
Resistance Evaluation of Dominant Varieties against Southern Rice Black-Streaked Dwarf Virus in Southern China

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rice, Virus, and Vector Insects
2.2. Virus Inoculation Simultaneously for Multiple Varieties
2.3. Virus Inoculation Separately for Single Varieties
2.4. Observation of Symptom Severity
2.5. Measurement of Virus Accumulation in Infected Plants
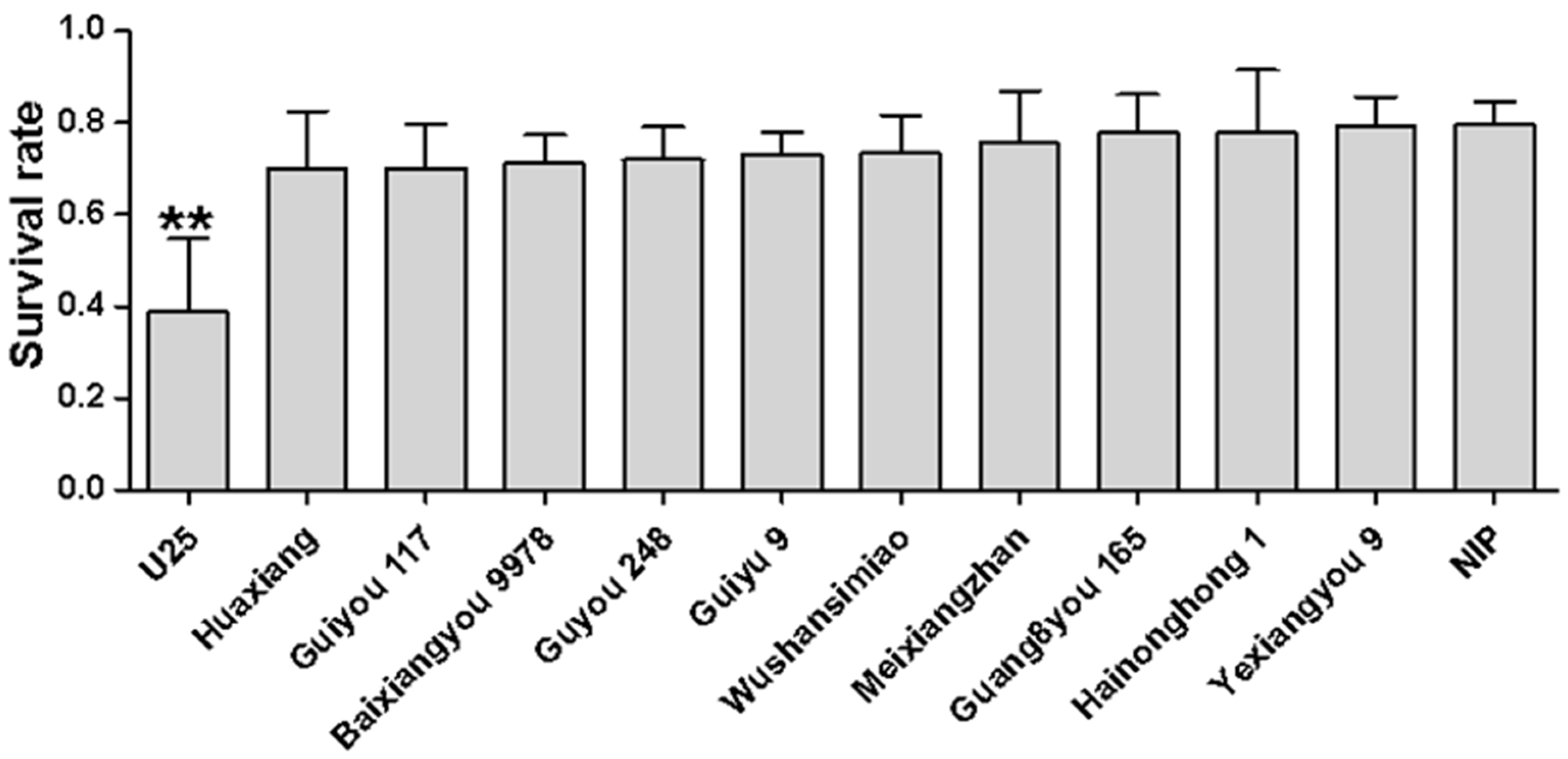
2.6. Determination of the WBPH Survival Rates on Different Rice Varieties
2.7. Evaluation of WBPH Virus Acquisition Ability from Infected Rice Plants
2.8. Date Analysis
3. Results
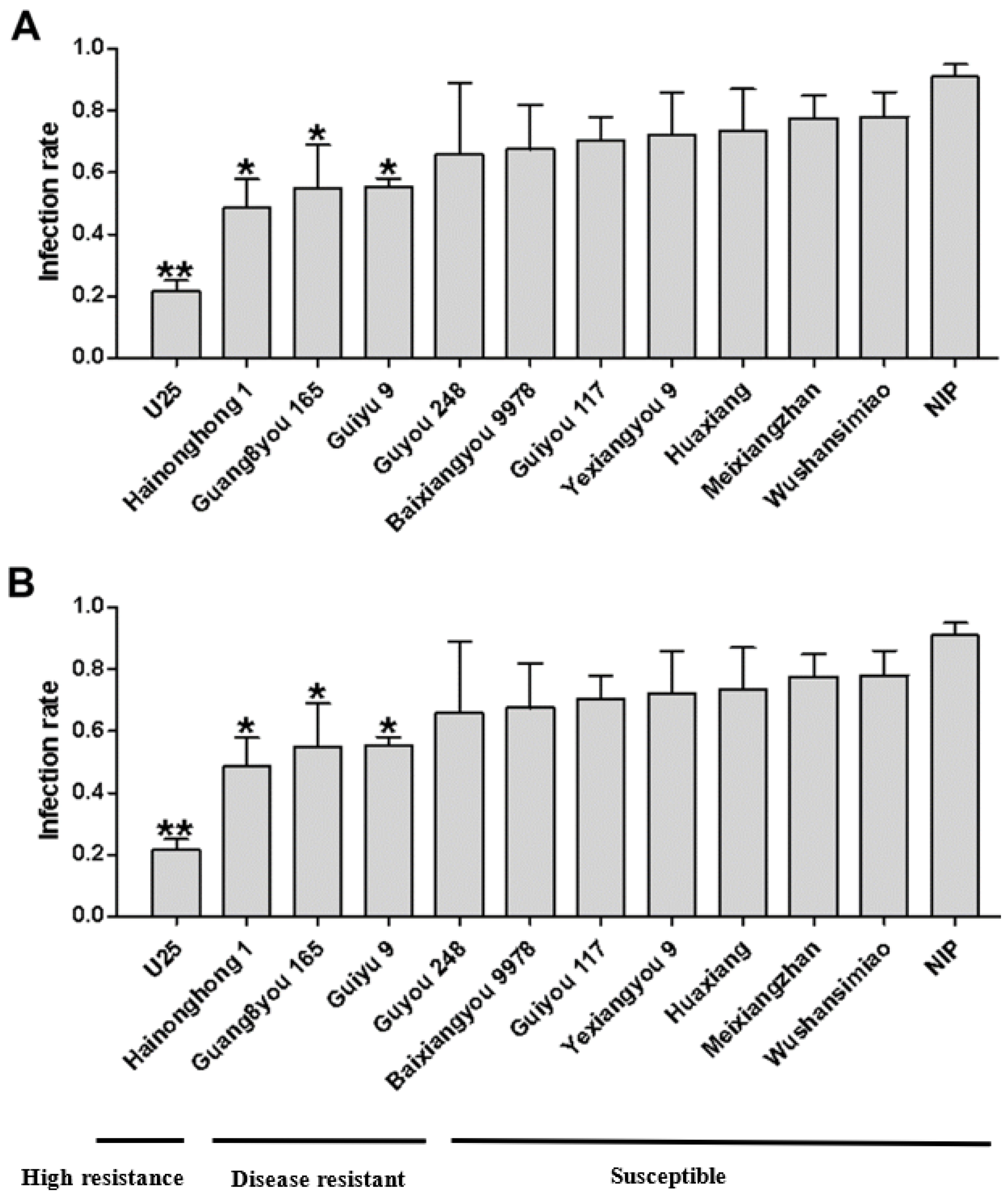
3.1. Resistance Based on Infection Rate
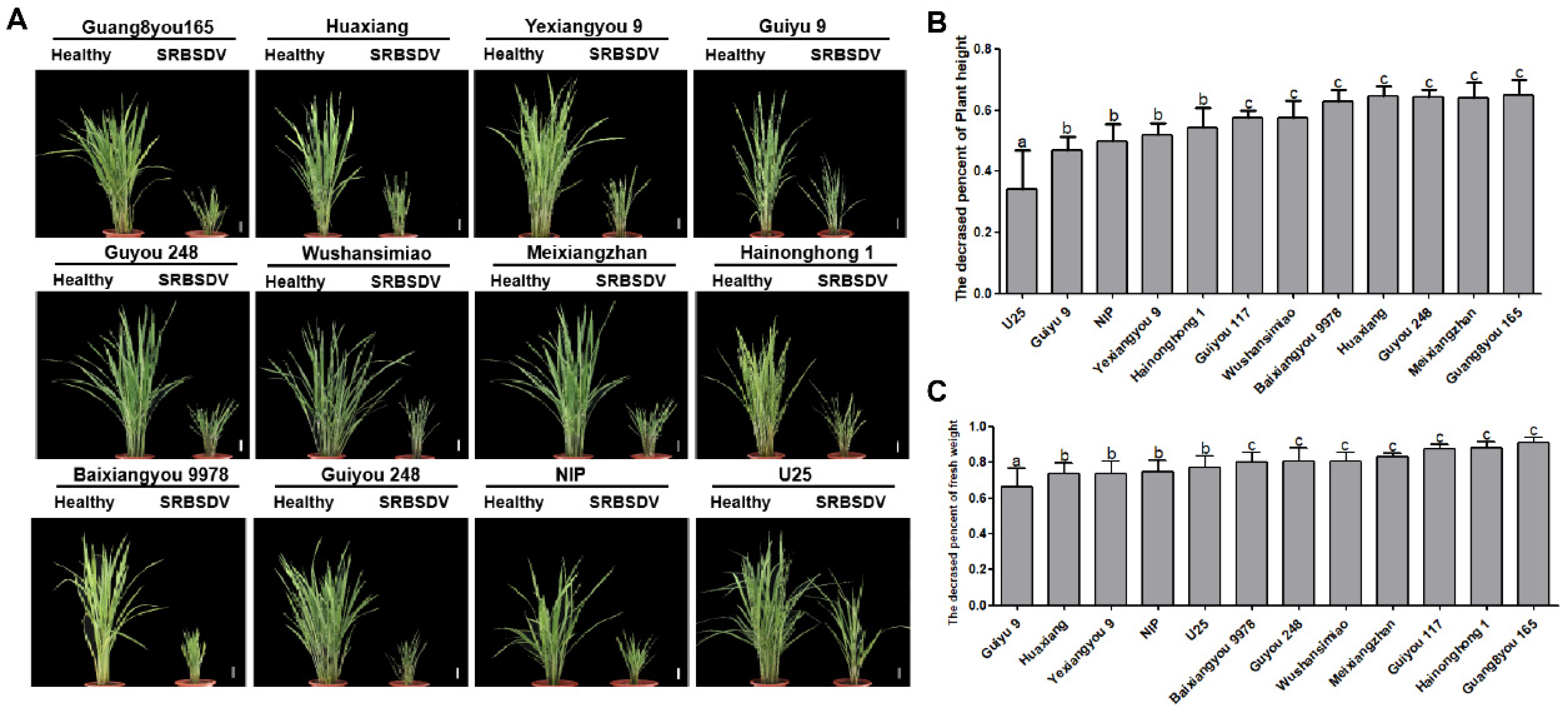
3.2. Resistance Based on Symptom Severity
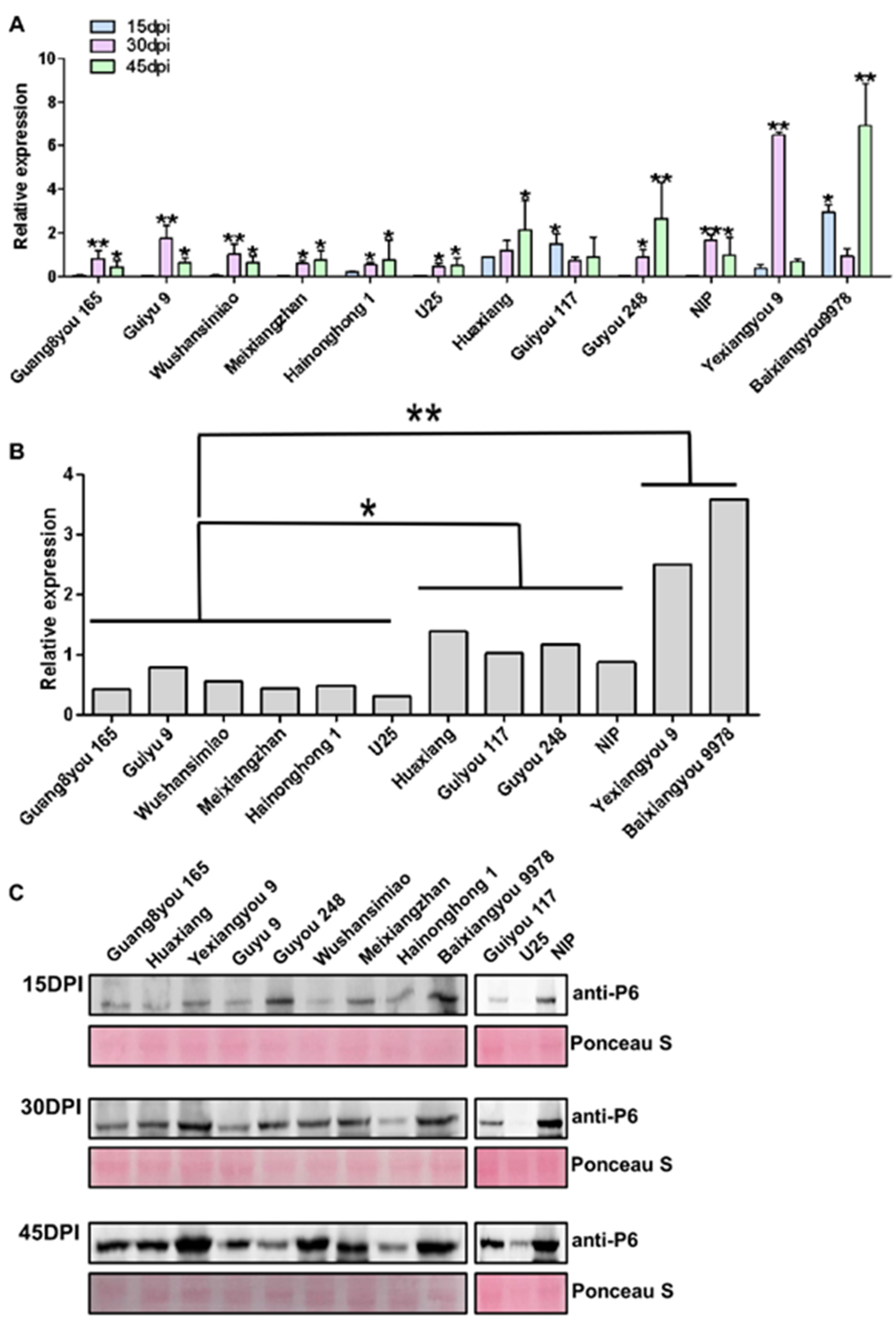
3.3. Resistance Based on Virus Accumulation in Infected Plants
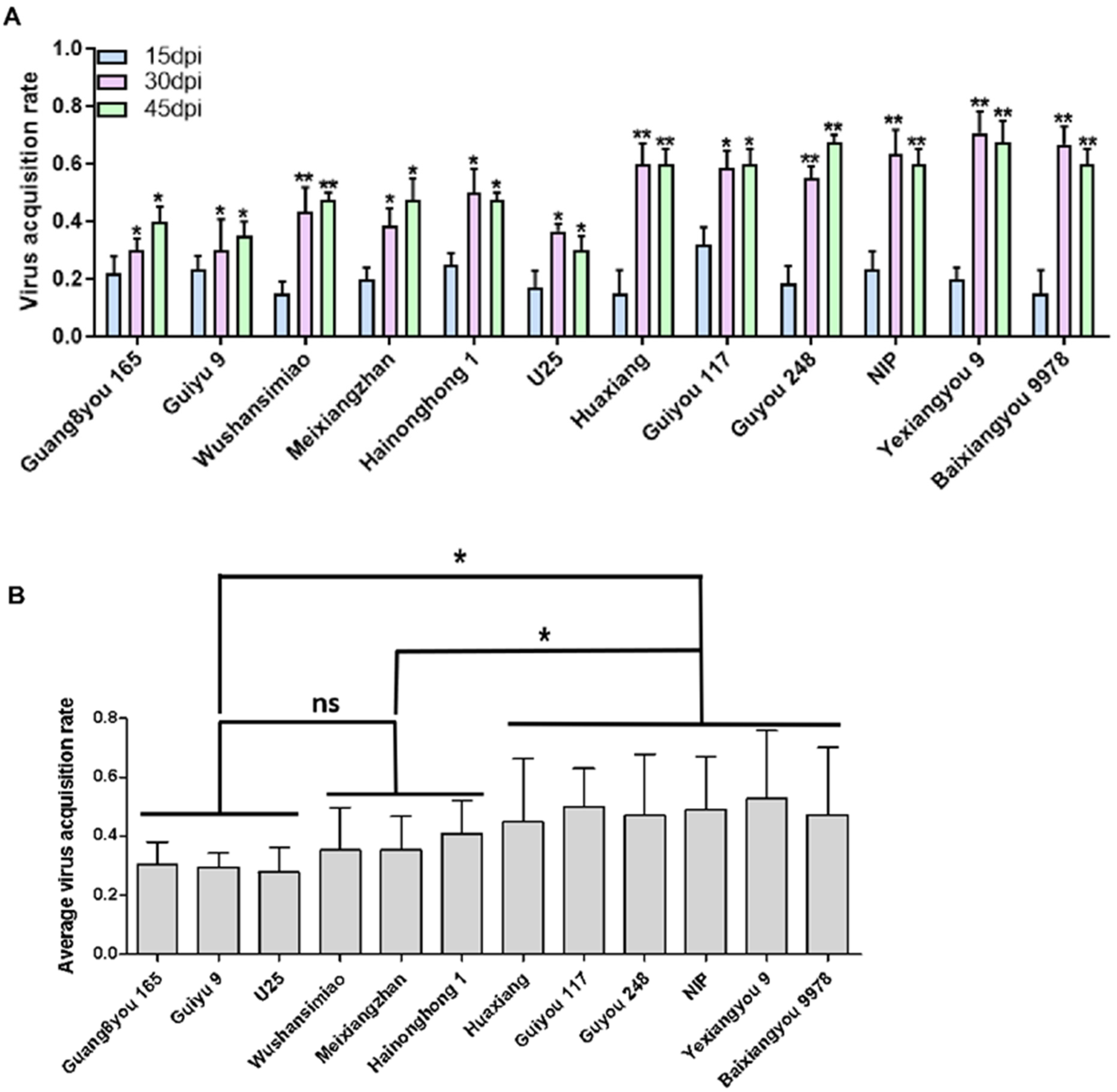
3.4. Resistance Based on Insect Vector Virus Acquisition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Uehara, L.T.; Shibaa, T.; Matsukura, K.; Ueno, T.; Hirae, M.; Sasaya, T. Detection and diagnosis of rice-infecting viruses. Front. Microbiol. 2013, 4, 289–295. [Google Scholar]
- Chen, H.; Chang, X.L.; Wang, Y.P.; Lu, M.H.; Liu, W.C.; Zhai, B.P.; Hu, G. The early northward migration of the white-backed planthopper (Sogatella furcifera) is often hindered by heavy precipitation in Southern China during the preflood season in May and June. Insects 2019, 10, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibino, H. Biology and epidemiology of rice viruses. Annu. Rev. Phytopathol. 1996, 34, 249–274. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lv, K.L.; Wang, M.Q.; Zhou, G.H. Investigation of viruses infecting rice in southern China using a multiplex RT-PCR assay. Crop Prot. 2017, 91, 8–12. [Google Scholar] [CrossRef]
- Anh, T.H.; Zhang, H.M.; Yang, J.; Chen, J.P.; Eugénie, H.; Zhou, G.H.; Vien, N.V.; Cheng, J.A. Identification, Characterization, and distribution of Southern rice black-streaked dwarf virus in Vietnam. Plant Dis. 2011, 95, 1063–1069. [Google Scholar]
- Matsukura, K.; Towata, T.; Sakai, J.; Onuki, M.; Okuda, M.; Matsumura, M. Dynamics of Southern rice black-streaked dwarf virus in rice and implication for virus acquisition. Phytopathology 2013, 103, 509–512. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.H.; Xu, D.L.; Xu, D.G.; Zhang, M.X. Southern rice black-streaked dwarf virus: A white-backed planthopper-transmitted fijivirus threatening rice production in Asia. Front. Microbiol. 2013, 4, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.H.; Wen, J.J.; Cai, D.J.; Li, P.; Xu, D.L.; Zhang, S.G. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae. Chin. Sci. Bull. 2008, 53, 3677–3685. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, J.; Zhou, G.H.; Zhang, H.M.; Chen, J.P.; Adams, M.J. The Complete genome sequence of two isolates of Southern rice black-streaked dwarf virus, a new bember of the genus fijivirus. J. Phytopathol. 2010, 158, 733–737. [Google Scholar] [CrossRef]
- Xu, D.; Zhong, T.; Feng, W.; Zhou, G.H. Tolerance and responsive gene expression of Sogatella furcifera under extreme temperature stresses are altered by its vectored plant virus. Sci. Rep. 2016, 6, 31521–31530. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Gong, Y.Q.; Fang, R.X. Research progress and perspective of tripartite interaction of virus-vector-plant in vector-borne viral diseases. Bull. Chin. Acad. Sci. 2017, 32, 845–855. (In Chinese) [Google Scholar]
- Towata, T.; Matsukura, K.; Sanada-Morimura, S.; Matsumura, M. Varietal differences in ovicidal response to the white-backed planthopper Sogatella furcifera (Hemiptera: Delphacidae) and susceptibility to Southern rice black-streaked dwarf virus in rice. Appl. Entomol. Zool. 2017, 52, 615–621. [Google Scholar] [CrossRef]
- Zhou, C.; Yang, H.; Yang, H.; Wang, Z.; Long, G.Y.; Jin, D.C. Effects of sublethal concentrations of deltamethrin on fitness of white-backed planthopper, Sogatella furcifera (Horváth). Int. J. Pest Manag. 2019, 65, 165–170. [Google Scholar] [CrossRef]
- Dhaka, S.S.; Rai, M.; Rai, M.; Yadav, A. Field evaluation of novel insecticides against brown planthopper (Nilaparvata lugens) and white backed planthopper (Sogatella furcifera) in rice. Indian J. Agric. Sci. 2020, 90, 1528–1531. [Google Scholar]
- Li, Z.; Zhang, T.; Huang, X.; Zhou, G. Impact of two reoviruses and their coinfection on the rice RNAi system and vsiRNA production. Viruses 2018, 10, 594. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.N.; Wang, Q.; Huang, D.Q.; Xu, Q.F.; Zhou, X.P.; Wu, J.X. Rice black-streaked dwarf virus P10 suppresses protein kinase C in insect vector through changing the subcellular localization of LsRACK1. Phil. Trans. R. Soc. B 2019, 374, 20180315. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wu, C.; Xiao, Y. Preference assessment of white-backed planthopper feeding on rice. J. Vis. Exp. 2020, 159, e59323. [Google Scholar]
- Zhai, B.P.; Zhou, G.H.; Tao, X.R.; Chen, X.; Shen, H.M. Macroscopic patterns and microscopic mechanisms of the outbreak of rice planthoppers and epidemic SRBSDV. Chin. J. Appl. Entomol. 2011, 48, 480–487. (In Chinese) [Google Scholar]
- Liu, W.C.; Lu, M.H.; Huang, C.; Zhou, G.H.; Hoang, A.T. Occurrence and forecasting practices of southern rice black-streaked dwarf virus disease in China. China Plant Prot. 2016, 36, 20–26. (In Chinese) [Google Scholar]
- Caldwell, R.M. Breeding for general and/or specific plant disease resistance. In Proceedings of the International Wheat Genetics Symposium, 3rd ed.; Finlay, K.W., Shephard, K.W., Eds.; Australian Academy of Sciences: Canberra, Australia, 1968; pp. 263–272. [Google Scholar]
- Hiebert, C.W.; Julian, B.; Thomas, J.B.; McCallum, B.D.; Humphreys, D.G.; DePauw, R.M.; Hayden, M.J.; Mago, R.; Schnippenkoetter, W.; Spielmeyer, W. An introgression on wheat chromosome 4DL in RL6077 (Thatcher*6/PI 250413) confers adult plant resistance to stripe rust and leaf rust (Lr67). Theor. Appl. Genet. 2010, 121, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.H.; Zhou, J.H.; Gong, X.P.; Zhao, G.Y.; Jia, J.Z.; Qi, X.Q. Identification and validation of a major quantitative trait locus for slow-rusting resistance to stripe rust in wheat. J. Integr. Plant Biol. 2012, 54, 330–344. [Google Scholar] [CrossRef] [PubMed]
- Huerta, E.J.; Singh, R.; Crespo, H.L.; Villaseñor, M.H.; Rodriguez, G.M.; Dreisigacker, S.; Barcenas, S.D.; Lagudah, E. Adult plant slow rusting genes confer gigh levels of resistance to rusts in bread wheat cultivars from Mexico. Front. Plant Sci. 2020, 11, 824. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.P.; Li, W.L.; Huang, X.Q.; Chen, B.; Zhang, T.; Zhou, G.H. Symptoms and yield loss caused by rice stripe mosaic virus. Virol. J. 2019, 16, 145–154. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, G.H. Advances in the researches on the southern rice black-streaked dwarf disease. J. Plant Prot. 2017, 44, 896–904. (In Chinese) [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Zhao, Y.; Liang, Z.; Wu, R.; Chen, B.; Zhang, T.; Yang, X.; Zhou, G. Resistance Evaluation of Dominant Varieties against Southern Rice Black-Streaked Dwarf Virus in Southern China. Viruses 2021, 13, 1501. https://doi.org/10.3390/v13081501
Zhou S, Zhao Y, Liang Z, Wu R, Chen B, Zhang T, Yang X, Zhou G. Resistance Evaluation of Dominant Varieties against Southern Rice Black-Streaked Dwarf Virus in Southern China. Viruses. 2021; 13(8):1501. https://doi.org/10.3390/v13081501
Chicago/Turabian StyleZhou, Shunkang, Yaling Zhao, Zhenyi Liang, Ruifeng Wu, Biao Chen, Tong Zhang, Xin Yang, and Guohui Zhou. 2021. "Resistance Evaluation of Dominant Varieties against Southern Rice Black-Streaked Dwarf Virus in Southern China" Viruses 13, no. 8: 1501. https://doi.org/10.3390/v13081501
APA StyleZhou, S., Zhao, Y., Liang, Z., Wu, R., Chen, B., Zhang, T., Yang, X., & Zhou, G. (2021). Resistance Evaluation of Dominant Varieties against Southern Rice Black-Streaked Dwarf Virus in Southern China. Viruses, 13(8), 1501. https://doi.org/10.3390/v13081501