
Establishment of a Three-Dimensional In Vitro Model of Equine Papillomavirus Type 2 Infection

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Material
2.2. Isolation of Keratinocytes
2.3. EcPV2- and ecGAPDH-PCR
2.4. Three-Dimensional (3D) Air-Liquid Interface Culture
2.5. EcPV2 RNA In Situ Hybridization
2.6. Immunohistochemistry for Epithelial, Mesenchymal, Cancer and Proliferation Markers
3. Results
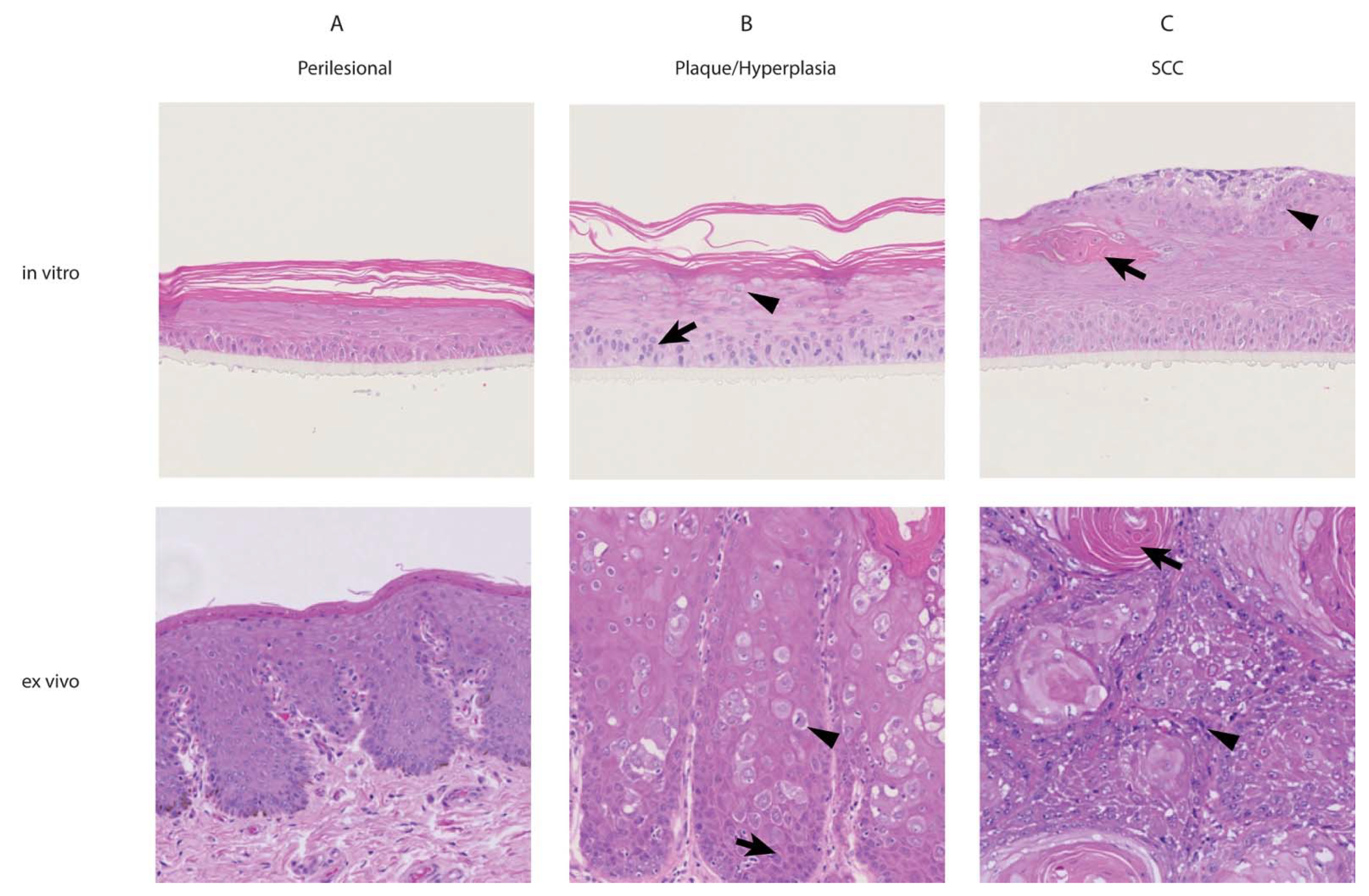
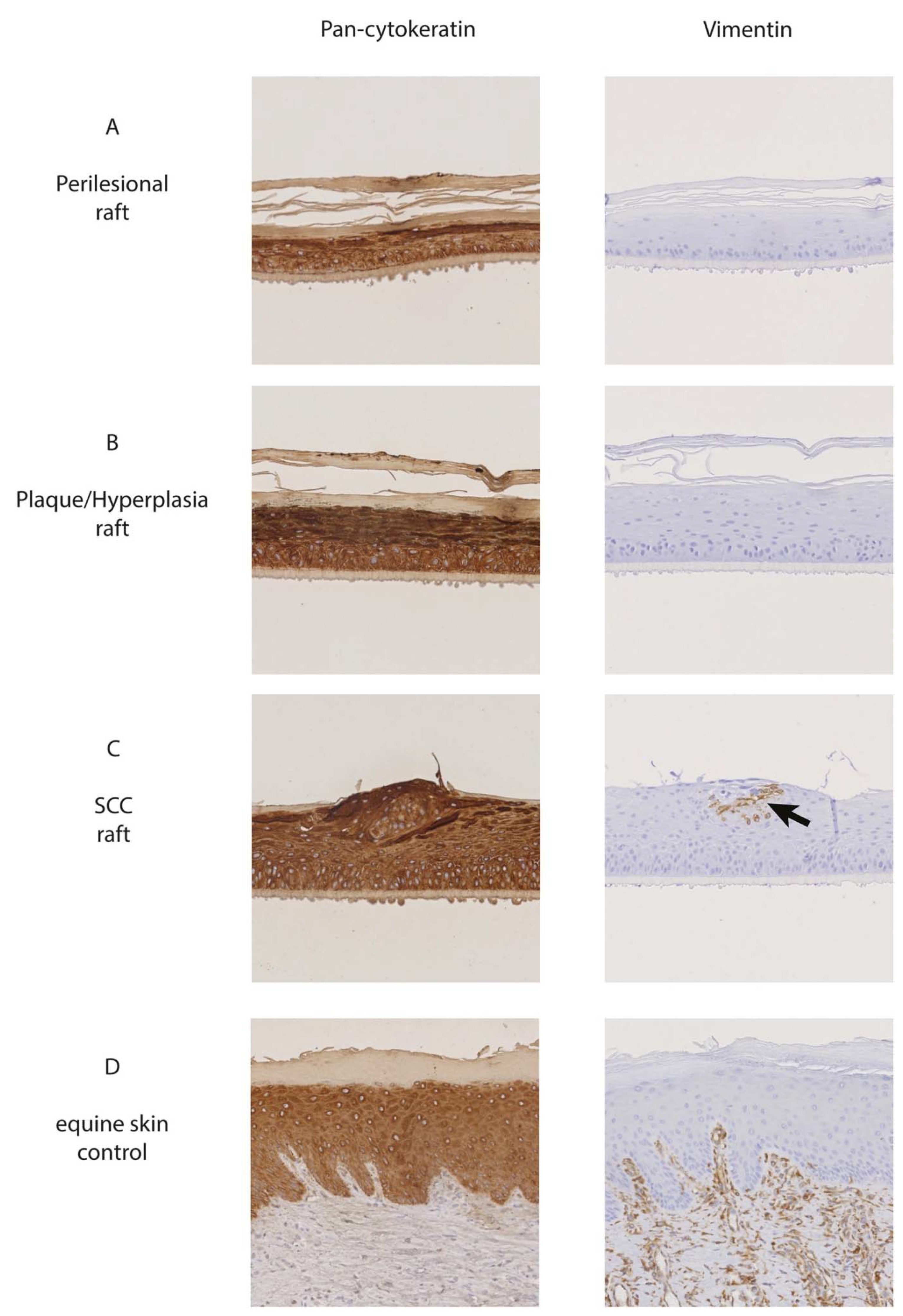
3.1. Sections from 3D Rafts and Corresponding Ex Vivo Tissue Material Exhibited Similar Histomorphological Features
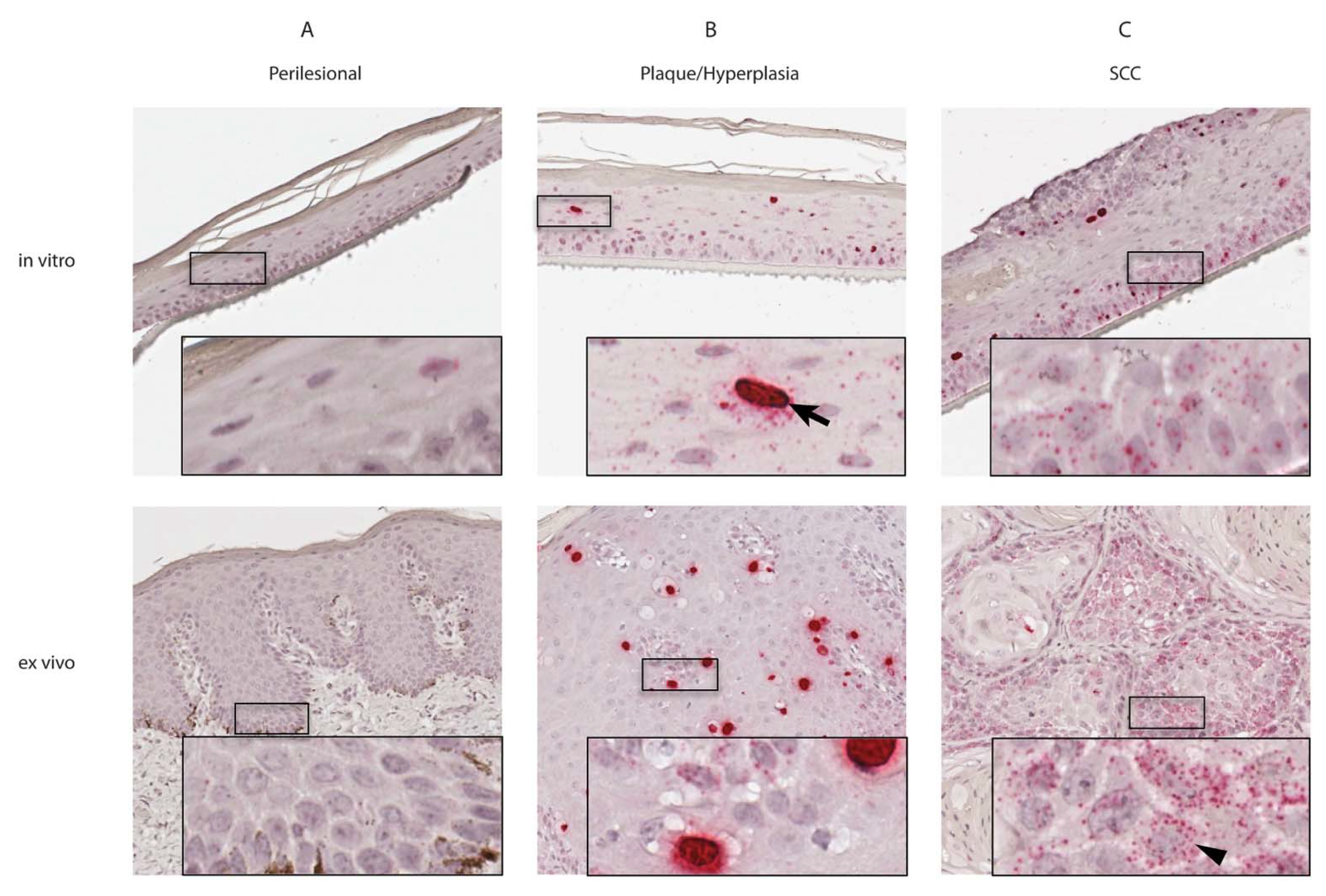
3.2. Equine Penile Lesions and Derived Raft Cultures Contained Multiple Copies of EcPV2 Nucleic Acids
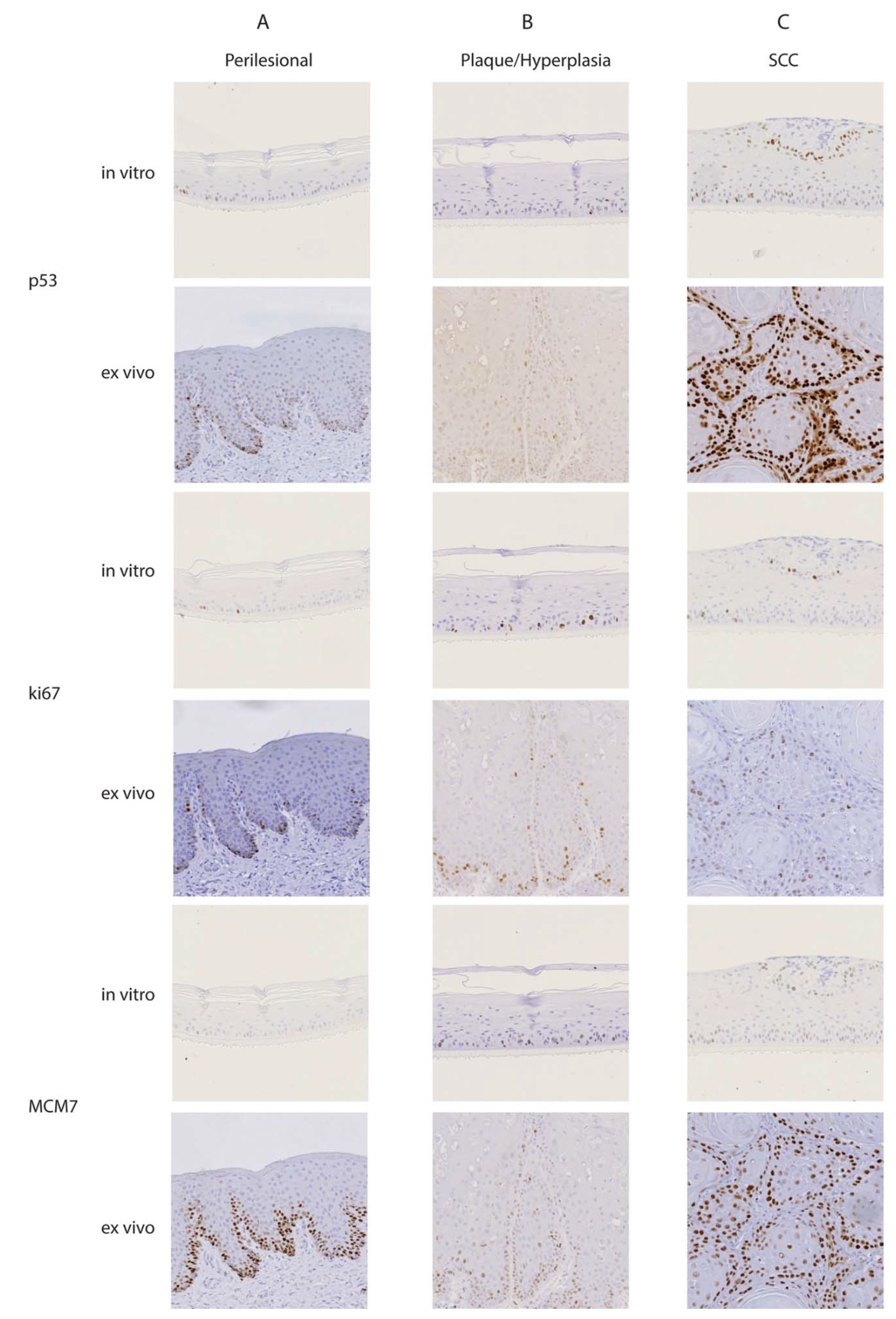
3.3. Lesional Raft Cultures Exhibited Enhanced Expression of Tumor and Proliferation Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Campo, M.S. Introduction. In Papillomavirus Research: From Natural History to Vaccines and Beyond, 1st ed.; Caister Academic Press: Norfolk, UK, 2006; pp. 1–2. [Google Scholar]
- Campo, M. Animal models of papillomavirus pathogenesis. Virus Res. 2002, 89, 249–261. [Google Scholar] [CrossRef]
- Doorbar, J. The papillomavirus life cycle. J. Clin. Virol. 2005, 32, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Day, P.M.; Lowy, D.R.; Schiller, J.T. Papillomaviruses infect cells via a clathrin-dependent pathway. Virology 2003, 307, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Giroglou, T.; Florin, L.; Schäfer, F.; Streeck, R.E.; Sapp, M. Human Papillomavirus Infection Requires Cell Surface Heparan Sulfate. J. Virol. 2001, 75, 1565–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spoden, G.; Freitag, K.; Husmann, M.; Boller, K.; Sapp, M.; Lambert, C.; Florin, L. Clathrin- and Caveolin-Independent Entry of Human Papillomavirus Type 16—Involvement of Tetraspanin-Enriched Microdomains (TEMs). PLoS ONE 2008, 3, e3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, S. Late events in the life cycle of human papillomaviruses. In Papillomavirus Research: From Natural History to Vaccines and Beyond, 1st ed.; Caister Academic Press: Norfolk, UK, 2006; pp. 193–212. [Google Scholar]
- Chambers, G.; Ellsmore, V.A.; O’Brien, P.M.; Reid, S.; Love, S.; Campo, M.S.; Nasir, L. Association of bovine papillomavirus with the equine sarcoid. J. Gen. Virol. 2003, 84, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Doorbar, J.; Quint, W.; Banks, L.; Bravo, I.G.; Stoler, M.; Broker, T.R.; Stanley, M.A. The Biology and Life-Cycle of Human Papillomaviruses. Vaccine 2012, 30, F55–F70. [Google Scholar] [CrossRef]
- Van Doorslaer, K.; Li, Z.; Xirasagar, S.; Maes, P.; Kaminsky, D.; Liou, D.; Sun, Q.; Kaur, R.; Huyen, Y.; McBride, A.A. The Papillomavirus Episteme: A major update to the papillomavirus sequence database. Nucleic Acids Res. 2017, 45, D499–D506. [Google Scholar] [CrossRef]
- Dayyani, F.; Etzel, C.J.; Liu, M.; Ho, C.-H.; Lippman, S.M.; Tsao, A.S. Meta-analysis of the impact of human papillomavirus (HPV) on cancer risk and overall survival in head and neck squamous cell carcinomas (HNSCC). Head Neck Oncol. 2010, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.W.; Miller, W.H., Jr. Squamous cell carcinoma. In Equine Dermatology, 1st ed.; Saunders Elsevier: St. Louis, MO, USA, 2003; pp. 707–712. [Google Scholar]
- Knottenbelt, D.C.; Patterson-Kane, J.C.; Snalune, K.L. Squamous Cell Carcinoma; Elsevier BV: Amsterdam, The Netherlands, 2015; pp. 220–236. [Google Scholar]
- Bogaert, L.; Willemsen, A.; Vanderstraeten, E.; Bracho, M.A.; De Baere, C.; Bravo, I.G.; Martens, A. EcPV2 DNA in equine genital squamous cell carcinomas and normal genital mucosa. Vet. Microbiol. 2012, 158, 33–41. [Google Scholar] [CrossRef]
- Knight, C.G.; Munday, J.S.; Peters, J.; Dunowska, M. Equine Penile Squamous Cell Carcinomas Are Associated With the Presence of Equine Papillomavirus Type 2 DNA Sequences. Vet. Pathol. 2011, 48, 1190–1194. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.E.; Tobler, K.; Lehner, A.; Grest, P.; Welle, M.M.; Schwarzwald, C.C.; Favrot, C. EcPV2 DNA in equine papillomas and in situ and invasive squamous cell carcinomas supports papillomavirus etiology. Vet. Pathol. 2013, 50, 686–692. [Google Scholar] [CrossRef]
- Scase, T.; Brandt, S.; Kainzbauer, C.; Sykora, S.; Bijmholt, S.; Hughes, K.; Sharpe, S.; Foote, A. Equus caballus papillomavirus-2 (EcPV-2): An infectious cause for equine genital cancer? Equine Vet. J. 2010, 42, 738–745. [Google Scholar] [CrossRef]
- Sykora, S.; Brandt, S. Papillomavirus infection and squamous cell carcinoma in horses. Vet. J. 2017, 223, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Sykora, S.; Jindra, C.; Hofer, M.; Steinborn, R.; Brandt, S. Equine papillomavirus type 2: An equine equivalent to human papillomavirus 16? Vet. J. 2017, 225, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Ramsauer, A.S.; Kubacki, J.; Favrot, C.; Ackermann, M.; Fraefel, C.; Tobler, K. RNA-seq analysis in equine papillomavirus type 2-positive carcinomas identifies affected pathways and potential cancer markers as well as viral gene expression and splicing events. J. Gen. Virol. 2019, 100, 985–998. [Google Scholar] [CrossRef]
- Kainzbauer, C.; Rushton, J.; Tober, R.; Scase, T.; Nell, B.; Sýkora, S.; Brandt, S. Bovine papillomavirus type 1 and Equus caballus papillomavirus 2 in equine squamous cell carcinoma of the head and neck in a Connemara mare. Equine Vet. J. 2011, 44, 112–115. [Google Scholar] [CrossRef]
- Alloway, E.; Linder, K.; May, S.; Rose, T.; DeLay, J.; Bender, S.; Tucker, A.; Luff, J. A Subset of Equine Gastric Squamous Cell Carcinomas Is Associated with Equus Caballus Papillomavirus–2 Infection. Vet. Pathol. 2020, 57, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.G.; Dunowska, M.; Munday, J.S.; Peters-Kennedy, J.; Rosa, B.V.; Rosa, B. Comparison of the levels of Equus caballus papillomavirus type 2 (EcPV-2) DNA in equine squamous cell carcinomas and non-cancerous tissues using quantitative PCR. Vet. Microbiol. 2013, 166, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Laimins, L.A. Differentiation of HPV-Containing Cells Using Organotypic “Raft” Culture or Methylcellulose. In Human Papillomaviruses; Davy, C.J.D., Ed.; Humana Press Inc: Totowa, NJ, USA, 2005; pp. 157–169. [Google Scholar]
- Liu, X.; Krawczyk, E.; Suprynowicz, F.A.; Palechor-Ceron, N.; Yuan, H.; Dakic, A.; Simic, V.; Zheng, Y.-L.; Sripadhan, P.; Chen, C.; et al. Conditional reprogramming and long-term expansion of normal and tumor cells from human biospecimens. Nat. Protoc. 2017, 12, 439–451. [Google Scholar] [CrossRef]
- Alkhilaiwi, F.; Wang, L.; Zhou, D.; Raudsepp, T.; Ghosh, S.; Paul, S.; Palechor-Ceron, N.; Brandt, S.; Luff, J.; Liu, X.; et al. Long-term expansion of primary equine keratinocytes that maintain the ability to differentiate into stratified epidermis. Stem Cell Res. Ther. 2018, 9, 181. [Google Scholar] [CrossRef]
- Liu, X.; Ory, V.; Chapman, S.; Yuan, H.; Albanese, C.; Kallakury, B.; Timofeeva, O.A.; Nealon, C.; Dakic, A.; Simic, V.; et al. ROCK inhibitor and feeder cells induce the conditional reprogramming of epithelial cells. Am. J. Pathol. 2012, 180, 599–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsauer, A.S.; Wachoski-Dark, G.L.; Fraefel, C.; Tobler, K.; Brandt, S.; Knight, C.G.; Favrot, C.; Grest, P. Paving the way for more precise diagnosis of EcPV2-associated equine penile lesions. BMC Vet. Res. 2019, 15, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, K.W.; Affolter, V.K.; Gaynor, A.M.; Dela Cruz, F.N., Jr.; Pesavento, P.A. Equine Genital Squamous Cell Carcinoma: In Situ Hybridization Identifies a Distinct Subset Containing Equus caballus Papillomavirus 2. Vet. Pathol. 2015, 52, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Matoltsy, A.G. Keratinization. J. Investig. Dermatol. 1976, 67, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Guiraud, B.; Hernandez-Pigeon, H.; Ceruti, I.; Mas, S.; Palvadeau, Y.; Saint-Martory, C.; Castex-Rizzi, N.; Duplan, H.; Bessou-Touya, S. Characterization of a human epidermis model reconstructed from hair follicle keratinocytes and comparison with two commercially models and native skin. Int. J. Cosmet. Sci. 2014, 36, 485–493. [Google Scholar] [CrossRef]
- Rigon, R.B.; Kaessmeyer, S.; Wolff, C.; Hausmann, C.; Zhang, N.; Sochorova, M.; Kováčik, A.; Haag, R.; Vávrová, K.; Ulrich, M.; et al. Ultrastructural and Molecular Analysis of Ribose-Induced Glycated Reconstructed Human Skin. Int. J. Mol. Sci. 2018, 19, 3521. [Google Scholar] [CrossRef] [Green Version]
- Cerrato, S.; Ramió-Lluch, L.; Brazis, P.; Rabanal, R.M.; Fondevila, D.; Puigdemont, A. Development and characterization of an equine skin-equivalent model. Vet. Dermatol. 2014, 25, 475-e77. [Google Scholar] [CrossRef]
- Wechsler, E.I.; Wang, Q.; Roberts, I.; Pagliarulo, E.; Jackson, D.; Untersperger, C.; Coleman, N.; Griffin, H.; Doorbar, J. Reconstruction of human papillomavirus type 16-mediated early-stage neoplasia implicates E6/E7 deregulation and the loss of contact inhibition in neoplastic progression. J. Virol. 2012, 86, 6358–6364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anacker, D.; Moody, C. Generation of Organotypic Raft Cultures from Primary Human Keratinocytes. J. Vis. Exp. 2012, 2012, e3668. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.; Hillpot, E.; Mondal, S.; Khurana, K.K.; Woodworth, C.D. HPV16-Immortalized Cells from Human Transformation Zone and Endocervix are More Dysplastic than Ectocervical Cells in Organotypic Culture. Sci. Rep. 2018, 8, 15402. [Google Scholar] [CrossRef]
- Srivastava, K.; Pickard, A.; McDade, S.; McCance, D.J. p63 drives invasion in keratinocytes expressing HPV16 E6/E7 genes through regulation of Src-FAK signalling. Oncotarget 2017, 8, 16202–16219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.F.; Peng, Z.; Clark, K.M.; Adamson, C.S.-C.; Ma, X.-J.; Wu, X.; Wang, H.; Luo, Y.; Cooper, K. HPV E6/E7 RNA In Situ Hybridization Signal Patterns as Biomarkers of Three-Tier Cervical Intraepithelial Neoplasia Grade. PLoS ONE 2014, 9, e91142. [Google Scholar] [CrossRef] [Green Version]
- Johansson, C.; Schwartz, S. Regulation of human papillomavirus gene expression by splicing and polyadenylation. Nat. Rev. Genet. 2013, 11, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Gilles, C.; Polette, M.; Piette, J.; Delvigne, A.-C.; Thompson, E.W.; Foidart, J.-M.; Birembaut, P. Vimentin Expression in Cervical Carcinomas: Association with Invasive and Migratory Potential. J. Pathol. 1996, 180, 175–180. [Google Scholar] [CrossRef]
- Franke, W.W.; Schmid, E.; Winter, S.; Osborn, M.; Weber, K. Widespread occurrence of intermediate-sized filaments of the vimentin-type in cultured cells from diverse vertebrates. Exp. Cell Res. 1979, 123, 25–46. [Google Scholar] [CrossRef]
- Virtanen, I.; Lehto, V.; Lehtonen, E.; Vartio, T.; Stenman, S.; Kurki, P.; Wager, O.; Small, J.; Dahl, D.; Badley, R. Expression of intermediate filaments in cultured cells. J. Cell Sci. 1981, 50, 45–63. [Google Scholar] [CrossRef]
- Steenbergen, R.D.; Parker, J.N.; Isern, S.; Snijders, P.J.; Walboomers, J.M.; Meijer, C.J.; Broker, T.R.; Chow, L.T. Viral E6-E7 transcription in the basal layer of organotypic cultures without apparent p21cip1 protein precedes immortalization of human papillomavirus type 16- and 18-transfected human keratinocytes. J. Virol. 1998, 72, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.C.; Gonçalves, A.; Cobucci, R.N.; Mendonça, R.C.; Lima, P.H.; Cavalcanti, G. Immunohistochemical expression of p16, Ki-67 and p53 in cervical lesions—A systematic review. Pathol. Res. Pract. 2017, 213, 723–729. [Google Scholar] [CrossRef]
- Gariglio, P.; Organista-Nava, J.; Alvarez-Rios, E. Role of HR-HPVs E6 and E7 Oncoproteins in Cervical Carcinogenesis. J. Mol. Genet. Med. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kho, E.-Y.; Wang, H.-K.; Banerjee, N.S.; Broker, T.R.; Chow, L.T. HPV-18 E6 mutants reveal p53 modulation of viral DNA amplification in organotypic cultures. Proc. Natl. Acad. Sci. USA 2013, 110, 7542–7549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Zapien, D.; Ruiz, F.X.; Poirson, J.; Mitschler, A.; Ramirez, J.; Forster, A.; Cousido-Siah, A.; Masson, M.; Pol, S.V.; Podjarny, A. Structure of the E6/E6AP/p53 complex required for HPV-mediated degradation of p53. Nature 2016, 529, 541–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brimer, N.; Drews, C.M.; Pol, S.B.V. Association of papillomavirus E6 proteins with either MAML1 or E6AP clusters E6 proteins by structure, function, and evolutionary relatedness. PLoS Pathog. 2017, 13, e1006781. [Google Scholar] [CrossRef]
- Murnyák, B.; Hortobágyi, T. Immunohistochemical correlates of TP53 somatic mutations in cancer. Oncotarget 2016, 7, 64910–64920. [Google Scholar] [CrossRef] [Green Version]
- Ando, K.; Oki, E.; Saeki, H.; Yan, Z.; Tsuda, Y.; Hidaka, G.; Kasagi, Y.; Otsu, H.; Kawano, H.; Kitao, H.; et al. Discrimination of p53 immunohistochemistry-positive tumors by its staining pattern in gastric cancer. Cancer Med. 2014, 4, 75–83. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Use | Retrieval | 2nd Antibody | Chromogen | Control |
|---|---|---|---|---|---|
| Monoclonal mouse vimentin (M7020, Dako, Agilent, Santa Clara, CA, USA) | 1:300, 30 min | 98 °C, pH 6 | MACH4 (BioCare Medical, Pacheco, CA, USA) | DAB | Equine skin |
| Monoclonal mouse PCK-26 (NB120-6401; Novus Biologicals LLC, Boulder, CO, USA) | 1:500, 60 min | 98 °C, pH 9, blocking serum | EnVision Mouse (K4001, Dako, Agilent, Santa Clara, CA, USA) | DAB | Equine skin |
| Monoclonal mouse anti-p53 (clone DO1; Santa CruzBiotechnology Inc, Santa Cruz, CA, USA) | 1:100, 60 min | 98 °C; pH 9 | EnVision Mouse (K4001; Dako, Agilent, Santa Clara, CA, USA) | DAB | Equine squmamous cell carcinoma |
| Monoclonal mouse anti-Ki67 (clone MIB-1; Dako, Agilent, Santa Clara, CA, USA) | 1:50, 60 min | 98 °C; pH 9 | REAL Kit (K5001; Dako, Agilent, Santa Clara, CA, USA) | AEC | Equine skin |
| Monoclonal mouse anti-MCM7 (clone DCS-141.2; Santa CruzBiotechnology Inc, Santa Cruz, CA, USA) | 1:400, 60 min | 98 °C; pH 9 | EnVision Mouse (K4001, Dako, Agilent, Santa Clara, CA, USA) | DAB | Equine lymph node |
| Sections from | GS | DNS | Figure |
|---|---|---|---|
| Perilesional skin-derived rafts | +/− | − | Figure 2A |
| Perilesional skin | +/− | − | |
| Plaque-derived rafts | + | + | Figure 2B |
| Plaque tissue | + | ++ | |
| SCC-derived rafts | ++ | + | Figure 2C |
| SCC tissue | ++ | +/− |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramsauer, A.S.; Wachoski-Dark, G.L.; Fraefel, C.; Ackermann, M.; Brandt, S.; Grest, P.; Knight, C.G.; Favrot, C.; Tobler, K. Establishment of a Three-Dimensional In Vitro Model of Equine Papillomavirus Type 2 Infection. Viruses 2021, 13, 1404. https://doi.org/10.3390/v13071404
Ramsauer AS, Wachoski-Dark GL, Fraefel C, Ackermann M, Brandt S, Grest P, Knight CG, Favrot C, Tobler K. Establishment of a Three-Dimensional In Vitro Model of Equine Papillomavirus Type 2 Infection. Viruses. 2021; 13(7):1404. https://doi.org/10.3390/v13071404
Chicago/Turabian StyleRamsauer, Anna Sophie, Garrett Louis Wachoski-Dark, Cornel Fraefel, Mathias Ackermann, Sabine Brandt, Paula Grest, Cameron Greig Knight, Claude Favrot, and Kurt Tobler. 2021. "Establishment of a Three-Dimensional In Vitro Model of Equine Papillomavirus Type 2 Infection" Viruses 13, no. 7: 1404. https://doi.org/10.3390/v13071404
APA StyleRamsauer, A. S., Wachoski-Dark, G. L., Fraefel, C., Ackermann, M., Brandt, S., Grest, P., Knight, C. G., Favrot, C., & Tobler, K. (2021). Establishment of a Three-Dimensional In Vitro Model of Equine Papillomavirus Type 2 Infection. Viruses, 13(7), 1404. https://doi.org/10.3390/v13071404