
HTLV-2 Encoded Antisense Protein APH-2 Suppresses HIV-1 Replication


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Plasmids
2.2. Virus Release and Infectivity Assays
2.3. APH-2 and Virus-Like Particle Release
2.4. Cellular Gag mRNA Quantification
2.5. Virus Release Efficiency
2.6. Assays for Detection of Role of APH-2 on HIV-1 Transcription
2.7. Immunoprecipitation
3. Results
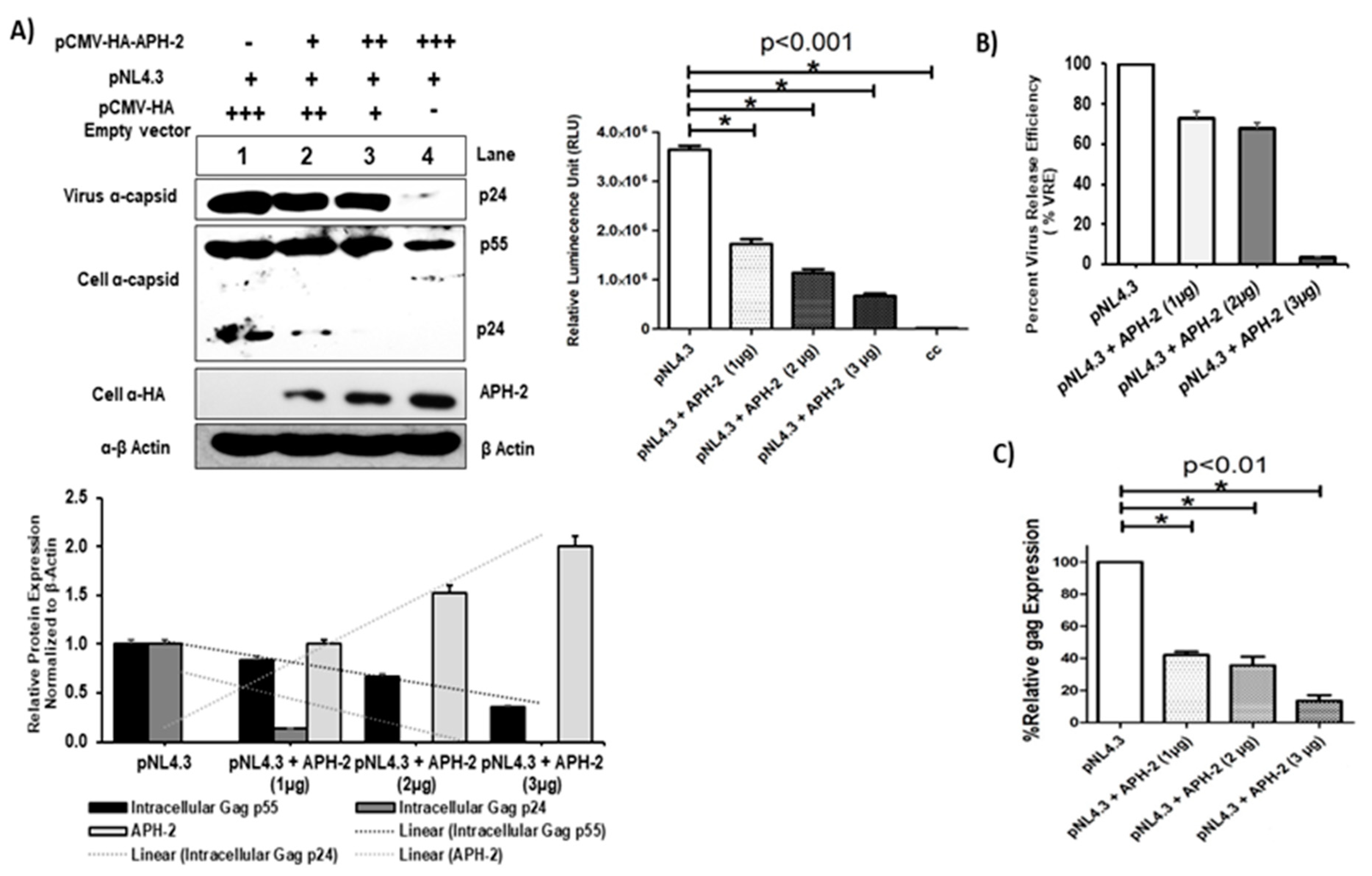
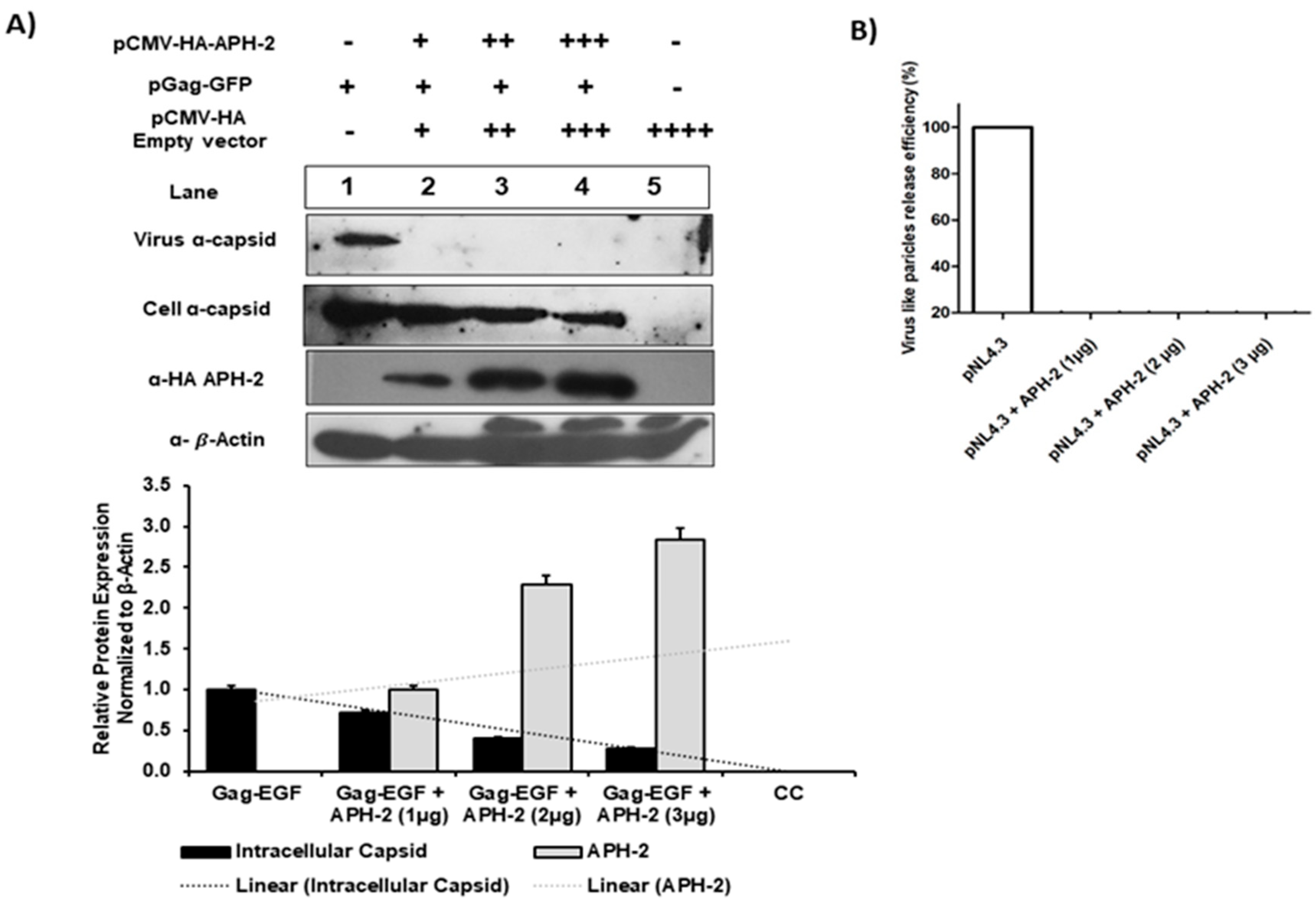
3.1. Dose-Dependent Effect of HTLV-2 APH-2 Expression on HIV-1 Release and Infectivity
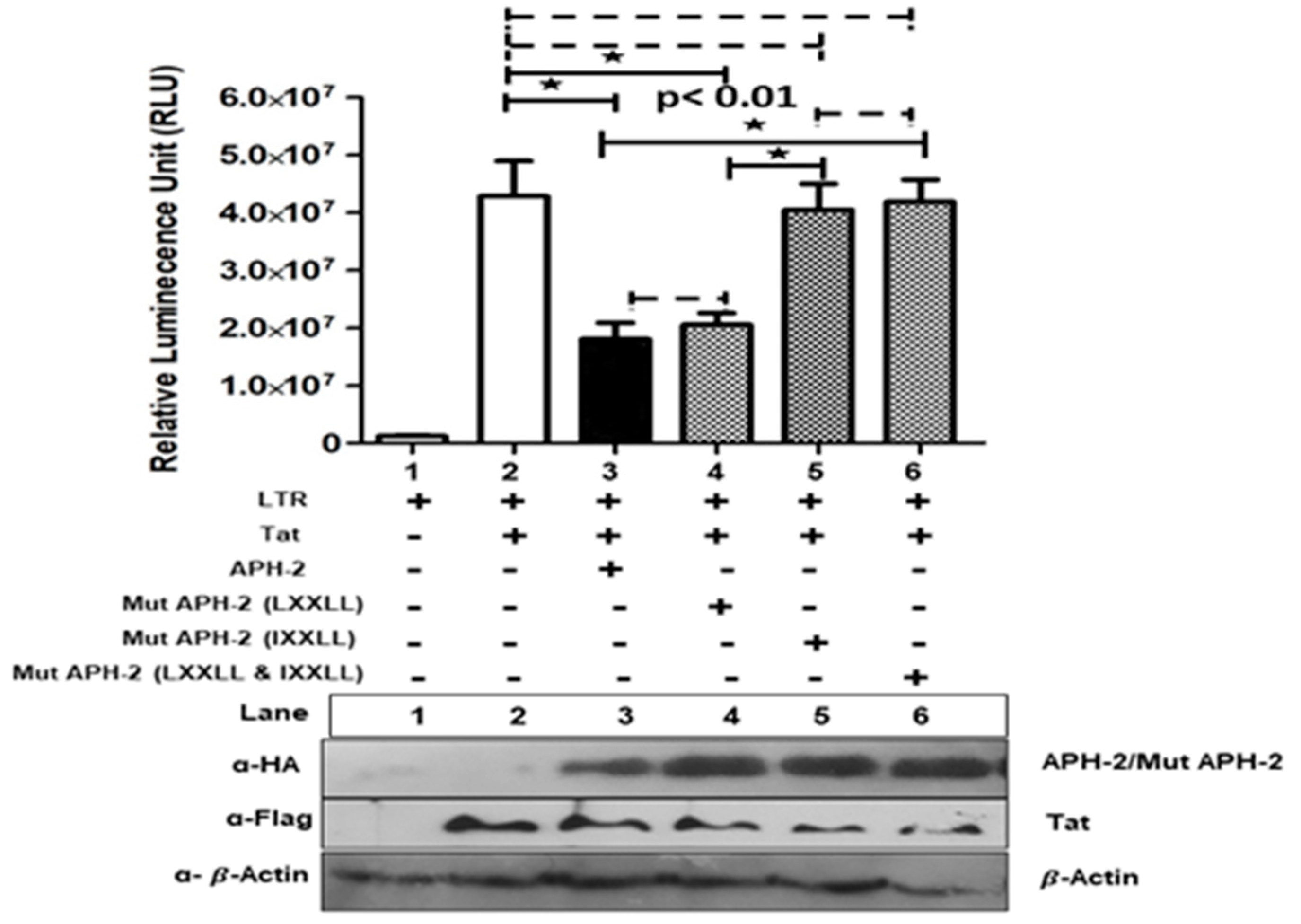
3.2. Role of APH-2 in HIV-1 Transcription
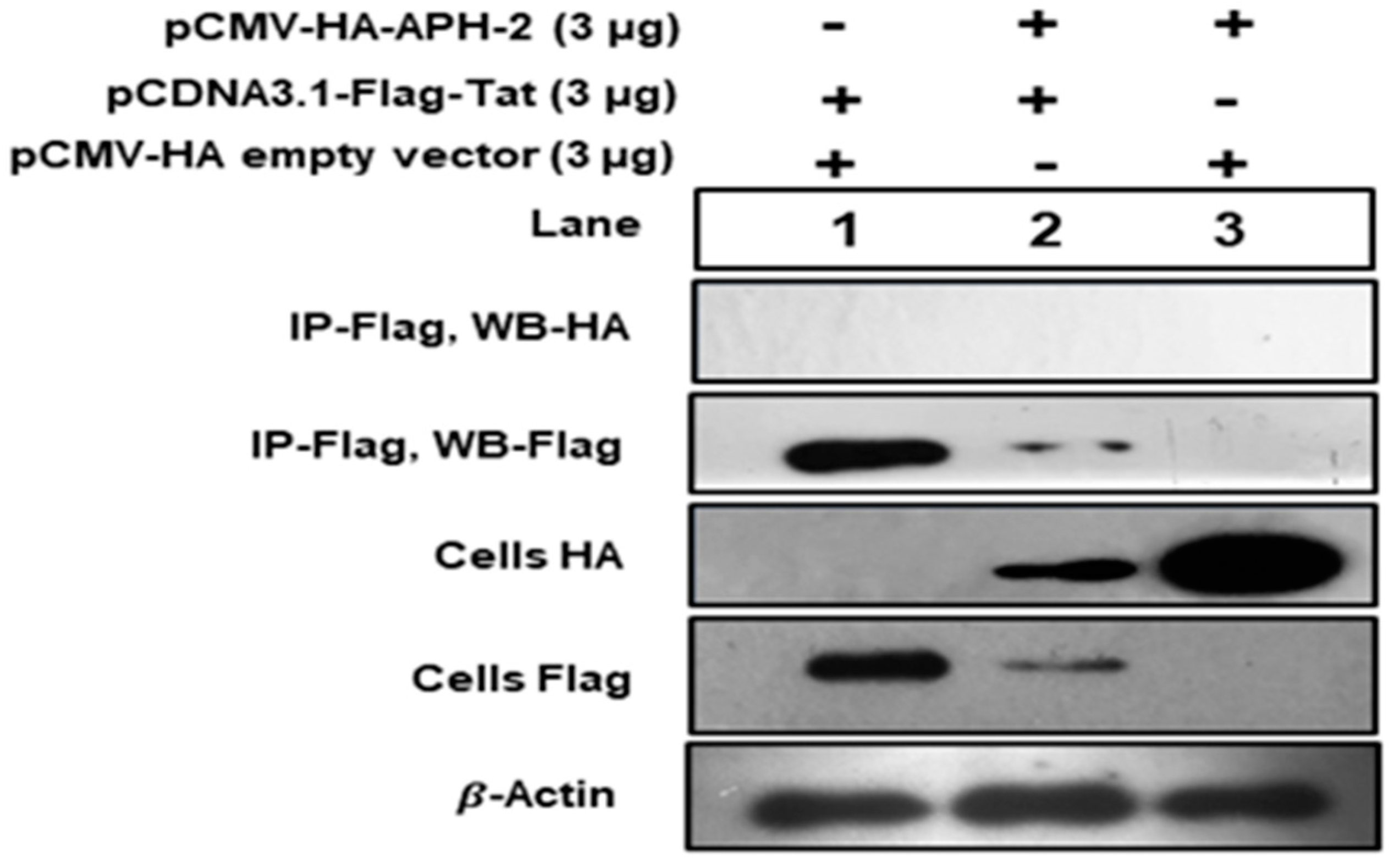
3.3. Association of Tat and APH-2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tagaya, Y.; Matsuoka, M.; Gallo, R. 40 years of the human T-cell leukemia virus: Past, present, and future. F1000Research 2019, 8, 228. [Google Scholar] [CrossRef] [Green Version]
- Bartman, M.T.; Kaidarova, Z.; Hirschkorn, D.; Sacher, R.A.; Fridey, J.; Garratty, G.; Gibble, J.; Smith, J.W.; Newman, B.; Yeo, A.E.; et al. Long-term increases in lymphocytes and platelets in human T-lymphotropic virus type II infection. Blood 2008, 112, 3995–4002. [Google Scholar] [CrossRef] [Green Version]
- Orland, J.R.; Wang, B.; Wright, D.J.; Nass, C.C.; Garratty, G.; Smith, J.W.; Newman, B.; Smith, D.M.; Murphy, E.L. Increased mortality associated with HTLV-II infection in blood donors: A prospective cohort study. Retrovirology 2004, 1, 4. [Google Scholar] [CrossRef] [Green Version]
- Gaudray, G.; Gachon, F.; Basbous, J.; Biard-Piechaczyk, M.; Devaux, C.; Mesnard, J.-M. The Complementary Strand of the Human T-Cell Leukemia Virus Type 1 RNA Genome Encodes a bZIP Transcription Factor That Down-Regulates Viral Transcription. J. Virol. 2002, 76, 12813–12822. [Google Scholar] [CrossRef] [Green Version]
- Halin, M.; Douceron, E.; Clerc, I.; Journo, C.; Ko, N.L.; Landry, S.; Murphy, E.L.; Gessain, A.; Lemasson, I.; Mesnard, J.-M.; et al. Human T-cell leukemia virus type 2 produces a spliced antisense transcript encoding a protein that lacks a classic bZIP domain but still inhibits Tax2-mediated transcription. Blood 2009, 114, 2427–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Kannian, P.; Dissinger, N.; Haines, R.; Niewiesk, S.; Green, P.L. Human T-Cell Leukemia Virus Type 2 Antisense Viral Protein 2 Is Dispensable forIn VitroImmortalization but Functions To Repress Early Virus Replication In Vivo. J. Virol. 2012, 86, 8412–8421. [Google Scholar] [CrossRef] [Green Version]
- Douceron, E.; Kaidarova, Z.; Miyazato, P.; Matsuoka, M.; Murphy, E.L.; Mahieux, R. HTLV-2 APH-2 Expression Is Correlated With Proviral Load but APH-2 Does Not Promote Lymphocytosis. J. Infect. Dis. 2011, 205, 82–86. [Google Scholar] [CrossRef]
- Lemasson, I.; Lewis, M.R.; Polakowski, N.; Hivin, P.; Cavanagh, M.-H.; Thébault, S.; Barbeau, B.; Nyborg, J.K.; Mesnard, J.-M. Human T-Cell Leukemia Virus Type 1 (HTLV-1) bZIP Protein Interacts with the Cellular Transcription Factor CREB To Inhibit HTLV-1 Transcription. J. Virol. 2007, 81, 1543–1553. [Google Scholar] [CrossRef] [Green Version]
- Satou, Y.; Yasunaga, J.-I.; Yoshida, M.; Matsuoka, M. HTLV-I basic leucine zipper factor gene mRNA supports proliferation of adult T cell leukemia cells. Proc. Natl. Acad. Sci. USA 2006, 103, 720–725. [Google Scholar] [CrossRef] [Green Version]
- Fochi, S.; Mutascio, S.; Bertazzoni, U.; Zipeto, D.; Romanelli, M.G. HTLV Deregulation of the NF-κB Pathway: An Update on Tax and Antisense Proteins Role. Front. Microbiol. 2018, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Fochi, S.; Bergamo, E.; Serena, M.; Mutascio, S.; Journo, C.; Mahieux, R.; Ciminale, V.; Bertazzoni, U.; Zipeto, D.; Romanelli, M.G. TRAF3 Is Required for NF-κB Pathway Activation Mediated by HTLV Tax Proteins. Front. Microbiol. 2019, 10, 1302. [Google Scholar] [CrossRef] [Green Version]
- Panfil, A.R.; Dissinger, N.J.; Howard, C.; Murphy, B.M.; Landes, K.; Fernandez, S.A.; Green, P.L. Functional Comparison of HBZ and the Related APH-2 Protein Provides Insight into Human T-Cell Leukemia Virus Type 1 Pathogenesis. J. Virol. 2016, 90, 3760–3772. [Google Scholar] [CrossRef] [Green Version]
- Barrios, C.S.; Abuerreish, M.; Lairmore, M.D.; Castillo, L.; Giam, C.-Z.; Beilke, M.A. Recombinant Human T-Cell Leukemia Virus Types 1 and 2 Tax Proteins Induce High Levels of CC-Chemokines and Downregulate CCR5 in Human Peripheral Blood Mononuclear Cells. Viral Immunol. 2011, 24, 429–439. [Google Scholar] [CrossRef]
- Casoli, C.; Pilotti, E.; Bertazzoni, U. Molecular and cellular interactions of HIV-1/HTLV coinfection and impact on AIDS progression. Aids Rev. 2007, 9, 140–149. [Google Scholar]
- Casoli, C.; Vicenzi, E.; Cimarelli, A.; Magnani, G.; Ciancianaini, P.; Cattaneo, E.; Dall’Aglio, P.; Poli, G.; Bertazzoni, U. HTLV-II down-regulates HIV-1 replication in IL-2-stimulated primary PBMC of coinfected individuals through expression of MIP-1alpha. Blood 2000, 95, 2760–2769. [Google Scholar] [CrossRef]
- Gudo, E.S.; Bhatt, N.B.; Bila, D.R.; Abreu, C.M.; Tanuri, A.; Savino, W.; Silva-Barbosa, S.D.; Jani, I.V. Co-infection by human immunodeficiency virus type 1 (HIV-1) and human T cell leukemia virus type 1 (HTLV-1): Does immune activation lead to a faster progression to AIDS? BMC Infect. Dis. 2009, 9, 211–213. [Google Scholar] [CrossRef] [Green Version]
- Pedroso, C.; Netto, E.; Weyll, N.; Brites, C. Coinfection by HIV-1 and Human Lymphotropic Virus Type 1 in Brazilian Children Is Strongly Associated With a Shorter Survival Time. JAIDS J. Acquir. Immune Defic. Syndr. 2011, 57, S208–S211. [Google Scholar] [CrossRef]
- Pilotti, E.; Elviri, L.; Vicenzi, E.; Bertazzoni, U.; Re, M.C.; Allibardi, S.; Poli, G.; Casoli, C. Postgenomic up-regulation of CCL3L1 expression in HTLV-2–infected persons curtails HIV-1 replication. Blood 2006, 109, 1850–1856. [Google Scholar] [CrossRef] [Green Version]
- Bogerd, H.P.; Huckaby, G.L.; Ahmed, Y.F.; Hanly, S.M.; Greene, W.C. The type I human T-cell leukemia virus (HTLV-I) Rex trans-activator binds directly to the HTLV-I Rex and the type 1 human immunodeficiency virus Rev RNA response elements. Proc. Natl. Acad. Sci. USA 1991, 88, 5704–5708. [Google Scholar] [CrossRef] [Green Version]
- Lewis, M.J.; Gautier, V.W.; Wang, X.-P.; Kaplan, M.H.; Hall, W.W. Spontaneous Production of C-C Chemokines by Individuals Infected with Human T Lymphotropic Virus Type II (HTLV-II) Alone and HTLV-II/HIV-1 Coinfected Individuals. J. Immunol. 2000, 165, 4127–4132. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, Y.; Kubota, R.; Tara, M.; Izumo, S.; Osame, M. Existence of escape mutant in HTLV-I tax during the development of adult T-cell leukemia. Blood 2001, 97, 987–993. [Google Scholar] [CrossRef] [Green Version]
- Moriuchi, H.; Moriuchi, M.; Fauci, A.S. Factors Secreted by Human T Lymphotropic Virus Type I (HTLV-I)–infected Cells Can Enhance or Inhibit Replication of HIV-1 in HTLV-I–uninfected Cells: Implications for In Vivo Coinfection with HTLV-I and HIV-1. J. Exp. Med. 1998, 187, 1689–1697. [Google Scholar] [CrossRef] [Green Version]
- Torresilla, C.; Carmo, S.D.; Larocque, É.; Douceron, E.; Mesnard, J.-M.; Mahieux, R.; Barbeau, B. The antisense protein of HTLV-2 positively modulates HIV-1 replication. Retrovirology 2014, 11, P118. [Google Scholar] [CrossRef] [Green Version]
- Marzio, G.; Tyagi, M.; Gutierrez, M.I.; Giacca, M. HIV-1 Tat transactivator recruits p300 and CREB-binding protein histone acetyltransferases to the viral promoter. Proc. Natl. Acad. Sci. USA 1998, 95, 13519–13524. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, H.S.; Pauza, C.D.; Bukrinsky, M.; Zhao, R.Y. Roles of HIV-1 auxiliary proteins in viral pathogenesis and host-pathogen interactions. Cell Res. 2005, 15, 923–934. [Google Scholar] [CrossRef]
- Ross, S.G.H.L.; Gartner, S.; McArthur, J.C.; Corboy, J.R.; McAllister, J.J.; Millhouse, S.; Wigdahl, B.; Ross, H.L. HIV-1 LTR C/EBP binding site sequence configurations preferentially encountered in brain lead to enhanced C/EBP factor binding and increased LTR-specific activity. J. NeuroVirol. 2001, 7, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Martini, F.; Arone, C.; Hasset, A.; Hall, W.W.; Sheehy, N. The ESCRT-0 Protein HRS Interacts with the Human T Cell Leukemia Virus Type 2 Antisense Protein APH-2 and Suppresses Viral Replication. J. Virol. 2019, 94. [Google Scholar] [CrossRef]
- Martin-Serrano, J.; Zang, T.; Bieniasz, P.D. HIV-1 and Ebola virus encode small peptide motifs that recruit Tsg101 to sites of particle assembly to facilitate egress. Nat. Med. 2001, 7, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- VerPlank, L.; Bouamr, F.; LaGrassa, T.J.; Agresta, B.; Kikonyogo, A.; Leis, J.; Carter, C.A. Tsg101, a homologue of ubiquitin-conjugating (E2) enzymes, binds the L domain in HIV type 1 Pr55Gag. Proc. Natl. Acad. Sci. USA 2001, 98, 7724–7729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrus, J.E.; von Schwedler, U.K.; Pornillos, O.W.; Morham, S.G.; Zavitz, K.H.; Wang, H.E.; Wettstein, D.A.; Stray, K.M.; Côté, M.; Rich, R.L.; et al. Tsg101 and the Vacuolar Protein Sorting Pathway Are Essential for HIV-1 Budding. Cell 2001, 107, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.; Hall, W.W.; Ratner, L.; Sheehy, N. Novel interactions between the HTLV antisense proteins HBZ and APH-2 and the NFAR protein family: Implications for the HTLV lifecycles. Virology 2016, 494, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Marban, C.; McCabe, Á.; Bukong, T.N.; Hall, W.W.; Sheehy, N. Interplay between the HTLV-2 Tax and APH-2 proteins in the regulation of the AP-1 pathway. Retrovirology 2012, 9, 98. [Google Scholar] [CrossRef]
- Isono, O.; Ohshima, T.; Saeki, Y.; Matsumoto, J.; Hijikata, M.; Tanaka, K.; Shimotohno, K. Human T-cell Leukemia Virus Type 1 HBZ Protein Bypasses the Targeting Function of Ubiquitination. J. Biol. Chem. 2008, 283, 34273–34282. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Londhe, R.; Kulkarni, S. HTLV-2 Encoded Antisense Protein APH-2 Suppresses HIV-1 Replication. Viruses 2021, 13, 1432. https://doi.org/10.3390/v13081432
Londhe R, Kulkarni S. HTLV-2 Encoded Antisense Protein APH-2 Suppresses HIV-1 Replication. Viruses. 2021; 13(8):1432. https://doi.org/10.3390/v13081432
Chicago/Turabian StyleLondhe, Rajkumar, and Smita Kulkarni. 2021. "HTLV-2 Encoded Antisense Protein APH-2 Suppresses HIV-1 Replication" Viruses 13, no. 8: 1432. https://doi.org/10.3390/v13081432
APA StyleLondhe, R., & Kulkarni, S. (2021). HTLV-2 Encoded Antisense Protein APH-2 Suppresses HIV-1 Replication. Viruses, 13(8), 1432. https://doi.org/10.3390/v13081432