
Organoids to Dissect Gastrointestinal Virus–Host Interactions: What Have We Learned?


Abstract
:1. Introduction
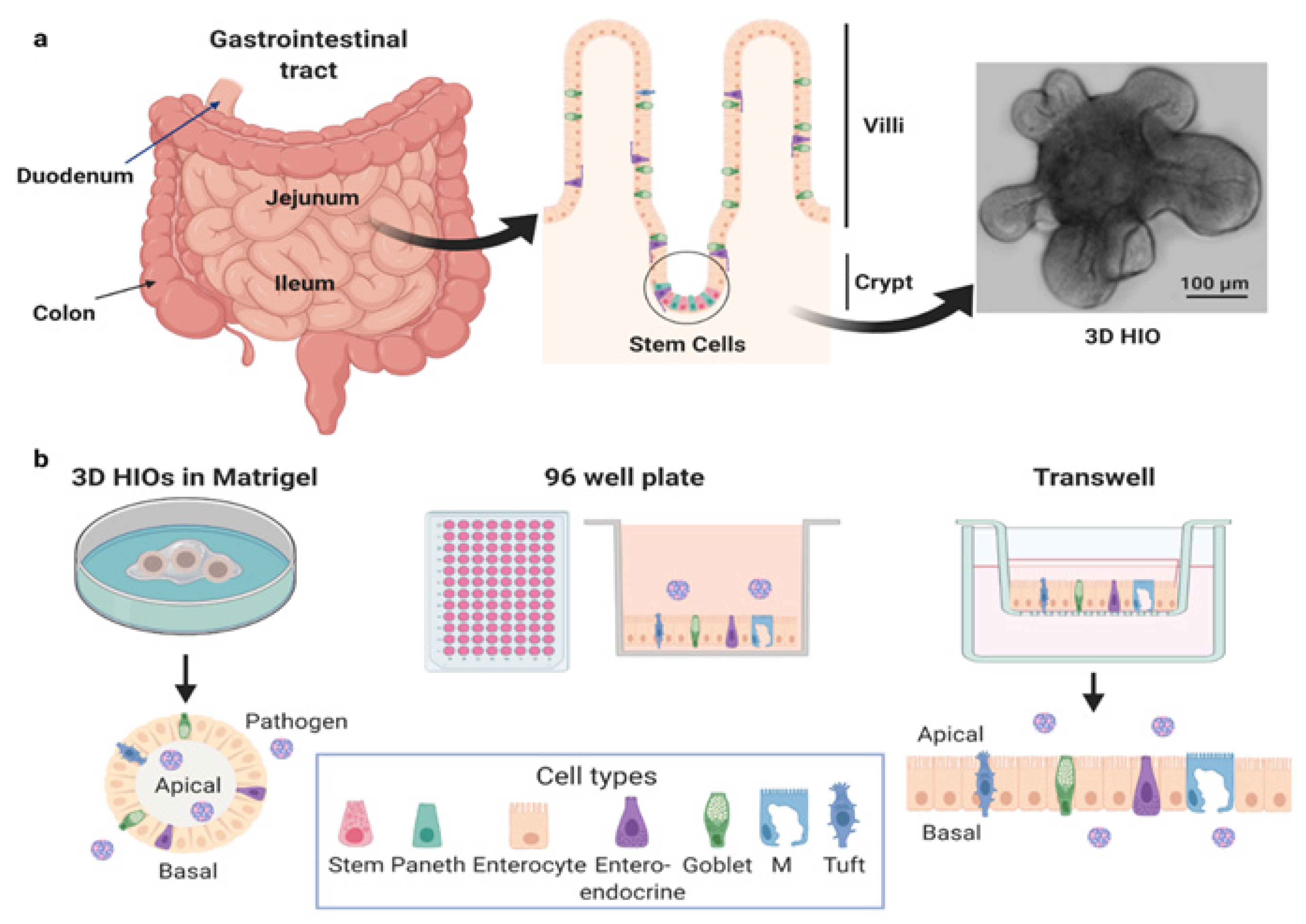
1.1. The Human Gastrointestinal Tract
1.2. Human Intestinal Organoids
1.3. Gastroenteritis Causing Viruses
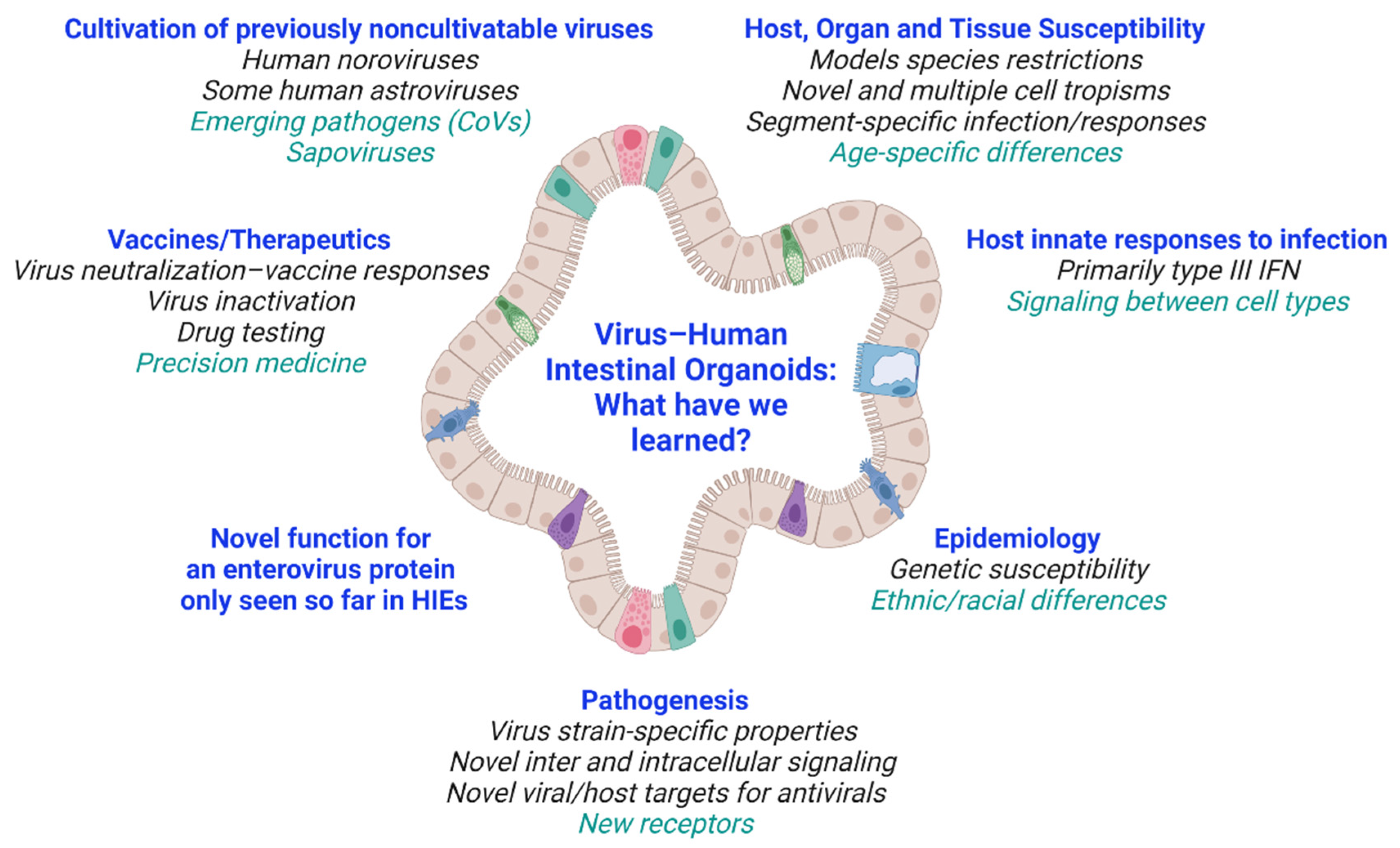
2. New Discoveries and Biology of Host–Microbe Interactions Revealed by HIO–Virus Studies
2.1. Human Viruses Infect More Intestinal Cell Types than Previously Recognized, and the State of Cell Differentiation Can Affect Infections
2.2. Host Cell Polarity and Virus Infections
2.3. Host Restrictions to Susceptibility to Viruses Are Recapitulated in HIOs
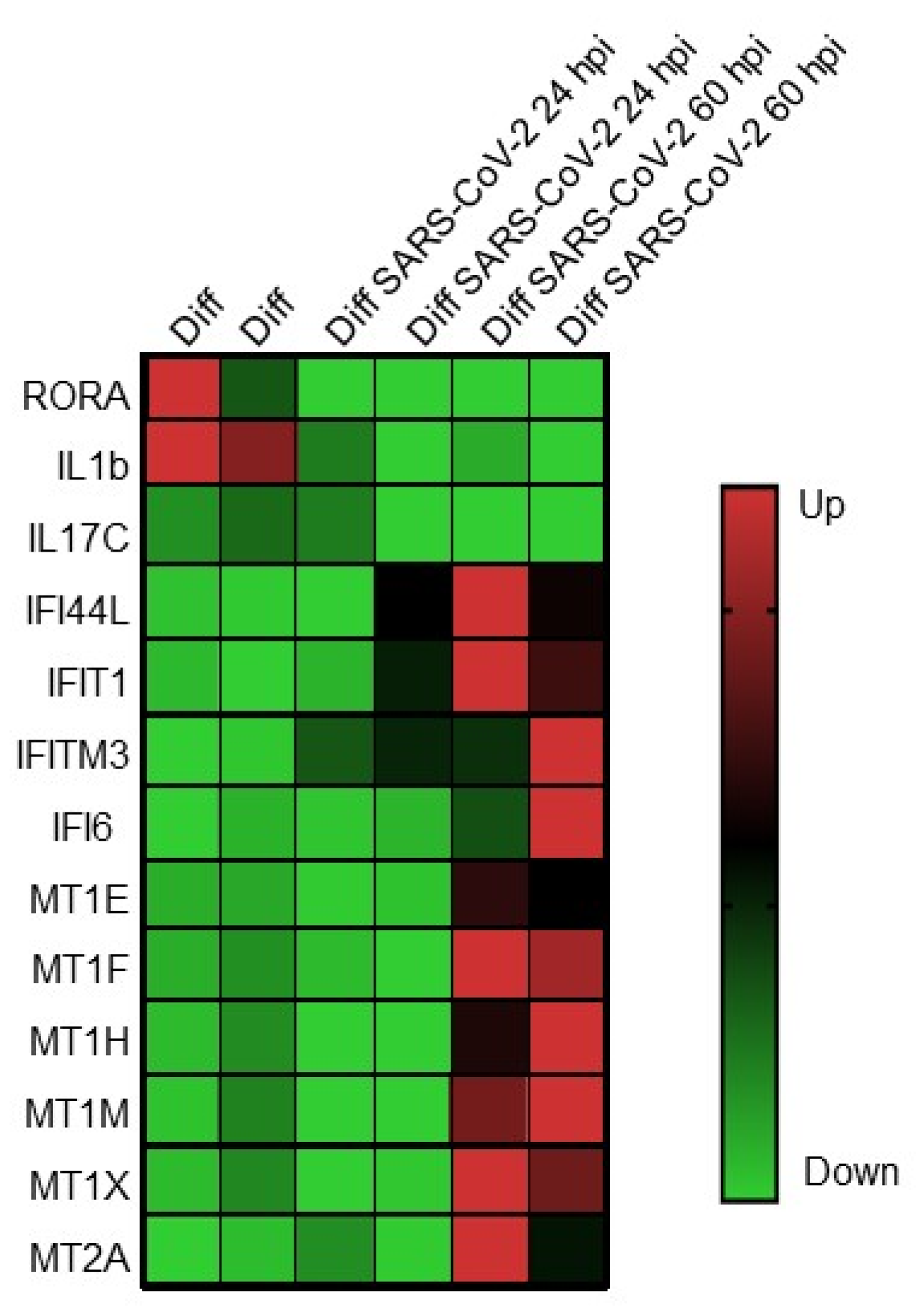
2.4. Host Innate Responses to Infection
2.5. Dissection of Mechanisms of Pathogenesis
2.6. Neutralizing Antibodies, Therapeutics, Vaccines, and Viral Inactivation Evaluated in HIOs
2.7. New Knowledge of Intestinal Biology Is Being Gained by Viral Infection Studies in HIOs
3. Summary and Current Questions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barker, N.; van Es, J.H.; Kuipers, J.; Kujala, P.; van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of Stem Cells in Small Intestine and Colon by Marker Gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.; Van Es, J.H.; Van den Brink, S.; Van Houdt, W.J.; Pronk, A.; Van Gorp, J.; Siersema, P.D.; et al. Long-term Expansion of Epithelial Organoids from Human Colon, Adenoma, Adenocarcinoma, and Barrett’s Epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 Stem Cells Build Crypt-villus Structures in vitro without a Mesenchymal Niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef]
- Sato, T.; Clevers, H. Growing Self-organizing Mini-guts from a Single Intestinal Stem Cell: Mechanism and Applications. Science 2013, 340, 1190–1194. [Google Scholar] [CrossRef] [Green Version]
- Saxena, K.; Blutt, S.E.; Ettayebi, K.; Zeng, X.L.; Broughman, J.R.; Crawford, S.E.; Karandikar, U.C.; Sastri, N.P.; Conner, M.E.; Opekun, A.R.; et al. Human Intestinal Enteroids: A New Model to Study Human Rotavirus Infection, Host Restriction, and Pathophysiology. J. Virol. 2016, 90, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.L.; Qu, L.; et al. Replication of Human Noroviruses in Stem Cell-derived Human Enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Lulla, V.; Dinan, A.M.; Hosmillo, M.; Chaudhry, Y.; Sherry, L.; Irigoyen, N.; Nayak, K.M.; Stonehouse, N.J.; Zilbauer, M.; Goodfellow, I.; et al. An Upstream Protein-coding Region in Enteroviruses Modulates Virus Infection in Gut Epithelial Cells. Nat. Microbiol. 2019, 4, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Holly, M.K.; Smith, J.G. Adenovirus Infection of Human Enteroids Reveals Interferon Sensitivity and Preferential Infection of Goblet Cells. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmberg, F.E.; Seidelin, J.B.; Yin, X.; Mead, B.E.; Tong, Z.; Li, Y.; Karp, J.M.; Nielsen, O.H. Culturing Human Intestinal Stem Cells for Regenerative Applications in the Treatment of Inflammatory Bowel Disease. EMBO Mol. Med. 2017, 9, 558–570. [Google Scholar] [CrossRef]
- Spence, J.R.; Mayhew, C.N.; Rankin, S.A.; Kuhar, M.F.; Vallance, J.E.; Tolle, K.; Hoskins, E.E.; Kalinichenko, V.V.; Wells, S.I.; Zorn, A.M.; et al. Directed Differentiation of Human Pluripotent Stem Cells into Intestinal Tissue in vitro. Nature 2011, 470, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Willyard, C. The Boom in Mini Stomachs, Brains, Breasts, Kidneys and More. Nature 2015, 523, 520–522. [Google Scholar] [CrossRef] [Green Version]
- Finkbeiner, S.R.; Zeng, X.L.; Utama, B.; Atmar, R.L.; Shroyer, N.F.; Estes, M.K. Stem Cell-derived Human Intestinal Organoids as an Infection Model for Rotaviruses. mBio 2012, 3, e00159-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Tan, M.; Zhong, W.; Xia, M.; Huang, P.; Jiang, X. Human Intestinal Organoids Express Histo-blood Group Antigens, Bind Norovirus VLPs, and Support Limited Norovirus Replication. Sci. Rep. 2017, 7, 12621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, S.; Hisaie, K.; Kurokawa, S.; Suzuki, A.; Sakon, N.; Uchida, Y.; Yuki, Y.; Kiyono, H. Human Norovirus Propagation in Human Induced Pluripotent Stem Cell-derived Intestinal Epithelial Cells. Cell Mol. Gastroenterol. Hepatol. 2019, 7, 686–688.e5. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Duan, X.; Yang, L.; Nilsson-Payant, B.E.; Wang, P.; Duan, F.; Tang, X.; Yaron, T.M.; Zhang, T.; Uhl, S.; et al. Identification of SARS-CoV-2 Inhibitors Using Lung and Colonic Organoids. Nature 2021, 589, 270–275. [Google Scholar] [CrossRef]
- Finkbeiner, S.R.; Hill, D.R.; Altheim, C.H.; Dedhia, P.H.; Taylor, M.J.; Tsai, Y.H.; Chin, A.M.; Mahe, M.M.; Watson, C.L.; Freeman, J.J.; et al. Transcriptome-wide Analysis Reveals Hallmarks of Human Intestine Development and Maturation in vitro and in vivo. Stem Cell Rep. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, M.A.; Sequeira, D.J.; McNeill, E.P.; Criss, Z.K., 2nd; Shroyer, N.F.; Speer, A.L. In vivo Transplantation of Human Intestinal Organoids Enhances Select Tight Junction Gene Expression. J. Surg. Res. 2021, 259, 500–508. [Google Scholar] [CrossRef]
- Watson, C.L.; Mahe, M.M.; Munera, J.; Howell, J.C.; Sundaram, N.; Poling, H.M.; Schweitzer, J.I.; Vallance, J.E.; Mayhew, C.N.; Sun, Y.; et al. An in vivo Model of Human Small Intestine Using Pluripotent Stem Cells. Nat. Med. 2014, 20, 1310–1314. [Google Scholar] [CrossRef] [Green Version]
- DeBerardinis, R.J.; Chandel, N.S. Fundamentals of Cancer Metabolism. Sci. Adv. 2016, 2, e1600200. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, A.; Kamphorst, J.J.; Markert, E.K.; Schug, Z.T.; Tardito, S.; Gottlieb, E. Cancer Metabolism at a Glance. J. Cell Sci. 2016, 129, 3367–3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolawole, A.O.; Mirabelli, C.; Hill, D.R.; Svoboda, S.A.; Janowski, A.B.; Passalacqua, K.D.; Rodriguez, B.N.; Dame, M.K.; Freiden, P.; Berger, R.P.; et al. Astrovirus Replication in Human Intestinal Enteroids Reveals Multi-cellular Tropism and an Intricate Host Innate Immune Landscape. PLoS Pathog. 2019, 15, e1008057. [Google Scholar] [CrossRef] [Green Version]
- Heo, I.; Dutta, D.; Schaefer, D.A.; Iakobachvili, N.; Artegiani, B.; Sachs, N.; Boonekamp, K.E.; Bowden, G.; Hendrickx, A.P.A.; Willems, R.J.L.; et al. Modelling Cryptosporidium Infection in Human Small Intestinal and Lung Organoids. Nat. Microbiol. 2018, 3, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Nickerson, K.P.; Senger, S.; Zhang, Y.; Lima, R.; Patel, S.; Ingano, L.; Flavahan, W.A.; Kumar, D.K.V.; Fraser, C.M.; Faherty, C.S.; et al. Salmonella Typhi Colonization Provokes Extensive Transcriptional Changes Aimed at Evading Host Mucosal Immune Defense during Early Infection of Human Intestinal Tissue. EBioMedicine 2018, 31, 92–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duizer, E.; Schwab, K.J.; Neill, F.H.; Atmar, R.L.; Koopmans, M.P.G.; Estes, M.K. Laboratory Efforts to Cultivate Noroviruses. J. Gen. Virol. 2004, 85, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Mestas, J.; Hughes, C.C. Of Mice and Not Men: Differences between Mouse and Human Immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [Green Version]
- Hermant, P.; Demarez, C.; Mahlakoiv, T.; Staeheli, P.; Meuleman, P.; Michiels, T. Human but not Mouse Hepatocytes Respond to Interferon-lambda in vivo. PLoS ONE 2014, 9, e87906. [Google Scholar] [CrossRef]
- Collaborators, G.B.D.D.D. Estimates of the Global, Regional, and National Morbidity, Mortality, and Aetiologies of Diarrhoea in 195 Countries: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Infect Dis 2018, 18, 1211–1228. [Google Scholar] [CrossRef] [Green Version]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Steele, A.D.; Duque, J.; Parashar, U.D.; Network, W.H.-C.G.R.S. 2008 Estimate of Worldwide Rotavirus-associated Mortality in Children Younger Than 5 Years Before the Introduction of Universal Rotavirus Vaccination Programmes: A Systematic Review and Meta-analysis. Lancet Infect. Dis. 2012, 12, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.M.; Hall, A.J.; Robinson, A.E.; Verhoef, L.; Premkumar, P.; Parashar, U.D.; Koopmans, M.; Lopman, B.A. Global Prevalence of Norovirus in Cases of Gastroenteritis: A Systematic Review and Meta-analysis. Lancet Infect. Dis. 2014, 14, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.L.; Neill, F.H.; Estes, M.K.; Munoz, F.M.; Cameron, A.; DuPont, H.L.; Atmar, R.L. Noroviruses: The Most Common Pediatric Viral Enteric Pathogen at a Large University Hospital after Introduction of Rotavirus Vaccination. J. Pediatric Infect. Dis. Soc. 2013, 2, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Payne, D.C.; Vinje, J.; Szilagyi, P.G.; Edwards, K.M.; Staat, M.A.; Weinberg, G.A.; Hall, C.B.; Chappell, J.; Bernstein, D.I.; Curns, A.T.; et al. Norovirus and Medically Attended Gastroenteritis in U.S. Children. N. Engl. J. Med. 2013, 368, 1121–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.L.; Bosch, A.; Pinto, R.M.; Guix, S. Epidemiology of Classic and Novel Human Astrovirus: Gastroenteritis and Beyond. Viruses 2017, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Wang, Q.; Katayama, K.; Saif, L.J. Comprehensive Review of Human Sapoviruses. Clin. Microbiol. Rev. 2015, 28, 32–53. [Google Scholar] [CrossRef] [Green Version]
- Redd, W.D.; Zhou, J.C.; Hathorn, K.E.; McCarty, T.R.; Bazarbashi, A.N.; Thompson, C.C.; Shen, L.; Chan, W.W. Prevalence and Characteristics of Gastrointestinal Symptoms in Patients with Severe Acute Respiratory Syndrome Coronavirus 2 Infection in the United States: A Multicenter Cohort Study. Gastroenterology 2020, 159, 765–767.e2. [Google Scholar] [CrossRef]
- Livanos, A.E.; Jha, D.; Cossarini, F.; Gonzalez-Reiche, A.S.; Tokuyama, M.; Aydillo, T.; Parigi, T.L.; Ramos, I.; Dunleavy, K.; Lee, B.; et al. Gastrointestinal Involvement Attenuates COVID-19 Severity and Mortality. medRxiv 2020. [Google Scholar] [CrossRef]
- Lamers, M.M.; Beumer, J.; van der Vaart, J.; Knoops, K.; Puschhof, J.; Breugem, T.I.; Ravelli, R.B.G.; Paul van Schayck, J.; Mykytyn, A.Z.; Duimel, H.Q.; et al. SARS-CoV-2 Productively Infects Human Gut Enterocytes. Science 2020, 369, 50–54. [Google Scholar] [CrossRef]
- Leung, W.K.; To, K.F.; Chan, P.K.; Chan, H.L.; Wu, A.K.; Lee, N.; Yuen, K.Y.; Sung, J.J. Enteric Involvement of Severe Acute Respiratory Syndrome-associated Coronavirus Infection. Gastroenterology 2003, 125, 1011–1017. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.H.; Poon, L.L.; Cheng, V.C.; Guan, Y.; Hung, I.F.; Kong, J.; Yam, L.Y.; Seto, W.H.; Yuen, K.Y.; Peiris, J.S. Detection of SARS Coronavirus in Patients with Suspected SARS. Emerg. Infect. Dis. 2004, 10, 294–299. [Google Scholar] [CrossRef] [Green Version]
- Assiri, A.; Al-Tawfiq, J.A.; Al-Rabeeah, A.A.; Al-Rabiah, F.A.; Al-Hajjar, S.; Al-Barrak, A.; Flemban, H.; Al-Nassir, W.N.; Balkhy, H.H.; Al-Hakeem, R.F.; et al. Epidemiological, Demographic, and Clinical Characteristics of 47 Cases of Middle East Respiratory Syndrome Coronavirus Disease from Saudi Arabia: A Descriptive Study. Lancet Infect. Dis. 2013, 13, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brunink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 Novel Coronavirus (2019-nCoV) by Real-time RT-PCR. Eurosurveillance 2020, 25. [Google Scholar] [CrossRef] [Green Version]
- Saxena, K.; Simon, L.M.; Zeng, X.L.; Blutt, S.E.; Crawford, S.E.; Sastri, N.P.; Karandikar, U.C.; Ajami, N.J.; Zachos, N.C.; Kovbasnjuk, O.; et al. A Paradox of Transcriptional and Functional Innate Interferon Responses of Human Intestinal Enteroids to Enteric Virus Infection. Proc. Natl. Acad. Sci. USA 2017, 114, E570–E579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, N.; Hu, L.; Ding, S.; Sanyal, M.; Zhao, B.; Sankaran, B.; Ramani, S.; McNeal, M.; Yasukawa, L.L.; Song, Y.; et al. Human VP8* mAbs Neutralize Rotavirus Selectively in Human Intestinal Epithelial Cells. J. Clin. Invest. 2019, 129, 3839–3851. [Google Scholar] [CrossRef]
- Ettayebi, K.; Tenge, V.R.; Cortes-Penfield, N.W.; Crawford, S.E.; Neill, F.H.; Zeng, X.L.; Yu, X.; Ayyar, B.V.; Burrin, D.; Ramani, S.; et al. New Insights and Enhanced Human Norovirus Cultivation in Human Intestinal Enteroids. mSphere 2021, 6. [Google Scholar] [CrossRef]
- Lin, S.C.; Qu, L.; Ettayebi, K.; Crawford, S.E.; Blutt, S.E.; Robertson, M.J.; Zeng, X.L.; Tenge, V.R.; Ayyar, B.V.; Karandikar, U.C.; et al. Human Norovirus Exhibits Strain-specific Sensitivity to Host Interferon Pathways in Human Intestinal Enteroids. Proc. Natl. Acad. Sci. USA 2020, 117, 23782–23793. [Google Scholar] [CrossRef] [PubMed]
- Hosmillo, M.; Chaudhry, Y.; Nayak, K.; Sorgeloos, F.; Koo, B.K.; Merenda, A.; Lillestol, R.; Drumright, L.; Zilbauer, M.; Goodfellow, I. Norovirus Replication in Human Intestinal Epithelial Cells is Restricted by the Interferon-induced JAK/STAT Signaling Pathway and RNA Polymerase II-mediated Transcriptional Responses. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, C.G.; Bolock, A.M.; Ma, C.; Luke, C.J.; Good, M.; Coyne, C.B. Enteroviruses Infect Human Enteroids and Induce Antiviral Signaling in a Cell Lineage-specific Manner. Proc. Natl. Acad. Sci. USA 2017, 114, 1672–1677. [Google Scholar] [CrossRef] [Green Version]
- Good, C.; Wells, A.I.; Coyne, C.B. Type III Interferon Signaling Restricts Enterovirus 71 Infection of Goblet Cells. Sci. Adv. 2019, 5, eaau4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagbom, M.; Istrate, C.; Engblom, D.; Karlsson, T.; Rodriguez-Diaz, J.; Buesa, J.; Taylor, J.A.; Loitto, V.M.; Magnusson, K.E.; Ahlman, H.; et al. Rotavirus Stimulates Release of Serotonin (5-HT) from Human Enterochromaffin Cells and Activates Brain Structures Involved in Nausea and Vomiting. PLoS Pathog. 2011, 7, e1002115. [Google Scholar] [CrossRef] [PubMed]
- Karandikar, U.C.; Crawford, S.E.; Ajami, N.J.; Murakami, K.; Kou, B.; Ettayebi, K.; Papanicolaou, G.A.; Jongwutiwes, U.; Perales, M.A.; Shia, J.; et al. Detection of Human Norovirus in Intestinal Biopsies from Immunocompromised Transplant Patients. J. Gen. Virol. 2016, 97, 2291–2300. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.Y.; Blutt, S.E.; Crawford, S.E.; Ettayebi, K.; Zeng, X.L.; Saxena, K.; Ramani, S.; Karandikar, U.C.; Zachos, N.C.; Estes, M.K. Human Intestinal Enteroids: New Models to Study Gastrointestinal Virus Infections. Methods Mol. Biol. 2019, 1576, 229–247. [Google Scholar] [CrossRef]
- Davidson, G.P.; Barnes, G.L. Structural and Functional Abnormalities of the Small Intestine in Infants and Young Children with Rotavirus Enteritis. Acta Paediatr. Scand. 1979, 68, 181–186. [Google Scholar] [CrossRef]
- Green, K.Y.; Kaufman, S.S.; Nagata, B.M.; Chaimongkol, N.; Kim, D.Y.; Levenson, E.A.; Tin, C.M.; Yardley, A.B.; Johnson, J.A.; Barletta, A.B.F.; et al. Human Norovirus Targets Enteroendocrine Epithelial Cells in the Small Intestine. Nat. Commun. 2020, 11, 2759. [Google Scholar] [CrossRef]
- Chang-Graham, A.L.; Danhof, H.A.; Engevik, M.A.; Tomaro-Duchesneau, C.; Karandikar, U.C.; Estes, M.K.; Versalovic, J.; Britton, R.A.; Hyser, J.M. Human Intestinal Enteroids with Inducible Neurogenin-3 Expression as a Novel Model of Gut Hormone Secretion. Cell Mol Gastroenterol. Hepatol. 2019, 8, 209–229. [Google Scholar] [CrossRef] [Green Version]
- Hagbom, M.; Novak, D.; Ekstrom, M.; Khalid, Y.; Andersson, M.; Lindh, M.; Nordgren, J.; Svensson, L. Ondansetron Treatment Reduces Rotavirus Symptoms-A Randomized Double-blinded Placebo-controlled Trial. PLoS ONE 2017, 12, e0186824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.G.; Silvestry, M.; Lindert, S.; Lu, W.; Nemerow, G.R.; Stewart, P.L. Insight into the Mechanisms of Adenovirus Capsid Disassembly from Studies of Defensin Neutralization. PLoS Pathog. 2010, 6, e1000959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanifer, M.L.; Kee, C.; Cortese, M.; Zumaran, C.M.; Triana, S.; Mukenhirn, M.; Kraeusslich, H.G.; Alexandrov, T.; Bartenschlager, R.; Boulant, S. Critical Role of Type III Interferon in Controlling SARS-CoV-2 Infection in Human Intestinal Epithelial Cells. Cell Rep. 2020, 32, 107863. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, C.; Liu, X.; Chiu, M.C.; Zhao, X.; Wang, D.; Wei, Y.; Lee, A.; Zhang, A.J.; Chu, H.; et al. Infection of Bat and Human Intestinal Organoids by SARS-CoV-2. Nat. Med. 2020, 26, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Zang, R.; Gomez Castro, M.F.; McCune, B.T.; Zeng, Q.; Rothlauf, P.W.; Sonnek, N.M.; Liu, Z.; Brulois, K.F.; Wang, X.; Greenberg, H.B.; et al. TMPRSS2 and TMPRSS4 Promote SARS-CoV-2 Infection of Human Small Intestinal Enterocytes. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef]
- Xiao, F.; Tang, M.; Shan, H. Reply to Letter to the Editor: Presence of SARS-Coronavirus-2 in the Ileal Mucosa: Another Evidence for Infection of GI Tract by This Virus (GASTRO-D-20-01382). Gastroenterology 2020. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.U.; Brown, J.W.; Li, Q.K.; Bayguinov, P.O.; Wang, J.S.; Mills, J.C.; Collaborators; Whelan, S.P.J.; Fitzpatrick, J.A.J.; Meeker, A.K. Tropism of SARS-CoV-2 for Barrett’s Esophagus May Increase Susceptibility to Developing COVID-19. Gastroenterology 2021. [Google Scholar] [CrossRef]
- Gu, J.; Gong, E.; Zhang, B.; Zheng, J.; Gao, Z.; Zhong, Y.; Zou, W.; Zhan, J.; Wang, S.; Xie, Z.; et al. Multiple Organ Infection and the Pathogenesis of SARS. J. Exp. Med. 2005, 202, 415–424. [Google Scholar] [CrossRef] [PubMed]
- VanDussen, K.L.; Marinshaw, J.M.; Shaikh, N.; Miyoshi, H.; Moon, C.; Tarr, P.I.; Ciorba, M.A.; Stappenbeck, T.S. Development of an Enhanced Human Gastrointestinal Epithelial Culture System to Facilitate Patient-based Assays. Gut 2015, 64, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Leslie, J.L.; Weddle, E.; Yum, L.K.; Lin, Y.; Jenior, M.L.; Lee, B.; Ma, J.Z.; Kirkpatrick, B.D.; Nayak, U.; Platts-Mills, J.A.; et al. Lewis Blood Group Antigens are Associated with Altered Susceptibility to Shigellosis. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Hutson, A.M.; Atmar, R.L.; Graham, D.Y.; Estes, M.K. Norwalk Virus Infection and Disease is Associated with ABO Histo-blood Group Type. J. Infect. Dis. 2002, 185, 1335–1337. [Google Scholar] [CrossRef]
- Ayouni, S.; Estienney, M.; Sdiri-Loulizi, K.; Ambert-Balay, K.; de Rougemont, A.; Aho, S.; Hammami, S.; Aouni, M.; Guediche, M.N.; Pothier, P.; et al. Relationship between GII.3 Norovirus Infections and Blood Group Antigens in Young Children in Tunisia. Clin. Microbiol. Infect. 2015, 21, 874.e1–874.e8. [Google Scholar] [CrossRef] [Green Version]
- Ruvoen-Clouet, N.; Belliot, G.; Le Pendu, J. Noroviruses and Histo-blood Groups: The Impact of Common Host Genetic Polymorphisms on Virus Transmission and Evolution. Rev. Med. Virol. 2013, 23, 355–366. [Google Scholar] [CrossRef]
- Haga, K.; Ettayebi, K.; Tenge, V.R.; Karandikar, U.C.; Lewis, M.A.; Lin, S.C.; Neill, F.H.; Ayyar, B.V.; Zeng, X.L.; Larson, G.; et al. Genetic Manipulation of Human Intestinal Enteroids Demonstrates the Necessity of a Functional Fucosyltransferase 2 Gene for Secretor-dependent Human Norovirus Infection. mBio 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Frias, A.H.; Jones, R.M.; Fifadara, N.H.; Vijay-Kumar, M.; Gewirtz, A.T. Rotavirus-induced IFN-beta Promotes Anti-viral Signaling and Apoptosis That Modulate Viral Replication in Intestinal Epithelial Cells. Innate Immun. 2012, 18, 294–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, A.; Rothenberg, M.E.; Mukherjee, G.; Feng, N.; Kalisky, T.; Nair, N.; Johnstone, I.M.; Clarke, M.F.; Greenberg, H.B. Innate Immune Response to Homologous Rotavirus Infection in the Small Intestinal Villous Epithelium at Single-cell Resolution. Proc. Natl. Acad. Sci. USA 2012, 109, 20667–20672. [Google Scholar] [CrossRef] [Green Version]
- Pelaseyed, T.; Bergstrom, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.; Schutte, A.; van der Post, S.; Svensson, F.; Rodriguez-Pineiro, A.M.; Nystrom, E.E.; et al. The Mucus and Mucins of the Goblet Cells and Enterocytes Provide the First Defense Line of the Gastrointestinal Tract and Interact with the Immune System. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.R.; Silva, J.; Sundaram, M.; Ellingson, M.K.; Mao, T.; Oh, J.E.; Israelow, B.; et al. Longitudinal Analyses Reveal Immunological Misfiring in Severe COVID-19. Nature 2020, 584, 463–469. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Hsin-Hui, H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.; Madduri, D.; Stock, A.; et al. An Inflammatory Cytokine Signature Helps Predict COVID-19 Severity and Death. medRxiv 2020. [Google Scholar] [CrossRef]
- Rajan, A.; Robertson, M.J.; Carter, H.E.; Poole, N.M.; Clark, J.R.; Green, S.I.; Criss, Z.K.; Zhao, B.; Karandikar, U.; Xing, Y.; et al. Enteroaggregative E. coli Adherence to Human Heparan Sulfate Proteoglycans Drives Segment and Host Specific Responses to Infection. PLoS Pathog. 2020, 16, e1008851. [Google Scholar] [CrossRef]
- Dekkers, J.F.; Wiegerinck, C.L.; de Jonge, H.R.; Bronsveld, I.; Janssens, H.M.; de Winter-de Groot, K.M.; Brandsma, A.M.; de Jong, N.W.; Bijvelds, M.J.; Scholte, B.J.; et al. A Functional CFTR Assay Using Primary Cystic Fibrosis Intestinal Organoids. Nat. Med. 2013, 19, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Hyser, J.M.; Estes, M.K. Pathophysiological Consequences of Calcium-conducting Viroporins. Annu. Rev. Virol. 2015, 2, 473–496. [Google Scholar] [CrossRef]
- Morris, A.P.; Scott, J.K.; Ball, J.M.; Zeng, C.Q.; O’Neal, W.K.; Estes, M.K. NSP4 Elicits Age-dependent Diarrhea and Ca(2+)mediated I(-) Influx into Intestinal Crypts of CF Mice. Am. J. Physiol. 1999, 277, G431–G444. [Google Scholar] [CrossRef] [PubMed]
- Ball, J.M.; Tian, P.; Zeng, C.Q.; Morris, A.P.; Estes, M.K. Age-dependent Diarrhea Induced by a Rotaviral Nonstructural Glycoprotein. Science 1996, 272, 101–104. [Google Scholar] [CrossRef]
- Pham, T.; Perry, J.L.; Dosey, T.L.; Delcour, A.H.; Hyser, J.M. The Rotavirus NSP4 Viroporin Domain is a Calcium-conducting Ion Channel. Sci. Rep. 2017, 7, 43487. [Google Scholar] [CrossRef]
- Crawford, S.E.; Hyser, J.M.; Utama, B.; Estes, M.K. Autophagy Hijacked through Viroporin-activated Calcium/calmodulin-dependent Kinase Kinase-beta Signaling is Required for Rotavirus Replication. Proc. Natl. Acad. Sci. USA 2012, 109, E3405–E3413. [Google Scholar] [CrossRef] [Green Version]
- Chang-Graham, A.L.; Perry, J.L.; Engevik, M.A.; Engevik, K.A.; Scribano, F.J.; Gebert, J.T.; Danhof, H.A.; Nelson, J.C.; Kellen, J.S.; Strtak, A.C.; et al. Rotavirus Induces Intercellular Calcium Waves through ADP Signaling. Science 2020, 370. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, G.; Ettayebi, K.; Atmar, R.L.; Bombardi, R.G.; Kose, N.; Estes, M.K.; Crowe, J.E., Jr. Human Monoclonal Antibodies That Neutralize Pandemic GII.4 Noroviruses. Gastroenterology 2018, 155, 1898–1907. [Google Scholar] [CrossRef] [Green Version]
- Atmar, R.L.; Ettayebi, K.; Ayyar, B.V.; Neill, F.H.; Braun, R.P.; Ramani, S.; Estes, M.K. Comparison of Microneutralization and Histo-blood Group Antigen-blocking Assays for Functional Norovirus Antibody Detection. J. Infect. Dis. 2020, 221, 739–743. [Google Scholar] [CrossRef]
- Koromyslova, A.D.; Morozov, V.A.; Hefele, L.; Hansman, G.S. Human Norovirus Neutralized by a Monoclonal Antibody Targeting the Histo-blood Group Antigen Pocket. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindesmith, L.C.; McDaniel, J.R.; Changela, A.; Verardi, R.; Kerr, S.A.; Costantini, V.; Brewer-Jensen, P.D.; Mallory, M.L.; Voss, W.N.; Boutz, D.R.; et al. Sera Antibody Repertoire Analyses Reveal Mechanisms of Broad and Pandemic Strain Neutralizing Responses after Human Norovirus Vaccination. Immunity 2019, 50, 1530–1541.e8. [Google Scholar] [CrossRef] [Green Version]
- Costantini, V.; Morantz, E.K.; Browne, H.; Ettayebi, K.; Zeng, X.L.; Atmar, R.L.; Estes, M.K.; Vinje, J. Human Norovirus Replication in Human Intestinal Enteroids as Model to Evaluate Virus Inactivation. Emerg. Infect. Dis. 2018, 24, 1453–1464. [Google Scholar] [CrossRef] [Green Version]
- Blutt, S.E.; Crawford, S.E.; Ramani, S.; Zou, W.Y.; Estes, M.K. Engineered Human Gastrointestinal Cultures to Study the Microbiome and Infectious Diseases. Cell Mol. Gastroenterol. Hepatol. 2018, 5, 241–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, K.; Tenge, V.R.; Karandikar, U.C.; Lin, S.C.; Ramani, S.; Ettayebi, K.; Crawford, S.E.; Zeng, X.L.; Neill, F.H.; Ayyar, B.V.; et al. Bile Acids and Ceramide Overcome the Entry Restriction for GII.3 Human Norovirus Replication in Human Intestinal Enteroids. Proc. Natl. Acad. Sci. USA 2020, 117, 1700–1710. [Google Scholar] [CrossRef]
- Randazzo, W.; Costantini, V.; Morantz, E.K.; Vinje, J. Human Intestinal Enteroids to Evaluate Human Norovirus GII.4 Inactivation by Aged-green Tea. Front Microbiol. 2020, 11, 1917. [Google Scholar] [CrossRef] [PubMed]
- de Lau, W.; Kujala, P.; Schneeberger, K.; Middendorp, S.; Li, V.S.; Barker, N.; Martens, A.; Hofhuis, F.; DeKoter, R.P.; Peters, P.J.; et al. Peyer’s Patch M Cells Derived from Lgr5(+) Stem Cells Require SpiB and are Induced by RankL in Cultured "Miniguts". Mol. Cell Biol. 2012, 32, 3639–3647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouch, J.D.; Scott, A.; Lei, N.Y.; Solorzano-Vargas, R.S.; Wang, J.; Hanson, E.M.; Kobayashi, M.; Lewis, M.; Stelzner, M.G.; Dunn, J.C.; et al. Development of Functional Microfold (M) Cells from Intestinal Stem Cells in Primary Human Enteroids. PLoS ONE 2016, 11, e0148216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasciano, A.C.; Blutt, S.E.; Estes, M.K.; Mecsas, J. Induced Differentiation of M Cell-like Cells in Human Stem Cell-derived Ileal Enteroid Monolayers. J. Vis. Exp. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Song, Y.; Brulois, K.F.; Pan, J.; Co, J.Y.; Ren, L.; Feng, N.; Yasukawa, L.L.; Sanchez-Tacuba, L.; Wosen, J.E.; et al. Retinoic Acid and Lymphotoxin Signaling Promote Differentiation of Human Intestinal M Cells. Gastroenterology 2020, 159, 214–226.e1. [Google Scholar] [CrossRef]
- Marion, O.; Lhomme, S.; Nayrac, M.; Dubois, M.; Pucelle, M.; Requena, M.; Migueres, M.; Abravanel, F.; Peron, J.M.; Carrere, N.; et al. Hepatitis E Virus Replication in Human Intestinal Cells. Gut 2020, 69, 901–910. [Google Scholar] [CrossRef]
- Noel, G.; Baetz, N.W.; Staab, J.F.; Donowitz, M.; Kovbasnjuk, O.; Pasetti, M.F.; Zachos, N.C. A Primary Human Macrophage-enteroid Co-culture Model to Investigate Mucosal Gut Physiology and Host-pathogen Interactions. Sci. Rep. 2017, 7, 45270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreurs, R.; Baumdick, M.E.; Sagebiel, A.F.; Kaufmann, M.; Mokry, M.; Klarenbeek, P.L.; Schaltenberg, N.; Steinert, F.L.; van Rijn, J.M.; Drewniak, A.; et al. Human Fetal TNF-alpha-cytokine-producing CD4(+) Effector Memory T Cells Promote Intestinal Development and Mediate Inflammation Early in Life. Immunity 2019, 50, 462–476.e8. [Google Scholar] [CrossRef] [Green Version]
- Fofanova, T.Y.; Stewart, C.J.; Auchtung, J.M.; Wilson, R.L.; Britton, R.A.; Grande-Allen, K.J.; Estes, M.K.; Petrosino, J.F. A Novel Human Enteroid-anaerobe Co-culture System to Study Microbial-host Interaction under Physiological Hypoxia. BioRxiv 2019. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Y.J.; Yoon, J.Y.; Kemmitt, J.; Wright, C.; Schneider, K.; Sphabmixay, P.; Hernandez-Gordillo, V.; Holcomb, S.J.; Bhushan, B.; et al. Primary Human Colonic Mucosal Barrier Crosstalk with Super Oxygen-sensitive Faecalibacterium prausnitzii in Continuous Culture. Med (N Y) 2021, 2, 74–98.e9. [Google Scholar] [CrossRef]
- Hewes, S.A.; Wilson, R.L.; Estes, M.K.; Shroyer, N.F.; Blutt, S.E.; Grande-Allen, K.J. In vitro Models of the Small Intestine: Engineering Challenges and Engineering Solutions. Tissue Eng. Part B Rev. 2020, 26, 313–326. [Google Scholar] [CrossRef]
- Sugimoto, S.; Kobayashi, E.; Fujii, M.; Ohta, Y.; Arai, K.; Matano, M.; Ishikawa, K.; Miyamoto, K.; Toshimitsu, K.; Takahashi, S.; et al. An Organoid-based Organ-repurposing Approach to Treat Short Bowel Syndrome. Nature 2021. [Google Scholar] [CrossRef]
- Sunuwar, L.; Yin, J.; Kasendra, M.; Karalis, K.; Kaper, J.; Fleckenstein, J.; Donowitz, M. Mechanical Stimuli Affect Escherichia coli Heat-stable Enterotoxin-cyclic GMP Signaling in a Human Enteroid Intestine-chip Model. Infect. Immun. 2020, 88. [Google Scholar] [CrossRef]
- Wilson, R.L.; Hewes, S.A.; Rajan, A.; Lin, S.C.; Bomidi, C.; Iida, T.; Estes, M.K.; Maresso, A.W.; Grande-Allen, K.J. A Millifluidic Perfusion Cassette for Studying the Pathogenesis of Enteric Infections Using ex-vivo Organoids. Ann. Biomed. Eng. 2021. [Google Scholar] [CrossRef]
- Lussier, C.; Basora, N.; Bouatrouss, Y.; Beaulieu, J.F. Integrins as Mediators of Epithelial Cell-matrix Interactions in the Human Small Intestinal Mucosa. Microsc. Res. Tech. 2000, 51, 169–178. [Google Scholar] [CrossRef]
- Hofmann, C.; Obermeier, F.; Artinger, M.; Hausmann, M.; Falk, W.; Schoelmerich, J.; Rogler, G.; Grossmann, J. Cell-cell Contacts Prevent Anoikis in Primary Human Colonic Epithelial Cells. Gastroenterology 2007, 132, 587–600. [Google Scholar] [CrossRef]
- Sachs, N.; Tsukamoto, Y.; Kujala, P.; Peters, P.J.; Clevers, H. Intestinal Epithelial Organoids Fuse to Form Self-organizing Tubes in Floating Collagen Gels. Development 2017, 144, 1107–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Virus | Supports Virus Replication | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Previously Noncultivatable | Undifferentiated/ Proliferative | Differentiated (Adult) | Susceptible Cell Type(s) | Route of Infection | Route of Release | IFN Response (Predominant) | Replication | References | ||
| Human Rotavirus | No | Poorly | D, J, I, C $ | Enterocyte, Enteroendocrine | Baolateral > Apical | NT | Type III | [5,41,42] | ||
| Human Norovirus | Yes | No | D, J, I, not C | Enterocyte, Enteroendocrine | Apical ** | NT | Type III | neutralized by monoclonal antibodies | [6,43,44,45] | |
| Human Astrovirus | Some strains | D, J, I, C # | D, J ** | Progenitor, Enterocyte, Goblet | Apical ** | NT | Type I and Type III | inhibited by heat, 2CMC *; increased by ruxolitinib *** | [21] | |
| Human Adenoviruses | Enteric | No | I ** | I ** | Enterocyte | Apical ** | NT | |||
| Respiratory | No | I ** | I ** | Goblet | Apical ** | NT | neutralized by enteric α-defensin HD5 | [8] | ||
| Enteroviruses | Enterovirus 11 | No | NT | Fetal ** | Enterocyte, Enteroendocrine | Basolateral | Both but > Basolateral | Type III | increased by ruxolitinib | [46,47] |
| Enterovirus 71 | No | NT | Fetal ** | Goblet | Apical | Apical | Type III | |||
| SARS-CoV-2 | No | Yes | D, I, C | Enterocyte | Apical 1000 fold > basolateral | NT | Type III | inhibited by imatinib, mycophenolic acid, quinacrine dihydrochloride ## | [36,48,49,50] | |
| Can epithelial cell responses to virus infection help explain pathogenesis? |
| Do goblet, enteroendocrine, or tuft cells have unique antiviral innate pathways to limit virus infection and/or replication? What mechanisms do viruses use to escape host innate responses? |
| Can novel viral receptor(s) be identified using polarized HIO monolayers in the transwell system? |
| Can viral interference, or bacterial–viral, and parasite–viral interactions, be modeled in HIOs? |
| Do all viruses use the same mechanisms to cross the intestinal epithelial barrier for systemic dissemination? |
| Can unique aspects of enteric virus replication, including dissecting slow viral kinetics, be elucidated in HIOs? |
| Will cell tropism for infection or epithelial cell responses differ in more advanced HIO cultures containing immune cells, innervation, or mesenchyme? |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crawford, S.E.; Ramani, S.; Blutt, S.E.; Estes, M.K. Organoids to Dissect Gastrointestinal Virus–Host Interactions: What Have We Learned? Viruses 2021, 13, 999. https://doi.org/10.3390/v13060999
Crawford SE, Ramani S, Blutt SE, Estes MK. Organoids to Dissect Gastrointestinal Virus–Host Interactions: What Have We Learned? Viruses. 2021; 13(6):999. https://doi.org/10.3390/v13060999
Chicago/Turabian StyleCrawford, Sue E., Sasirekha Ramani, Sarah E. Blutt, and Mary K. Estes. 2021. "Organoids to Dissect Gastrointestinal Virus–Host Interactions: What Have We Learned?" Viruses 13, no. 6: 999. https://doi.org/10.3390/v13060999
APA StyleCrawford, S. E., Ramani, S., Blutt, S. E., & Estes, M. K. (2021). Organoids to Dissect Gastrointestinal Virus–Host Interactions: What Have We Learned? Viruses, 13(6), 999. https://doi.org/10.3390/v13060999