [Google Scholar]
- Wu, Y.M.; Yan, J.; Ojcius, D.M.; Chen, L.L.; Gu, Z.Y.; Pan, J.P. Correlation between infections with different genotypes of human cytomegalovirus and Epstein-Barr virus in subgingival samples and periodontal status of patients. J. Clin. Microbiol. 2007, 45, 3665–3670. [Google Scholar] [CrossRef] [Green Version]
- Bhattarakosol, P.; Chantaraarphonkun, S. Prevalence of human cytomegalovirus (HCMV) gB genotypes in Thai patients. Southeast Asian J. Trop. Med. Public Health 2007, 38, 835–840. [Google Scholar] [PubMed]
- Pang, X.; Humar, A.; Preiksaitis, J.K. Concurrent genotyping and quantitation of cytomegalovirus gB genotypes in solid-organ-transplant recipients by use of a real-time PCR assay. J. Clin. Microbiol. 2008, 46, 4004–4010. [Google Scholar] [CrossRef] [Green Version]
- Chantaraarphonkun, S.; Bhattarakosol, P. Intra- and Intergenotypic variations among human cytomegalovirus gB genotypes. Intervirology 2007, 10330, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.S.; Tang, L.F.; Zou, C.C.; Zheng, J.Y.; Zhao, Z.Y. Cytomegalovirus-associated idiopathic thrombocytopenic purpura in Chinese children. Scand. J. Infect. Dis. 2008, 40, 922–927. [Google Scholar] [CrossRef]
- Wu, K.G.; Hung, M.C.; Chang, Y.T.; Chen, C.J.; Yang, S.P.; Liu, C.Y.; Ho, D.M.T.; Chan, Y.J. Occurrence of human cytomegalovirus glycoprotein B genotypes in immunocompetent and immunosuppressed Taiwanese patients. Intervirology 2011, 54, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Li, J.W.; Liu, Y.M.; Zhong, H.; Wang, L.M.; Deng, F.M.; Qu, Y.Y.; Hui, J.; Cheng, J.; Tang, B.; et al. Human cytomegalovirus infection is associated with essential hypertension in Kazakh and Han Chinese populations. Med. Sci. Monit. 2014, 20, 2508–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taherkhani, R.; Farshadpour, F.; Makvandi, M.; Hamidifard, M.; Esmailizadeh, M.; Ahmadi, B.; Heidari, H. Determination of cytomegalovirus prevalence and glycoprotein B genotypes among ulcerative colitis patients in Ahvaz, Iran. Jundishapur J. Microbiol. 2015, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, N.; Suzuki, T.; Inoue, T.; Kobayashi, T.; Ohashi, Y. Polymorphisms in cytomegalovirus genotype in immunocompetent patients with corneal endotheliitis or iridocyclitis. J. Med. Virol. 2015, 87, 1441–1445. [Google Scholar] [CrossRef]
- Eran Daǧlar, D.; Öngüt, G.; Çolak, D.; Özkul, A.; Mutlu, D.; Zeytenoǧlu, A.; Midilli, K.; Gökahmetoǧlu, S.; Günseren, F.; Ögünç, D.; et al. Determination of cytomegalovirus glycoprotein B genotypes in different geographical regions and different patient groups in Turkey. Mikrobiyol. Bul. 2016, 50, 53–62. [Google Scholar] [CrossRef]
- Hu, H.; Cheng, Y.; Peng, Q.; Chen, K. Clinical features, treatment courses, and distribution of cytomegalovirus genotypes among thrombocytopenia patients aged younger than 12 months. Am. J. Perinatol. 2020, 1. [Google Scholar] [CrossRef]
- Nahar, S.; Hokama, A.; Iraha, A.; Ohira, T.; Kinjo, T.; Hirata, T.; Kinjo, T.; Parrott, G.L.; Fujita, J. Distribution of cytomegalovirus genotypes among ulcerative colitis patients in Okinawa, Japan. Intest. Res. 2018, 16, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, H.T.; El-Shinawi, M.; Nouh, M.A.; Bashtar, A.R.; Elsayed, E.T.; Schneider, R.J.; Mohamed, M.M. Inflammatory breast cancer: High incidence of detection of mixed human cytomegalovirus genotypes associated with disease pathogenesis. Front. Oncol. 2014, 4, 246. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wang, Y.; Xu, Y.; Wu, D.; Sun, A.; Zhu, Z.; Han, Y.; Qiu, H.; Tang, X.; Fu, Z.; et al. Cytomegalovirus glycoprotein B genotype in hematopoietic stem cell transplant patients from China. Biol. Blood Marrow Transplant. 2010, 16, 647–652. [Google Scholar] [CrossRef] [Green Version]
- Madi, N.; Al-Nakib, W.; Pacsa, A.; Saeed, T. Cytomegalovirus genotypes gB1 and gH1 Are the most predominant genotypes among renal transplant recipients in Kuwait. Transplant. Proc. 2011, 43, 1634–1637. [Google Scholar] [CrossRef]
- Xia, C.S.; Zhao, X.T.; Sun, Y.Y.; Zhang, Z. Human cytomegalovirus glycoprotein B genotypes in Chinese hematopoietic stem cell transplant recipients. Intervirology 2012, 55, 342–348. [Google Scholar] [CrossRef]
- Soleimani, A.R.; Jafari, M.; Piroozmand, A.; Nikoueinejad, H.; Akbari, H.; Einollahi, B. The Incidence of Cytomegalovirus Glycoprotein B Genotypes in Kidney Transplant Recipients in Iran. Int. J. Organ Transplant. Med. 2018, 9, 173–177. [Google Scholar] [PubMed]
- Jiang, X.J.; Zhang, J.; Xiong, Y.; Jahn, G.; Xiong, H.R.; Yang, Z.Q.; Liu, Y.Y. Human cytomegalovirus glycoprotein polymorphisms and increasing viral load in AIDS patients. PLoS ONE 2017, 12, e0176160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terabe, K.; Sugiyama, K.; Goto, K.; Mizutani, F.; Wada, Y.; Yokoyama, T.; Ando, Y. Relationship between human cytomegalovirus glycoprotein B genotype and serum alanine aminotransferase elevation in infants. Tohoku J. Exp. Med. 2004, 203, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Torok-Storb, B.; Boeckh, M.; Hoy, C.; Leisenring, W.; Myerson, D.; Gooley, T. Association of specific cytomegalovirus genotypes with death from myelosuppression after marrow transplantation. Blood 1997, 90, 2097–2102. [Google Scholar] [CrossRef] [PubMed]
- White, J.L.; Patel, E.U.; Abraham, A.G.; Grabowski, M.K.; Arav-Boger, R.; Avery, R.K.; Quinn, T.C.; Tobian, A.A.R. Prevalence, magnitude, and genotype distribution of urinary cytomegalovirus (CMV) shedding among CMV-seropositive children and adolescents in the United States. Open Forum Infect. Dis. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Sarcinella, L.; Mazzulli, T.; Willey, B.; Humar, A. Cytomegalovirus glycoprotein B genotype does not correlate with outcomes in liver transplant patients. J. Clin. Virol. 2002, 24, 99–105. [Google Scholar] [CrossRef]
- Humar, A.; Kumar, D.; Gilbert, C.; Boivin, G. Cytomegalovirus (CMV) glycoprotein B genotypes and response to antiviral therapy, in solid-organ-transplant recipients with CMV disease. J. Infect. Dis. 2003, 188, 581–584. [Google Scholar] [CrossRef]
- Manuel, O.; Pang, X.L.; Humar, A.; Kumar, D.; Doucette, K.; Preiksaitis, J.K. An assessment of donor-to-recipient transmission patterns of human cytomegalovirus by analysis of viral genomic variants. J. Infect. Dis. 2009, 199, 1621–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuel, O.; Åsberg, A.; Pang, X.; Rollag, H.; Emery, V.C.; Preiksaitis, J.K.; Kumar, D.; Pescovitz, M.D.; Bignamini, A.A.; Hartmann, A.; et al. Impact of genetic polymorphisms in cytomegalovirus glycoprotein b on outcomes in solid-organ transplant recipients with cytomegalovirus disease. Clin. Infect. Dis. 2009, 49, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.; Handfield, J.; Toma, E.; Lalonde, R.; Bergeron, M.G.; Boivin, G. Human cytomegalovirus glycoprotein B genotypes in blood of AIDS patients: Lack of association with either the viral DNA load in leukocytes or presence of retinitis. J. Med. Virol. 1999, 59, 98–103. [Google Scholar] [CrossRef]
- Drew, W.L.; Chou, S.; Miner, R.C.; Mohr, B.A.; Busch, M.P.; van der Horst, C.M.; Asmuth, D.M.; Kalish, L.A. Cytomegalovirus glycoprotein B groups in human immunodeficiency virus-infected patients with incident retinitis. J. Infect. Dis. 2002, 186, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Bale, J.F.; Murph, J.R.; Demmler, G.J.; Dawson, J.; Miller, J.E.; Petheram, S.J. Intrauterine cytomegalovirus infection and glycoprotein B genotypes. J. Infect. Dis. 2000, 182, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Pati, S.K.; Pinninti, S.; Novak, Z.; Chowdhury, N.; Patro, R.K.; Fowler, K.; Ross, S.; Boppana, S. Genotypic diversity and mixed infection in newborn disease and hearing loss in congenital cytomegalovirus infection. Pediatr. Infect. Dis. J. 2013, 32, 1050–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aquino, V.H.; Figueiredo, L.T.M. High prevalence of renal transplant recipients infected with more than one cytomegalovirus glycoprotein B genotype. J. Med. Virol. 2000, 61, 138–142. [Google Scholar] [CrossRef]
- Carraro, E.; Granato, C.F.H. Single human cytomegalovirus gB genotype shed in multiple sites at the time of diagnosis in renal transplant recipients. J. Med. Virol. 2003, 70, 240–243. [Google Scholar] [CrossRef]
- Correa, C.; Kourí, V.; Pérez, L.; Soto, Y.; Limia, C. Diagnosis, gB genotype distribution and viral load of symptomatic congenitally infected CMV patients in Cuba. J. Perinatol. 2016, 36, 837–842. [Google Scholar] [CrossRef]
- Ahumada-Ruiz, S.; Taylor-Castillo, L.; Visoná, K.; Luftig, R.B.; Herrero-Uribe, L. Determination of human cytomegalovirus genetic diversity in different patient populations in Costa Rica. Rev. Inst. Med. Trop. Sao Paulo 2004, 46, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourí, V.; González, E.; Martínez, P.; Capó, V.; González, R.; Pérez, L.; Viera, J.; Cardellá, L.; Hengge, U.R. Distinct genotypic distribution of cytomegalovirus (CMV) envelope glycoprotein B (gB) in a Cuban cohort of patients with different CMV diseases. Scand. J. Infect. Dis. 2007, 39, 1038–1044. [Google Scholar] [CrossRef]
- Slavov, S.N.; Kashima, S.; Wagatsuma, V.M.D.; Silva-Pinto, A.C.; Martinez, E.Z.; Favarin, M.D.C.; Covas, D.T. Glycoprotein B genotyping of human cytomegalovirus strains isolated from Brazilian patients with sickle cell disease and beta-thalassemia major. Viral Immunol. 2015, 28, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, E.; Ozaki, K.S.; Tomiyama, H.; Câmara, N.O.S.; Granato, C.F.H. Clinical correlations of human cytomegalovirus strains and viral load in kidney transplant recipients. Int. Immunopharmacol. 2009, 9, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Dieamant, D.D.C.; Bonon, S.H.A.; Prates, L.C.; Belangelo, V.M.S.; Pontes, E.R.; Costa, S.C.B. Active human cytomegalovirus infection and glycoprotein B genotypes in Brazilian pediatric renal or hematopoietic stem cell transplantation patients. Braz. J. Microbiol. 2009, 53, 1689–1699. [Google Scholar] [CrossRef] [Green Version]
- Correia-Silva, J.F.; Resende, R.G.; Arão, T.C.; Abreu, M.; Teixeira, M.M.; Bittencourt, H.; Silva, T.A.; Gomez, R.S. HCMV gB genotype and its association with cytokine levels in hematopoietic stem cell transplantation. Oral Dis. 2011, 17, 530–537. [Google Scholar] [CrossRef] [PubMed]
- González-Ramírez, J.; Uribe-Gutiérrez, G.; Jiménez-Hernandez, E.; Velázquez-Guadarrama, N.; Bello-González, A.; Vazquez-Meraz, E.; Arellano-Galindo, J. Cytomegalovirus gB genotype distribution in Mexican children undergoing allogeneic bone marrow transplantation. Intervirology 2012, 55, 318–320. [Google Scholar] [CrossRef]
- Vilas Boas, L.S.; De Souza, V.A.U.F.; De Oliveira, A.C.P.; Rodriguez Viso, A.T.; Nascimento Filho, A.M.; Nascimento, M.C.; Pannuti, C.S. Cytomegalovirus glycoprotein B genotypes and central nervous system disease in AIDS patients. J. Med. Virol. 2003, 71, 404–407. [Google Scholar] [CrossRef]
- Cunha, A.A.; Aquino, V.H.; Mariguela, V.; Nogueira, M.L.; Figueiredo, L.T.M. Evaluation of glycoprotein B genotypes and load of CMV infecting blood leukocytes on prognosis of AIDS patients. Rev. Inst. Med. Trop. Sao Paulo 2011, 53, 83–88. [Google Scholar] [CrossRef]
- Arellano-Galindo, J.; Villanueva-García, D.; Cruz-Ramírez, J.L.; Yalaupari-Mejía, J.P.; Uribe-Gutiérrez, G.; Velazquez-Guadarrama, N.; Nava-Frías, M.; Muñoz-Hernández, O.; Mejía-Aranguré, J.M. Detection and gB genotyping of CMV in Mexican preterm infants in the context of maternal seropositivity. J. Infect. Dev. Ctries. 2014, 8, 758–767. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Sanchez, H.M.; Alvarado-Hernandez, D.L.; Guerra-Palomares, S.; Garcia-Sepulveda, C.A.; Noyola, D.E. Cytomegalovirus glycoprotein B genotypes in Mexican children and women. Intervirology 2015, 58, 115–121. [Google Scholar] [CrossRef]
- Barrado, L.; Prieto, C.; Hernando, S.; Folgueira, L. Detection of glycoproteins B and H genotypes to predict the development of Cytomegalovirus disease in solid organ transplant recipients. J. Clin. Virol. 2018, 109, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Ciotti, M.; Cella, E.; Rittà, M.; Ciccozzi, M.; Cavallo, R.; Perno, C.F.; Costa, C. Cytomegalovirus glycoprotein B genotype distribution in Italian transplant patients. Intervirology 2018, 60, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Janković, M.; Ćupić, M.; Knežević, A.; Vujić, D.; Soldatović, I.; Zečević, Ž.; Gobeljić, B.; Jovanović, T. Cytomegalovirus glycoprotein B and N genotypes in pediatric recipients of the hematopoietic stem cell transplant. Virology 2020, 548, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Fidouh-Houhou, N.; Duval, X.; Bissuel, F.; Bourbonneux, V.; Flandre, P.; Ecobichon, J.L.; Jordan, M.C.; Vildé, J.L.; Brun-Vézinet, F.; Leport, C. Salivary cytomegalovirus (CMV) shedding, glycoprotein b genotype distribution, and CMV disease in human immunodeficiency virus-seropositive patients. Clin. Infect. Dis. 2001, 33, 1406–1411. [Google Scholar] [CrossRef] [PubMed]
- Tarragó, D.; Quereda, C.; Tenorio, A. Different cytomegalovirus glycoprotein B genotype distribution in serum and cerebrospinal fluid specimens determined by a novel multiplex nested PCR. J. Clin. Microbiol. 2003, 41, 2872–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, V.J.; Wolffs, P.F.; Van Loo, I.H.; Bruggeman, C.A.; Verbon, A. CMV DNA levels and CMV gB subtypes in ART-naive HAART-treated patients: A 2-year follow-up study in the Netherlands. Aids 2009, 23, 1425–1429. [Google Scholar] [CrossRef]
- Vogel, J.U.; Otte, J.; Koch, F.; Gümbel, H.; Doerr, H.W.; Cinatl, J. Role of human cytomegalovirus genotype polymorphisms in AIDS patients with cytomegalovirus retinitis. Med. Microbiol. Immunol. 2013, 202, 37–47. [Google Scholar] [CrossRef]
- Lukacsi, A.; Tarodi, B.; Endreffy, E.; Babinszki, A.; Pal, A.; Pusztai, R. Human cytomegalovirus gB genotype 1 is dominant in congenital infections in South Hungary. J. Med. Virol. 2001, 65, 537–542. [Google Scholar] [CrossRef]
- Picone, O.; Costa, J.M.; Leruez-Ville, M.; Ernault, P.; Olivi, M.; Ville, Y. Cytomegalovirus (CMV) glycoprotein B genotype and CMV DNA load in the amniotic fluid of infected fetuses. Prenat. Diagn. 2004, 24, 1001–1006. [Google Scholar] [CrossRef]
- Grosjean, J.; Trapes, L.; Hantz, S.; Mengelle, C.; Virey, B.; Undreiner, F.; Messager, V.; Denis, F.; Marin, B.; Alain, S. Human cytomegalovirus quantification in toddlers saliva from day care centers and emergency unit: A feasibility study. J. Clin. Virol. 2014, 61, 371–377. [Google Scholar] [CrossRef]
- Zawilińska, B.; Szostek, S.; Kopeć, J.; Koprynia, M.; Kosz-Vnenchak, M. UL55 genotype diversity of cytomegalovirus strains isolated from newborns and infants hospitalized in southern Poland. Przegląd Epidemiol. 2011, 65, 409–413. [Google Scholar]
- Paradowska, E.; Studzińska, M.; Nowakowska, D.; Wilczyński, J.; Rycel, M.; Suski, P.; Gaj, Z.; Kaczmarek, B.; Zbróg, Z.; Leśnikowski, Z.J. Distribution of UL144, US28 and UL55 genotypes in Polish newborns with congenital cytomegalovirus infections. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Nijman, J.; Mandemaker, F.S.; Verboon-Maciolek, M.A.; Aitken, S.C.; Van Loon, A.M.; De Vries, L.S.; Schuurman, R. Genotype distribution, viral load and clinical characteristics of infants with postnatal or congenital cytomegalovirus infection. PLoS ONE 2014, 9, e108018. [Google Scholar] [CrossRef] [Green Version]
- Paradowska, E.; Studzińska, M.; Suski, P.; Kasztelewicz, B.; Wisniewska-Ligier, M.; Zawilinska, B.; Gaj, Z.; Nowakowska, D. Human cytomegalovirus UL55, UL144, and US28 genotype distribution in infants infected congenitally or postnatally. J. Med. Virol. 2015, 87, 1737–1748. [Google Scholar] [CrossRef] [PubMed]
- Görzer, I.; Trajanoski, S.; Popow-Kraupp, T.; Puchhammer-Stöckl, E. Analysis of human cytomegalovirus strain populations in urine samples of newborns by ultra deep sequencing. J. Clin. Virol. 2015, 73, 101–104. [Google Scholar] [CrossRef]
- Arcangeletti, M.C.; Simone, R.V.; Rodighiero, I.; De Conto, F.; Medici, M.C.; Martorana, D.; Chezzi, C.; Calderaro, A. Combined genetic variants of human cytomegalovirus envelope glycoproteins as congenital infection markers. Virol. J. 2015, 12, 1–11. [Google Scholar] [CrossRef]
- Brañas, P.; Blázquez-Gamero, D.; Galindo, A.; Prieto, C.; Olabafrieta, I.; Cuadrado, I.; Folgueria, L. Cytomegalovirus genotype distribution among congenitally and postnatally infected patients: Association of particular glycoprotein (g)B and gN types with symptomatic disease. Open Forum Infect. Dis. 2015, 2, 2633851. [Google Scholar] [CrossRef] [Green Version]
- Kadambari, S.; Atkinson, C.; Luck, S.; Macartney, M.; Conibear, T.; Harrison, I.; Booth, C.; Sharland, M.; Griffiths, P.D. Characterising variation in five fenetic loci of cytomegalovirus during treatment for congenital infection. J. Med. Virol. 2017, 89, 502–507. [Google Scholar] [CrossRef]
- Puhakka, L.; Pati, S.; Lappalainen, M.; Lönnqvist, T.; Niemensivu, R.; Lindahl, P.; Nieminen, T.; Seuri, R.; Nupponen, I.; Boppana, S.; et al. Viral shedding, and distribution of cytomegalovirus glycoprotein H (UL75), glycoprotein B (UL55), and glycoprotein N (UL73) genotypes in congenital cytomegalovirus infection. J. Clin. Virol. 2020, 125, 104287. [Google Scholar] [CrossRef]
- Arista, S.; De Grazia, S.; Giammanco, G.M.; Di Carlo, P.; Iannitto, E. Human cytomegalovirus glycoprotein B genotypes in immunocompetent, immunocompromised, and congenitally infected Italian populations. Arch. Virol. 2003, 148, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Coaquette, A.; Bourgeois, A.; Dirand, C.; Varin, A.; Chen, W.; Herbein, G. Mixed cytomegalovirus glycoprotein B genotypes in immunocompromised patients. Clin. Infect. Dis. 2004, 39, 155–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Görzer, I.; Kerschner, H.; Redlberger-Fritz, M.; Puchhammer-Stöckl, E. Human cytomegalovirus (HCMV) genotype populations in immunocompetent individuals during primary HCMV infection. J. Clin. Virol. 2010, 48, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Bergallo, M.; Costa, C.; Gambarino, S.; Tornicelli, A.; Astegiano, S.; Terlizzi, M.E.; Solidoro, P.; Cavallo, R. Human cytomegalovirus glycoprotein B genotyping from bronchoalveolar lavage specimens. Can. J. Microbiol. 2011, 57, 273–277. [Google Scholar] [CrossRef]
- Jakovljevic, A.; Andric, M.; Knezevic, A.; Soldatovic, I.; Nikolic, N.; Karalic, D.; Milasin, J. Human cytomegalovirus and Epstein-Barr virus genotypes in apical periodontitis lesions. J. Endod. 2015, 41, 1847–1851. [Google Scholar] [CrossRef] [PubMed]
- Fries, B.C.; Chon, S.; Boeckh, M.; Torok-Storb, B. Frequency distribution of cytomegalovirus envelope glycoprotein genotypes in bone marrow transplant recipients. J. Infect. Dis. 1994, 169, 769–774. [Google Scholar] [CrossRef]
- Vogelberg, C.; Meyer-König, U.; Hufert, F.T.; Kirste, G.; Von Laer, D. Human cytomegalovirus glycoprotein B genotypes in renal transplant recipients. J. Med. Virol. 1996, 50, 31–34. [Google Scholar] [CrossRef]
- Hebart, H.; Greif, M.; Krause, H.; Kanz, L.; Jahn, G.; Müller, C.A.; Einsele, H. Interstrain variation of immediate early DNA sequences and glycoprotein B genotypes in cytomegalovirus clinical isolates. Med. Microbiol. Immunol. 1997, 186, 135–138. [Google Scholar] [CrossRef]
- Meyer-König, U.; Vogelberg, C.; Bongarts, A.; Kampa, D.; Delbrück, R.; Wolff-Vorbeck, G.; Kirste, G.; Haberland, M.; Hufert, F.T.; Von Laer, D. Glycoprotein B genotype correlates with cell tropism in vivo of human cytomegalovirus infection. J. Med. Virol. 1998, 55, 75–81. [Google Scholar] [CrossRef]
- De Vries, J.J.C.; Wessels, E.; Korver, A.M.H.; Van Der Eijk, A.A.; Rusman, L.G.; Kroes, A.C.M.; Vossen, A.C.T.M. Rapid genotyping of cytomegalovirus in dried blood spots by multiplex real-time PCR assays targeting the envelope glycoprotein gB and gH genes. J. Clin. Microbiol. 2012, 50, 232–237. [Google Scholar] [CrossRef] [Green Version]
- Zawilinska, B.; Szostek, S.; Kopec, J.; Piatkowska-Jakubas, B.; Kosz-Vnenchak, M. Multiplex real-time PCR to identify a possible reinfection with different strains of human cytomegalovirus in allogeneic hematopoietic stem cell transplant recipients. Acta Biochim. Pol. 2016, 63, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Trincado, D.E.; Scott, G.M.; White, P.A.; Hunt, C.; Rasmussen, L.; Rawlinson, W.D. Human cytomegalovirus strains associated with congenital and perinatal infections. J. Med. Virol. 2000, 61, 481–487. [Google Scholar] [CrossRef]
- Cranage, M.P.; Kouzarides, T.; Bankier, A.T.; Satchwell, S.; Weston, K.; Tomlinson, P.; Barrell, B.; Hart, H.; Bell, S.E.; Minson, A.C. Identification of the human cytomegalovirus glycoprotein B gene and induction of neutralizing antibodies via its expression in recombinant vaccinia virus. EMBO J. 1986, 5, 3057–3063. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.S.; Huffman, T.; Jenks, J.A.; De La Rosa, E.C.; Xie, G.; Vandergrift, N.; Pass, R.F.; Pollara, J.; Permar, S.R. HCMV glycoprotein B subunit vaccine efficacy mediated by nonneutralizing antibody effector functions. Proc. Natl. Acad. Sci. USA 2018, 115, 6267–6272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bootz, A.; Karbach, A.; Spindler, J.; Kropff, B.; Reuter, N.; Sticht, H.; Winkler, T.H.; Britt, W.J.; Mach, M. Protective capacity of neutralizing and non-neutralizing antibodies against glycoprotein B of cytomegalovirus. PLoS Pathog. 2017, 13, e1006601. [Google Scholar] [CrossRef] [PubMed]
- Baraniak, I.; Gomes, A.C.; Sodi, I.; Langstone, T.; Rothwell, E.; Atkinson, C.; Pichon, S.; Piras-Douce, F.; Griffiths, P.D.; Reeves, M.B. Seronegative patients vaccinated with cytomegalovirus gB-MF59 vaccine have evidence of neutralising antibody responses against gB early post-transplantation. EBioMedicine 2019, 50, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, D.I.; Munoz, F.M.; Callahan, S.T.; Rupp, R.; Wootton, S.H.; Edwards, K.M.; Turley, C.B.; Stanberry, L.R.; Patel, S.M.; Mcneal, M.M.; et al. Safety and efficacy of a cytomegalovirus glycoprotein B (gB) vaccine in adolescent girls: A randomized clinical trial. Vaccine 2016, 34, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Pass, R.F. Detection of cytomegalovirus infection during clinical trials of glycoprotein B vaccine. Vaccine 2004, 23, 507–510. [Google Scholar] [CrossRef]
- Nelson, C.S.; Vera Cruz, D.; Su, M.; Xie, G.; Vandergrift, N.; Pass, R.F.; Forman, M.; Diener-West, M.; Koelle, K.; Arav-Boger, R.; et al. Intrahost dynamics of human cytomegalovirus variants acquired by seronegative glycoprotein B vaccinees. J. Virol. 2018, 93, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Frascaroli, G.; Lebbink, R.J.; Ostermann, E.; Brune, W. Human cytomegalovirus glycoprotein B variants affect viral entry, cell fusion, and genome stability. Proc. Natl. Acad. Sci. USA 2019, 116, 18021–18030. [Google Scholar] [CrossRef] [Green Version]
- Foglierini, M.; Marcandalli, J.; Perez, L. HCMV envelope glycoprotein diversity demystified. Front. Microbiol. 2019, 10, 1005. [Google Scholar] [CrossRef]
- Gardner, T.J.; Tortorella, D. Virion glycoprotein-mediated immune evasion by human cytomegalovirus: A sticky virus makes a slick getaway. Microbiol. Mol. Biol. Rev. 2016, 80, 663–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimamura, M.; Mach, M.; Britt, W.J. Human cytomegalovirus infection elicits a glycoprotein M (gM)/gN-specific virus-neutralizing antibody response. J. Virol. 2006, 80, 4591–4600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkhardt, C.; Himmelein, S.; Britt, W.; Winkler, T.; Mach, M. Glycoprotein N subtypes of human cytomegalovirus induce a strain-specific antibody response during natural infection. J. Gen. Virol. 2009, 90, 1951–1961. [Google Scholar] [CrossRef] [PubMed]
- Pati, S.K.; Novak, Z.; Purser, M.; Arora, N.; Mach, M.; Britt, W.J.; Boppana, S.B. Strain-specific neutralizing antibody responses against human cytomegalovirus envelope glycoprotein N. Clin. Vaccine Immunol. 2012, 19, 909–913. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Lanchy, J.-M.; Ryckman, B.J. Human cytomegalovirus gH/gL/gO promotes the fusion step of entry into all cell types, whereas gH/gL/UL128-131 broadens virus tropism through a distinct mechanism. J. Virol. 2015, 89, 8999–9009. [Google Scholar] [CrossRef] [Green Version]
- Puchhammer-Stöckl, E.; Görzer, I.; Zoufaly, A.; Jaksch, P.; Bauer, C.C.; Klepetko, W.; Popow-Kraupp, T. Emergence of multiple cytomegalovirus strains in blood and lung of lung transplant recipients. Transplantation 2006, 81, 187–194. [Google Scholar] [CrossRef]
- Roubalova, K.; Strunecky, O.; Vitek, A.; Zufanova, S.; Prochazka, B. Genetic variability of cytomegalovirus glycoprotein O in hematopoietic stem cell transplant recipients. Transpl. Infect. Dis. 2011, 13, 237–243. [Google Scholar] [CrossRef]
- Kalser, J.; Adler, B.; Mach, M.; Kropff, B.; Puchhammer-Stöckl, E.; Görzer, I. Differences in growth properties among two human cytomegalovirus glycoprotein O genotypes. Front. Microbiol. 2017, 8, 1609. [Google Scholar] [CrossRef]
- Brait, N.; Stögerer, T.; Kalser, J.; Adler, B.; Kunz, I.; Benesch, M.; Kropff, B.; Mach, M.; Puchhammer-Stöckl, E.; Görzer, I. Influence of human cytomegalovirus glycoprotein O polymorphism on the inhibitory effect of soluble forms of trimer- and pentamer-specific entry receptors. J. Virol. 2020, 94, 1–17. [Google Scholar] [CrossRef]
- Day, L.Z.; Stegmann, C.; Schultz, E.P.; Lanchy, J.-M.; Yu, Q.; Ryckman, B.J. Polymorphisms in human cytomegalovirus glycoprotein O (gO) exert epistatic influences on cell-free and cell-to-cell spread and antibody neutralization on gH epitopes. J. Virol. 2020, 94, e02051-19. [Google Scholar] [CrossRef]
- Adler, B.; Scrivano, L.; Ruzcics, Z.; Rupp, B.; Sinzger, C.; Koszinowski, U. Role of human cytomegalovirus UL131A in cell type-specific virus entry and release. J. Gen. Virol. 2006, 87, 2451–2460. [Google Scholar] [CrossRef] [PubMed]
- Straschewski, S.; Patrone, M.; Walther, P.; Gallina, A.; Mertens, T.; Frascaroli, G. Protein pUL128 of human cytomegalovirus Is necessary for monocyte infection and blocking of migration. J. Virol. 2011, 85, 5150–5158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerna, G.; Percivalle, E.; Lilleri, D.; Lozza, L.; Fornara, C.; Hahn, G.; Baldanti, F.; Revello, M.G. Dendritic-cell infection by human cytomegalovirus is restricted to strains carrying functional UL131-128 genes and mediates efficient viral antigen presentation to CD8+ T cells. J. Gen. Virol. 2005, 86, 275–284. [Google Scholar] [CrossRef]
- Urban, M.; Klein, M.; Britt, W.J.; Haßfurther, E.; Mach, M. Glycoprotein H of human cytomegalovirus is a major antigen for the neutralizing humoral immune response. J. Gen. Virol. 1996, 77, 1537–1547. [Google Scholar] [CrossRef]
- Freed, D.C.; Tang, Q.; Tang, A.; Li, F.; He, X.; Huang, Z.; Meng, W.; Xia, L.; Finnefrock, A.C.; Durr, E.; et al. Pentameric complex of viral glycoprotein H is the primary target for potent neutralization by a human cytomegalovirus vaccine. Proc. Natl. Acad. Sci. USA 2013, 110, 4997–5005. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, K.; Tokumoto, T.; Tanabe, K.; Shirakawa, H.; Hashimoto, K.; Kushida, N.; Yanagida, T.; Inoue, N.; Yamaguchi, O.; Toma, H.; et al. Association of the outcome of renal transplantation with antibody response to cytomegalovirus strain--specific glycoprotein H epitopes. Clin. Infect. Dis. 2007, 45, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Liu, L.; Tao, R.; Zheng, X.; Li, H.; Shang, S. Epidemiological characteristics of human cytomegalovirus infection and glycoprotein H genotype in Chinese children. Pediatr. Neonatol. 2020, 61, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Tao, R.; Zhang, X.; Shu, Q.; Gao, H.H.; Shang, S.Q.; Peng, Z.Y.; Li, H.M. Rapid and sensitive identification of glycoprotein H genotypes in clinical human cytomegalovirus samples. Jpn. J. Infect. Dis. 2015, 68, 135–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradowska, E.; Jabłońska, A.; Studzińska, M.; Kasztelewicz, B.; Zawilińska, B.; Wiśniewska-Ligier, M.; Dzierżanowska-Fangrat, K.; Woźniakowska-Gęsicka, T.; Kosz-Vnenchak, M.; Leśnikowski, Z.J. Cytomegalovirus glycoprotein H genotype distribution and the relationship with hearing loss in children. J. Med. Virol. 2014, 86, 1421–1427. [Google Scholar] [CrossRef]
- Paradowska, E.; Jabłońska, A.; Studzińska, M.; Kasztelewicz, B.; Wiśniewska-Ligier, M.; Dzierżanowska-Fangrat, K.; Woźniakowska-Gęsicka, T.; Czech-Kowalska, J. Distribution of the CMV glycoprotein gH/gL/gO and gH/gL/pUL128/pUL130/pUL131A complex variants and associated clinical manifestations in infants infected congenitally or postnatally. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glycoprotein | Length (Amino Acids) | Genotypes (Subtypes) | Most Variable Region | References |
|---|---|---|---|---|
| gB (UL55) | 907 | gB-1, gB-2, gB-3, gB-4, gB-5 | codons 26–70, gp55 cleavage site (codon 460) | [32,33,34] |
| gN (UL73) | 135 | gN-1, gN-2, gN-3 (gN-3a, gN-3b), gN-4 (gN-4a, gN-4b, gN-4c) | N-terminal region (codons 1–87) | [35,36] |
| gO (UL74) | 457–472 | gO-1 (gO-1a, gO-1b, gO-1c), gO-2 (gO-2a, gO-2b), gO-3, gO-4, gO-5 | N-terminal region (codons 1–98), codons 270–313 | [37,38,39,40,41] |
| gH (UL75) | 743 | gH-1, gH-2 | N-terminal region (codons 1–37) | [42] |
| Virus Strain | gB (UL55) 1 | gN (UL73) 2 | gO (UL74) 3 | gH (UL75) 4 | References |
|---|---|---|---|---|---|
| Towne | gB-1 | gN-4b | gO-4 | gH-2 | [33,36,40,42] |
| TB40E | gB-1 | gN-4c | gO-1c | gH-1 | [36,40] |
| AD169 | gB-2 | gN-1 | gO-1a | gH-1 | [33,36,40,42] |
| Toledo | gB-3 | gN-4c | gO-1c | gH-1 | [36,38,43] |
| VR1814 | gB-3 | gN-4c | gO-1c | gH-1 | N/A |
| Merlin | gB-1 | gN-4c | gO-5 | gH-2 | [44] |
| Transplant Recipient | ||||
| Continent | Dominant gB Genotype 1 | References | Dominant gH Genotype 1 | References |
| Asia | gB-1, gB-2 | [60,61,72,83,94,95,96,97] | gH-1 | [72,79,95] |
| Europe | gB-1, gB-2 | [124,125,126,149,150,151,152,153] | gH-1, gH-2 | [124,152] |
| North America | gB-1 | [100,102,103,104,105,148] | gH-1, gH-2 | [148] |
| Latin America | gB-1, gB-2 | [110,111,116,117,118,119] | ||
| Oceania | N/A | N/A | ||
| AIDS patient | ||||
| Continent | Dominant gB Genotype 1 | References | ||
| Asia | gB-1 | [98] | ||
| Europe | gB-2 | [59,127,128,129,130] | ||
| North America | gB-2 | [106,107] | ||
| Latin America | gB-1, gB-2, gB-3 | [120,121] | ||
| Oceania | gB-1, gB-2, gB-4 | [154] | ||
| Congenital CMV infection or pregnant woman | ||||
| Continent | Dominant gB Genotype 1 | References | Dominant gH Genotype 1 | References |
| Asia | gB-1 | [62,63,64,65,66,67,68,69,70,71,73,74,75,76,77,78,99] | gH-1 | [66,73,78,138,180,181] |
| Europe | gB-1 | [24,55,131,132,133,134,135,136,137,138,139,140,141,142,152] | gH-1, gH-2 | [55,133,142,152,182,183] |
| North America | gB-1, gB-2 | [108,109] | gH-1, gH-2 | [109] |
| Latin America | gB-2 | [112,122,123] | ||
| Oceania | gB-1 | [154] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-Y.; Valencia, S.M.; Pfeifer, S.P.; Jensen, J.D.; Kowalik, T.F.; Permar, S.R. Common Polymorphisms in the Glycoproteins of Human Cytomegalovirus and Associated Strain-Specific Immunity. Viruses 2021, 13, 1106. https://doi.org/10.3390/v13061106
Wang H-Y, Valencia SM, Pfeifer SP, Jensen JD, Kowalik TF, Permar SR. Common Polymorphisms in the Glycoproteins of Human Cytomegalovirus and Associated Strain-Specific Immunity. Viruses. 2021; 13(6):1106. https://doi.org/10.3390/v13061106
Chicago/Turabian StyleWang, Hsuan-Yuan, Sarah M. Valencia, Susanne P. Pfeifer, Jeffrey D. Jensen, Timothy F. Kowalik, and Sallie R. Permar. 2021. "Common Polymorphisms in the Glycoproteins of Human Cytomegalovirus and Associated Strain-Specific Immunity" Viruses 13, no. 6: 1106. https://doi.org/10.3390/v13061106
APA StyleWang, H. -Y., Valencia, S. M., Pfeifer, S. P., Jensen, J. D., Kowalik, T. F., & Permar, S. R. (2021). Common Polymorphisms in the Glycoproteins of Human Cytomegalovirus and Associated Strain-Specific Immunity. Viruses, 13(6), 1106. https://doi.org/10.3390/v13061106