
COVID-19 Convalescent Plasma Is More than Neutralizing Antibodies: A Narrative Review of Potential Beneficial and Detrimental Co-Factors

 ,
,  , ,
, , {kind=link}
Abstract
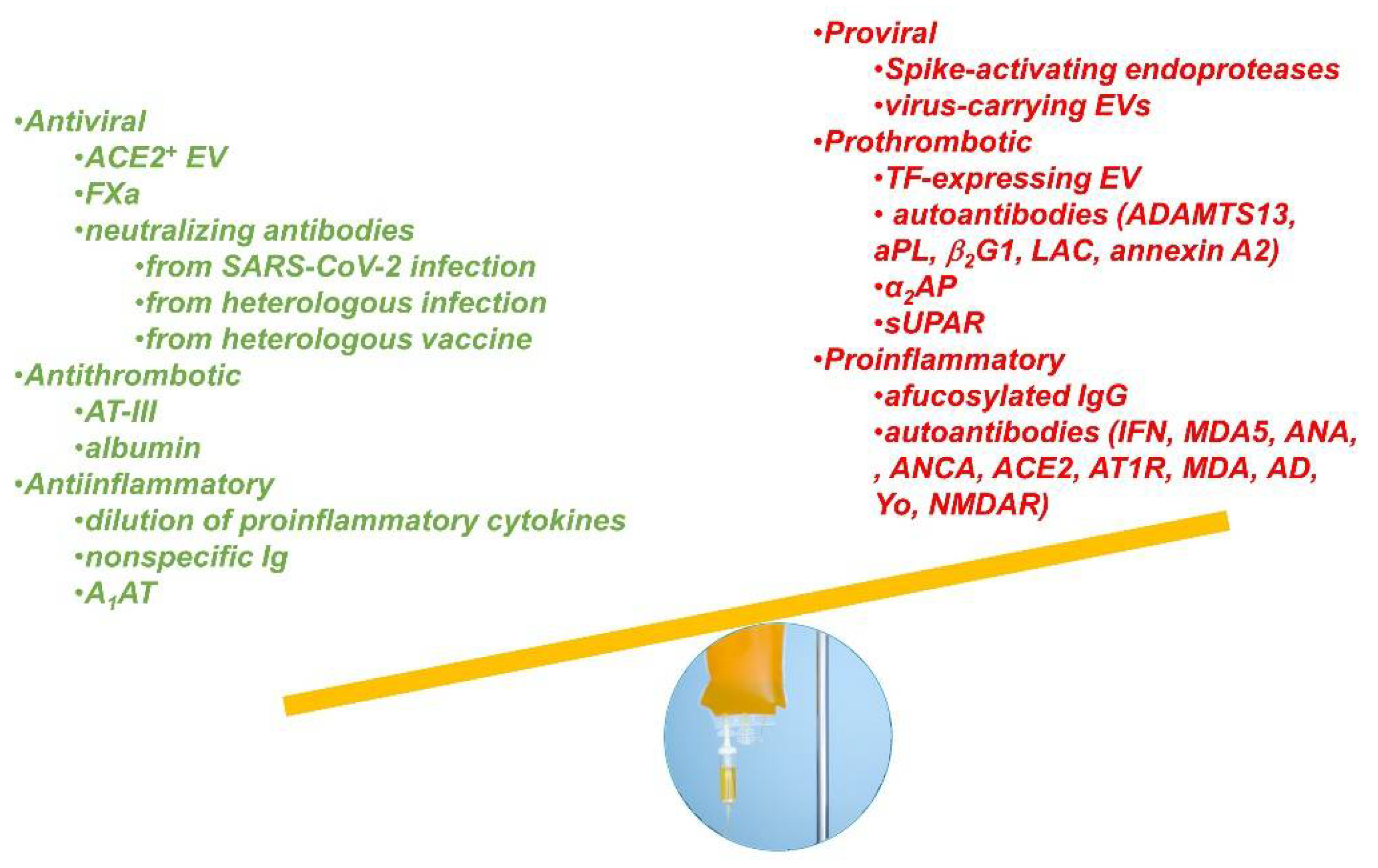
:1. Introduction
2. Potential Beneficial Factors in CCP
2.1. Immunomodulatory and Anti-Inflammatory Properties of CCP
2.2. Anti-Thrombotic Effect of CCP
- Antithrombin III is a universal constituent of donor plasma and works by improving the efficacy of heparin, which is one of the cornerstones of current COVID-19 management. Since AT-III levels are low in COVID-19 patients, it has been hypothesized that antithrombin III from CCP reduces the thrombotic risk in COVID-19 [26], but this has never been formally proven and no randomized controlled trial to date has reported a reduction in thrombotic events in the CCP arm;
- Albumin has been the object of intense research in the past few months. In an observational prospective cohort study, Violi and colleagues observed that albumin supplementation dampened hypercoagulability (measured as a reduction in D-dimer levels) in COVID-19 patients [28]. Similarly, a retrospective study by Kheir and colleagues found that higher albumin levels on admission were associated with a lower incidence of adverse outcomes, including venous thromboembolism (VTE), acute respiratory distress syndrome (ARDS) development, and intensive care unit (ICU) stay in COVID-19 patients [29].
2.3. Direct Antiviral Effects from CCP
- Extracellular vesicles (EVs) are also a universal component of donor plasma. EVs are lipid-bound vesicles secreted by cells into the extracellular space. The three main subtypes of EVs are micro-vesicles, exosomes, and apoptotic bodies. ACE2-positive EVs could act as decoy receptors since virions attaching to these EVs cannot complete a replicative cycle [30]. Recent experimental data show that ACE2-positive EVs can block SARS-CoV-2 spike-dependent infection [31]. EVs from plasma contain several other biomolecules such as miRNAs, proteins/cytokines, lipids, and glycan signatures that may alter the immune response to SARS-CoV-2 infection [32], but to date, a role for EVs in mediating the protective effect of CCP has not been demonstrated in vivo;
- Coagulation factor Xa (FXa) binds to and cleaves spike protein but produces a different cleavage pattern than that of furin and TMPRSS2, and, contrarily, what had been hypothesized initially [33], blocks S protein binding to ACE2. The effect was pronounced for the ancestral wild-type variant but was diminished in the B.1.1.7 variant. Exogenous FXa protected mice from lethal infection in a humanized hACE2 mouse model of COVID-19 using the wild-type variant but not the B.1.1.7 variant. The antiviral effect of FXa was attenuated by the direct FXa inhibitor rivaroxaban but not the indirect inhibitor fondaparinux, both in vivo and in vitro [34];
- Cross-reactive neutralizing antibodies: The basis for heterologous immune responses to SARS-CoV-2 is likely due to cross-reactivity between the surface antigens. Antigenic cross-reactivity can derive from previous exposure to a variety of …
- pathogens
- ○
- Seasonal coronaviruses [35]: Patients with severe COVID-19 had significantly lower levels of OC43 and HKU1 [36] or significantly higher NL63 and 229E [37] nucleoprotein-specific antibodies compared with other COVID-19 patients. The prognostic role of low OC43 antibodies was confirmed in another study: OC43 negative inpatients had an increased risk of severe disease (adjusted odds ratio 2.8), higher than the risk conferred by increased age or body mass index, and lower than the risk by male sex [35]. These findings may also imply convalescent plasma collections (e.g., CCP units with greater NL63 antibody responses and lower HKU1 antibodies) had higher neutralizing antibodies to the SARS-CoV-2 receptor-binding domain (RBD) [38]. Another study found better outcomes in recipients of CCP units with higher anti-NL63 or anti-OC43 antibodies [39];
- ○
- Influenza virus A(H3N2): Antibody binding to an epitope region from SARS-CoV-2 nucleocapsid, termed Ep9, is associated with greater COVID-19 disease severity [40]. Bioinformatics analysis identified the neuraminidase protein (not present in the influenza vaccine) of influenza virus A(H3N2) as responsible, a strain that circulated widely in 2014 [41];
- ○
- Acute malaria infection: Plasmodium infection induces cross-reactive antibodies to carbohydrate epitopes on the SARS-CoV-2 spike protein [42];
- ○
- Natural ABO isoagglutinins: The ABO blood group affects COVID-19 incidence and severity, as well as the type and duration of the cellular immune response [43]. Analogous to the events of SARS-CoV-1, it was hypothesized that natural isoagglutinins act as neutralizing antibodies owing to ABO antigens being carried over on virion envelope [44], although the evidence to date is weak [45].
- vaccination:
- ○
- MMR (measles-mumps-rubella) or Tdap (tetanus-diphtheria-acellular pertussis) vaccination [46]. Of interest, the SARS-CoV-2 spike protein displays biologically significant amino acid sequence similarities with paramyxovirus surface proteins [47]. A significant inverse correlation between mumps titers from MMR II and COVID-19 severity has also been reported [48];
- ○
- Influenza vaccination: Among 472,000 cases in Brazil, regression analysis showed an almost two-fold odds ratio for invasive ventilation, Intensive care unit (ICU) admission, and death in unvaccinated cases [49].
3. Potential Detrimental Factors in CCP
3.1. Direct Proviral Effect
- Spike-activating serine endoproteases can act as surrogates for TMPRSS2 at cleaving SARS-CoV-2 spike protein at the so-called furin cleavage site (FCS), creating S1 and S2 subunits. Thrombin is an endoprotease that increases SARS-COV-2 cell entry in vitro via this mechanism [33]. Since this enhances viral entry, more proteases can lead to more infection, but this has not been formally proven in vivo. A model of positive feedback was proposed whereby infection-induced hypercoagulation exacerbates SARS-CoV-2 infectivity. Anticoagulation is hence critical in managing COVID-19, and early intervention may provide collateral benefit by suppressing SARS-CoV-2 viral entry [33];
- Virus-carrying EVs: Despite SARS-COV-2 RNA viremia being extremely low and transient, SARS-CoV-2 RNA has been detected inside EVs [51]. Compared to the hyperinflammatory phase, EVs from the resolution phase induce opposing effects on eukaryotic translation and Notch signaling [52]. However, it is unclear whether these occur in recovered CCP donors and their infectious potential has not been established [53]. This concern represents an indication for applying pathogen reduction technologies to therapeutic CCP.
3.2. Pro-Coagulant Factors
- Tissue factor expressing EVs [56] are found in blood circulation, and their level parallels the intense thrombo-inflammatory state and thrombosis observed in severe COVID-19. However, we are not aware of studies examining the content and type of EVs in CCP. Clinical data using CCP did not identify a higher risk of thrombotic events suggesting that pro-coagulant tissue factors expressing EVs disappear quickly from the blood circulation upon resolution of the symptoms;
- Anti-ADAMTS13 autoantibodies: Doevelaar et al. reported antibodies to ADAMTS13 in 31 (34.4%) patients with COVID-19. Generation of ADAMTS13 antibodies was associated with a significantly lower ADAMTS13 activity, increased disease severity (a severe or critical disease in 90% vs. 62.3%), and a trend to higher mortality (35.5% vs. 18.6%). The median time to antibody development was 11 days after the first positive SARS-CoV-2-PCR specimen [57];
- Antiphospholipid (aPL) antibodies: A meta-analysis of over 1159 patients reported that aPL antibodies were detected in nearly half of patients with COVID-19, with a higher prevalence of aPL found in those with severe disease. However, there was no association between aPL positivity and disease outcomes, including thrombosis, invasive ventilation, and mortality [58]. Anti-cardiolipin IgA and IgM, and anti-β2 glycoprotein-1 IgA were found in 5–12% of hospitalized patients [59,60] and were elevated in severe COVID-19 [61]. Nevertheless, Borghi et al. reported that aPLs show a low prevalence in COVID-19 patients and are not associated with major thrombotic events. The aPLs in COVID-19 patients are mainly directed against β2-GPI but display an epitope specificity different from antibodies in aPL syndrome [62]. Lupus anticoagulant (LA), a misnomer for prothrombotic antibody, was found in 46.6% of hospitalized COVID-19 patients, but no association was found with mortality or the need for mechanical ventilation in survivors [63]. Most importantly, LA is transient, but other aPLs are persistent [64] and potentially found in CCP donors. Anti-prothrombin antibody levels are associated with disease severity and anti-SARS-CoV-2 antibody levels [65];
- Autoantibodies against annexin A2, which are known to induce systemic thrombosis, cell death, and non-cardiogenic pulmonary edema, were elevated among 86 hospitalized COVID-19 patients and predicted mortality (OR = 9.3) [66];
- α(2)-antiplasmin (α2AP), various fibrinogen chains, and Serum Amyloid A (SAA) are substantially increased and trapped in the solubilized fibrinolytic-resistant pellet deposits found in plasma from patients experiencing long COVID-19/post-acute sequelae of COVID-19 (PASC) syndrome. Albeit, such patients are unlikely to donate CCP and mild presentations could pass screening visits [67];
- Soluble urokinase-type plasminogen activator receptor (sUPAR) is highly expressed by an abnormally expanded circulating myeloid cell population in severe COVID-19 patients with ARDS [68]. Plasma sUPAR level was found to be linked to a characteristic proteomic signature of plasma, linked to coagulation disorders, and complement activation.
3.3. Proinflammatory or Immunosuppressive Factors
- Afucosylated IgG defines an exacerbated phenotype in COVID-19: afucosylated immune complexes in the lungs trigger an inflammatory infiltrate and cytokine production dependent on the expression of the receptor for afucosylated IgGs, FcγRIIIa (CD16a) in monocytes [43]. Accordingly, elevated frequencies of CD16a+ monocytes were another antecedent in patients with more severe outcomes [43]. Immune complexes contained recombinant SARS-CoV-2 spike protein and aberrantly glycosylated anti-spike IgG with enhanced platelet-mediated thrombosis on von Willebrand Factor in vitro [69];
- Autoantibodies: SARS-CoV-2 infection can trigger autoimmune diseases such as myocarditis, and many single cases have been reported in the literature. In this review, we focus on large case series that help assess the prevalence of autoantibodies.
- ○
- Autoantibodies against interferons (IFNs) are commonly found in 40% of systemic lupus erythematosus patients. Bastard et al. identified nAb against type I IFN-α2 and IFN-ω in about 10% of patients with severe COVID-19 pneumonia, but not in patients with an asymptomatic or mild disease [70]. This cohort has the highest likelihood of having antiviral nAbs [71] and hence more likely to be selected as CCP donors. Accordingly, Vazquez et al. found nAbs to IFNs in 3% (4/116) of CCP donors [72]. By neutralizing one of the key mediators of the effector arm of the immune response, these antibodies may function as immune suppressants, which could help or hurt the recipient patient depending on the stage of the disease;
- ○
- Autoantibodies against melanoma differentiation-associated gene 5 (MDA5) characterizes a subtype of dermatomyositis (DM) and were found in 48.2% (132/274) of COVID-19 patients. The anti-MDA5 Ab positive patients tended to represent severe COVID-19 (88.6% vs. 66.9%). The titer of Ab to MDA5 was significantly elevated in non-surviving patients, and the positive rate was also higher than in survivors (23.5% vs. 12.0). With regards to those patients with severe COVID-19, high titers of Ab to MDA5 (≥10.0 U/mL) were more prevalent in those who did not survive (31.2% vs. 14.0%) [73]. Of interest, both MDA5-associated DM and COVID-19 can involve the lungs, skin, and skeletal muscles. The initial radiological features of lung pathology in DM patients with Abs to MDA5 are mainly subpleural ground-glass opacities or mixed with consolidation and signs of acute respiratory distress syndrome (ARDS) resembling severe and critical COVID-19. MDA5 is a crucial cytoplasmic sensor for viral RNA, and its expression is induced by RNA viruses (including SARS-CoV-2 [74]). Viral infection activates the expression of antiviral type I and III interferons (IFNs) and other inflammatory cytokines;
- ○
- Antinuclear autoantibodies (ANA) were found in 11–57% [59,60,75] of hospitalized COVID-19 patients. Woodruf et al. identified ANA in 44% of 31 critically-ill patients with COVID-19 with no known history of autoimmunity [76]. Specifically, Gomes et al. showed that antibodies to DNA determined hospital admission and correlated strongly with the later development of severe disease, showing a positive predictive value of 89.5% and accounting for 22% of total severe cases [77]. Anti-extractable nuclear antigen (ENA) antibodies were reported in 2.5% of hospitalized COVID-19 patients [60];
- ○
- ○
- IgM autoantibodies against ACE2 (the cellular receptor for SARS-CoV-2 spike protein) were detected in 27% of 66 severe COVID-19 patients vs. 3.8% of 52 non-hospitalized patients [78]. If and how they contribute to angiocentric pathology remains unknown. The antibodies do not undergo class-switching to IgG, suggesting a T cell-independent antibody response. Purified IgM from anti-ACE2 patients activates complement;
- ○
- Autoantibodies against angiotensin II type 1 receptor (AT1R): No statistically significant differences were found between COVID-19 cases and controls. However, there were trends toward a higher proportion with AT1R autoantibody positivity among severe cases versus controls (32% vs. 11%) and higher levels in those with mild COVID-19 compared with controls (median 9.5 U/mL vs. 5.9 U/mL [79]);
- ○
- Autoantibodies against anti-malondialdehyde (MDA) and anti-adipocyte-derived protein antigens (AD) are more frequent in lean than in obese COVID-19 patients compared to uninfected controls. However, serum levels of these autoantibodies are always higher in obese versus lean COVID-19 patients and associated with CRP levels [80];
- ○
- Anti-neuronal or anti-glial autoantibodies (e.g., against Yo or NMDA receptor), which theoretically crossed a leaky brain-blood barrier, were universally detected in plasma and cerebrospinal fluid of 11 severely ill COVID-19 patients presenting unexplained neurological symptoms [81].
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mahase, E. Covid-19: WHO declares pandemic because of “alarming levels” of spread, severity, and inaction. BMJ 2020, 368, m1036. [Google Scholar] [CrossRef] [Green Version]
- Boban, M. Novel coronavirus disease (COVID-19) update on epidemiology, pathogenicity, clinical course and treatments. Int. J. Clin. Pract. 2021, 75, e13868. [Google Scholar] [CrossRef]
- Pan, H.; Peto, R.; Henao-Restrepo, A.M.; Preziosi, M.P.; Sathiyamoorthy, V.; Abdool Karim, Q.; Alejandria, M.M.; Hernández García, C.; Kieny, M.P.; Malekzadeh, R.; et al. Repurposed Antiviral Drugs for Covid-19—Interim WHO Solidarity Trial Results. N. Engl. J. Med. 2021, 384, 497–511. [Google Scholar] [CrossRef]
- Shen, C.; Wang, Z.; Zhao, F.; Yang, Y.; Li, J.; Yuan, J.; Wang, F.; Li, D.; Yang, M.; Xing, L.; et al. Treatment of 5 Critically Ill Patients with COVID-19 With Convalescent Plasma. JAMA 2020, 323, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Franchini, M.; Liumbruno, G.M.; Piacentini, G.; Glingani, C.; Zaffanello, M. The Three Pillars of COVID-19 Convalescent Plasma Therapy. Life 2021, 11, 354. [Google Scholar] [CrossRef]
- Focosi, D.; Franchini, M. COVID-19 convalescent plasma therapy: Hit fast, hit hard! Vox Sang. 2021. [Google Scholar] [CrossRef]
- Joyner, M.J.; Carter, R.E.; Senefeld, J.W.; Klassen, S.A.; Mills, J.R.; Johnson, P.W.; Theel, E.S.; Wiggins, C.C.; Bruno, K.A.; Klompas, A.M.; et al. Convalescent Plasma Antibody Levels and the Risk of Death from Covid-19. N. Engl. J. Med. 2021, 384, 1015–1027. [Google Scholar] [CrossRef]
- Libster, R.; Pérez Marc, G.; Wappner, D.; Coviello, S.; Bianchi, A.; Braem, V.; Esteban, I.; Caballero, M.T.; Wood, C.; Berrueta, M.; et al. Early High-Titer Plasma Therapy to Prevent Severe Covid-19 in Older Adults. N. Engl. J. Med. 2021, 384, 610–618. [Google Scholar] [CrossRef]
- Maor, Y.; Cohen, D.; Paran, N.; Israely, T.; Ezra, V.; Axelrod, O.; Shinar, E.; Izak, M.; Rahav, G.; Rahimi-Levene, N.; et al. Compassionate use of convalescent plasma for treatment of moderate and severe pneumonia in COVID-19 patients and association with IgG antibody levels in donated plasma. EClinicalMedicine 2020, 26, 100525. [Google Scholar] [CrossRef]
- Natarajan, H.; Crowley, A.R.; Butler, S.E.; Xu, S.; Weiner, J.A.; Bloch, E.M.; Littlefield, K.; Wieland-Alter, W.; Connor, R.I.; Wright, P.F.; et al. Markers of Polyfunctional SARS-CoV-2 Antibodies in Convalescent Plasma. mBio 2021, 12, e00765-21. [Google Scholar] [CrossRef]
- Franchini, M.; Glingani, C.; Liumbruno, G.M. Potential mechanisms of action of convalescent plasma in COVID-19. Diagnosis 2021. [Google Scholar] [CrossRef]
- Casadevall, A.; Dadachova, E.; Pirofski, L.A. Passive antibody therapy for infectious diseases. Nat. Rev. Microbiol. 2004, 2, 695–703. [Google Scholar] [CrossRef]
- Acosta-Ampudia, Y.; Monsalve, D.M.; Rojas, M.; Rodríguez, Y.; Gallo, J.E.; Salazar-Uribe, J.C.; Santander, M.J.; Cala, M.P.; Zapata, W.; Zapata, M.I.; et al. COVID-19 convalescent plasma composition and immunological effects in severe patients. J. Autoimmun. 2021, 118, 102598. [Google Scholar] [CrossRef] [PubMed]
- Bandopadhyay, P.; Rozario, R.; Lahiri, A.; Sarif, J.; Ray, Y.; Paul, S.R.; Roy, R.; Maiti, R.; Chaudhuri, K.; Bagchi, S.; et al. Nature and dimensions of the systemic hyper-inflammation and its attenuation by convalescent plasma in severe COVID-19. J. Infect. Dis. 2021, jiab010. [Google Scholar] [CrossRef] [PubMed]
- Garraud, O.; Heshmati, F.; Pozzetto, B.; Lefrere, F.; Girot, R.; Saillol, A.; Laperche, S. Plasma therapy against infectious pathogens, as of yesterday, today and tomorrow. Transfus. Clin. Biol. J. Soc. Fr. Transfus. Sang. 2016, 23, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Perotti, C.; Baldanti, F.; Bruno, R.; Delfante, C.; Seminari, E.; Casari, S.; Percivalle, E.; Glingani, C.; Musella, V.; Belliato, M.; et al. Mortality reduction in 46 severe Covid-19 patients treated with hyperimmune plasma. A proof of concept single arm multicenter interventional trial. Haematologica 2020, 105, 2834–2840. [Google Scholar] [CrossRef] [PubMed]
- Salazar, E.; Perez, K.K.; Ashraf, M.; Chen, J.; Castillo, B.; Christensen, P.A.; Eubank, T.; Bernard, D.W.; Eagar, T.N.; Long, S.W.; et al. Treatment of Coronavirus Disease 2019 (COVID-19) Patients with Convalescent Plasma. Am. J. Pathol. 2020, 190, 1680–1690. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Gazca, J.C.; Priesca-Marín, J.M.; Ojeda-Laguna, M.; Garces-Eisele, J.; Soto-Olvera, S.; Palacios-Alonso, A.; Izquierdo-Vega, J.; Chacon-Cano, R.; Arizpe-Bravo, D.; López-Trujillo, M.A.; et al. Infusion of convalescent plasma is associated with clinical improvement in critically ill patients with covid-19: A pilot study. Rev. Investig. Clin. Organo Hosp. Enferm. Nutr. 2020, 72, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Shen, C.; Xia, C.; Huang, X.; Fu, Y.; Tian, L. A Retrospective Study on the Effects of Convalescent Plasma Therapy in 24 Patients Diagnosed with COVID-19 Pneumonia in February and March 2020 at 2 Centers in Wuhan, China. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e928755. [Google Scholar] [CrossRef]
- Franchini, M.; Glingani, C.; Morandi, M.; Corghi, G.; Cerzosimo, S.; Beduzzi, G.; Storti, A.; Di Stasi, V.; Rastrelli, G.; Vignozzi, L.; et al. Safety and Efficacy of Convalescent Plasma in Elderly COVID-19 Patients: The RESCUE Trial. Mayo Clin. Proc. Innov. Qual. Outcomes 2021, 5, 403–412. [Google Scholar] [CrossRef]
- Bonny, T.S.; Patel, E.U.; Zhu, X.; Bloch, E.M.; Grabowski, M.K.; Abraham, A.G.; Littlefield, K.; Shrestha, R.; Benner, S.E.; Laeyendecker, O.; et al. Cytokine and Chemokine Levels in Coronavirus Disease 2019 Convalescent Plasma. Open Forum Infect. Dis. 2021, 8, ofaa574. [Google Scholar] [CrossRef]
- Yang, C.; Chapman, K.R.; Wong, A.; Liu, M. α1-Antitrypsin deficiency and the risk of COVID-19: An urgent call to action. Lancet Respir. Med. 2021, 9, 337–339. [Google Scholar] [CrossRef]
- Franchini, M.; Marano, G.; Cruciani, M.; Mengoli, C.; Pati, I.; Masiello, F.; Veropalumbo, E.; Pupella, S.; Vaglio, S.; Liumbruno, G.M. COVID-19-associated coagulopathy. Diagnosis 2020, 7, 357–363. [Google Scholar] [CrossRef]
- Dobesh, P.P.; Trujillo, T.C. Coagulopathy, Venous Thromboembolism, and Anticoagulation in Patients with COVID-19. Pharmacotherapy 2020, 40, 1130–1151. [Google Scholar] [CrossRef] [PubMed]
- Kichloo, A.; Dettloff, K.; Aljadah, M.; Albosta, M.; Jamal, S.; Singh, J.; Wani, F.; Kumar, A.; Vallabhaneni, S.; Khan, M.Z. COVID-19 and Hypercoagulability: A Review. Clin. Appl. Thromb. Hemost. Off. J. Int. Acad. Clin. Appl. Thromb. Hemost. 2020, 26, 1076029620962853. [Google Scholar] [CrossRef] [PubMed]
- Gazzaruso, C.; Valenti, C.; Coppola, A.; Gallotti, P. Impact of convalescent and nonimmune plasma on mortality of patients with COVID-19: A potential role for antithrombin. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2020, 27, 637–638. [Google Scholar] [CrossRef] [PubMed]
- Violi, F.; Cangemi, R.; Romiti, G.F.; Ceccarelli, G.; Oliva, A.; Alessandri, F.; Pirro, M.; Pignatelli, P.; Lichtner, M.; Carraro, A.; et al. Is Albumin Predictor of Mortality in COVID-19? Antioxid. Redox Signal. 2020, 35, 139–142. [Google Scholar] [CrossRef]
- Violi, F.; Ceccarelli, G.; Loffredo, L.; Alessandri, F.; Cipollone, F.; D’Ardes, D.; D’Ettorre, G.; Pignatelli, P.; Venditti, M.; Mastroianni, C.M.; et al. Albumin Supplementation Dampens Hypercoagulability in COVID-19: A Preliminary Report. Thromb. Haemost. 2021, 121, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Kheir, M.; Saleem, F.; Wang, C.; Mann, A.; Chua, J. Higher albumin levels on admission predict better prognosis in patients with confirmed COVID-19. PLoS ONE 2021, 16, e0248358. [Google Scholar] [CrossRef]
- El-Shennawy, L.; Hoffmann, A.D.; Dashzeveg, N.K.; Mehl, P.J.; Yu, Z.; Tokars, V.L.; Nicolaescu, V.; Ostiguin, C.; Jia, Y.; Li, L.; et al. Circulating ACE2-expressing Exosomes Block SARS-CoV-2 Virus Infection as an Innate Antiviral Mechanism. bioRxiv 2020. [Google Scholar] [CrossRef]
- Cocozza, F.; Névo, N.; Piovesana, E.; Lahaye, X.; Buchrieser, J.; Schwartz, O.; Manel, N.; Tkach, M.; Théry, C.; Martin-Jaular, L. Extracellular vesicles containing ACE2 efficiently prevent infection by SARS-CoV-2 Spike protein-containing virus. J. Extracell. Vesicles 2020, 10, e12050. [Google Scholar] [CrossRef]
- ACTIV-3: Therapeutics for Inpatients with COVID-19—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04501978 (accessed on 17 July 2021).
- Kastenhuber, E.R.; Jaimes, J.A.; Johnson, J.L.; Mercadante, M.; Muecksch, F.; Weisblum, Y.; Bram, Y.; Schwartz, R.E.; Whittaker, G.R.; Cantley, L.C. Coagulation factors directly cleave SARS-CoV-2 spike and enhance viral entry. bioRxiv 2021. [Google Scholar] [CrossRef]
- Dong, W.; Wang, J.; Tian, L.; Zhang, J.; Mead, H.; Jaramillo, S.A.; Li, A.; Zumwalt, R.E.; Whelan, S.P.J.; Settles, E.W.; et al. FXa cleaves the SARS-CoV-2 spike protein and blocks cell entry to protect against infection with inferior effects in B.1.1.7 variant. bioRxiv 2021. [Google Scholar] [CrossRef]
- Dugas, M.; Grote-Westrick, T.; Merle, U.; Fontenay, M.; Kremer, A.E.; Vollenberg, R.; Lorentzen, E.; Tiwari-Heckler, S.; Duchemin, J.; Ellouze, S.; et al. Lack of antibodies against seasonal coronavirus OC43 nucleocapsid protein identifies patients at risk of critical COVID-19. J. Clin. Virol. 2020. [Google Scholar] [CrossRef]
- Dugas, M.; Grote-Westrick, T.; Vollenberg, R.; Lorentzen, E.; Brix, T.; Schmidt, H.; Tepasse, P.-R.; Kühn, J. Less severe course of COVID-19 is associated with elevated levels of antibodies against seasonal human coronaviruses OC43 and HKU1 (HCoV OC43, HCoV HKU1). Int. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Focosi, D.; Genoni, A.; Lucenteforte, E.; Tillati, S.; Tamborini, A.; Spezia, P.G.; Azzi, L.; Baj, A.; Maggi, F. Previous Humoral Immunity to the Endemic Seasonal Alphacoronaviruses NL63 and 229E Is Associated with Worse Clinical Outcome in COVID-19 and Suggests Original Antigenic Sin. Life 2021, 11, 298. [Google Scholar] [CrossRef] [PubMed]
- Morgenlander, W.R.; Henson, S.; Monaco, D.; Chen, A.; Littlefield, K.; Bloch, E.M.; Fujimura, E.; Ruczinski, I.; Crowley, A.R.; Natarajan, H.; et al. Antibody responses to endemic coronaviruses modulate COVID-19 convalescent plasma functionality. J. Clin. Investig. 2021. [Google Scholar] [CrossRef]
- Greenbaum, U.; Klein, K.; Martinez, F.; Song, J.; Thall, P.F.; Ramdial, J.L.; Knape, C.; Aung, F.M.; Scroggins, J.; Knopfelmacher, A.; et al. High levels of common cold coronavirus antibodies in convalescent plasma are associated with improved survival in COVID-19 patients. Front. Immunol. 2021. [Google Scholar] [CrossRef]
- Sen, S.R.; Sanders, E.C.; Gabriel, K.N.; Miller, B.M.; Isoda, H.M.; Salcedo, G.S.; Garrido, J.E.; Dyer, R.P.; Nakajima, R.; Jain, A.; et al. Predicting COVID-19 Severity with a Specific Nucleocapsid Antibody plus Disease Risk Factor Score. mSphere 2021, 6. [Google Scholar] [CrossRef]
- Sen, S.R.; Sanders, E.C.; Santos, A.M.; Bhuvan, K.; Tang, D.Y.; Gelston, A.A.; Miller, B.M.; Ricks-Oddie, J.L.; Weiss, G.A. Evidence for Deleterious Original Antigenic Sin in SARS-CoV-2 Immune Response. bioRxiv 2021. [Google Scholar] [CrossRef]
- Lapidus, S.; Liu, F.; Casanovas-Massana, A.; Dai, Y.; Huck, J.D.; Lucas, C.; Klein, J.; Filler, R.B.; Strine, M.S.; Sy, M.; et al. Plasmodium infection induces cross-reactive antibodies to carbohydrate epitopes on the SARS-CoV-2 Spike protein. medRxiv 2021. [Google Scholar] [CrossRef]
- Gil-Manso, S.; Miguens Blanco, I.; Motyka, B.; Halpin, A.; Lopez-Estaban, R.; Perez-Fernandez, V.A.; Carbonell-Munoz, D.; Lopez-Fernandez, L.A.; west, L.J.; Correa-Rocha, R.; et al. ABO blood group is involved in the quality of the specific immune response. bioRxiv 2021. [Google Scholar] [CrossRef]
- Focosi, D. Anti-A Isohemagglutinin titers and SARS-CoV2 neutralization: Implications for children and convalescent plasma selection. Br. J. Haematol. 2020, 190, e148–e150. [Google Scholar] [CrossRef] [PubMed]
- Deleers, M.; Breiman, A.; Daubie, V.; Maggetto, C.; Barreau, I.; Besse, T.; Clémenceau, B.; Ruvoën-Clouet, N.; Fils, J.F.; Maillart, E.; et al. Covid-19 and blood groups: ABO antibody levels may also matter. Int. J. Infect. Dis. Off. Publ. Int. Soc. Infect. Dis. 2020, 104, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Mayadas, T.; Mysore, V.; Cullere, X.; Settles, M.L.; Kattan, M.; Ji, X.; Desjardins, M.; Durbin-Johnson, B.; Gilboa, T.; Baden, L.; et al. Protective heterologous T cell immunity in COVID-19 induced by MMR and Tdap vaccine antigens. biorXiv 2021. [Google Scholar] [CrossRef]
- Ahmadi, E.; Zabihi, M.R.; Hosseinzadeh, R.; Noorbakhsh, F. SARS-CoV2 spike protein displays biologically significant similarities with paramyxovirus surface proteins; a bioinformatics study. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gold, J.E.; Baumgartl, W.H.; Okyay, R.A.; Licht, W.E.; Fidel, P.L.; Noverr, M.C.; Tilley, L.P.; Hurley, D.J.; Rada, B.; Ashford, J.W. Analysis of Measles-Mumps-Rubella (MMR) Titers of Recovered COVID-19 Patients. MBio 2020, 11, e02628-20. [Google Scholar] [CrossRef]
- Sardinha, D.M.; da Costa Lobato, D.; da Silva Ferreira, A.L.; Lima, K.V.; Lima, L.N. Analysis of 472,688 severe cases of COVID-19 in Brazil showed lower mortality in those vaccinated against influenza. medRxiv 2021. [Google Scholar] [CrossRef]
- Patidar, G.K.; Land, K.J.; Vrielink, H.; Rahimi-Levene, N.; Dann, E.J.; Al-Humaidan, H.; Spitalnik, S.L.; Dhiman, Y.; So-Osman, C.; Hindawi, S.I. Understanding the role of therapeutic plasma exchange in COVID-19: Preliminary guidance and practices. Vox Sang. 2021. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.; Nukala, S.B.; Srivastava, S.; Miyamoto, H.; Ismail, N.I.; Jousma, J.; Rehman, J.; Ong, S.-B.; Lee, W.H.; Ong, S.-G. Detection of viral RNA fragments in human iPSC cardiomyocytes following treatment with extracellular vesicles from SARS-CoV-2 coding sequence overexpressing lung epithelial cells. Stem Cell Res. Ther. 2020, 11, 514. [Google Scholar] [CrossRef]
- Lam, S.M.; Zhang, C.; Wang, Z.; Ni, Z.; Zhang, S.; Yang, S.; Huang, X.; Mo, L.; Li, J.; Lee, B.; et al. A multi-omics investigation of the composition and function of extracellular vesicles along the temporal trajectory of COVID-19. Nat. Metab. 2021. [Google Scholar] [CrossRef]
- Zani-Ruttenstock, E.; Antounians, L.; Khalaj, K.; Figueira, R.L.; Zani, A. The Role of Exosomes in the Treatment, Prevention, Diagnosis, and Pathogenesis of COVID-19. Eur. J. Pediatric Surg. 2021. [Google Scholar] [CrossRef]
- Marwah, V.; Choudhary, R.; Peter, D.; Bhati, G. Pulmonary thromboembolism post-COVID convalescent plasma therapy: Adding fuel to a smoldering fire! Adv. Respir. Med. 2021. [Google Scholar] [CrossRef]
- Joyner, M.J.; Bruno, K.A.; Klassen, S.A.; Kunze, K.L.; Johnson, P.W.; Lesser, E.R.; Wiggins, C.C.; Senefeld, J.W.; Klompas, A.M.; Hodge, D.O.; et al. Safety Update: COVID-19 Convalescent Plasma in 20,000 Hospitalized Patients. Mayo Clin. Proc. 2020, 95, 1888–1895. [Google Scholar] [CrossRef] [PubMed]
- Guervilly, C.; Bonifay, A.; Burtey, S.; Sabatier, F.; Cauchois, R.; Abdili, E.; Arnaud, L.; Lano, G.; Pietri, L.; Robert, T.; et al. Dissemination of extreme levels of extracellular vesicles: Tissue factor activity in patients with severe COVID-19. Blood Adv. 2021, 5, 628–634. [Google Scholar] [CrossRef]
- Doevelaar, A.A.N.; Bachmann, M.; Hoelzer, B.; Seibert, F.S.; Rohn, B.J.; Witzke, O.; Dittmer, U.; Brenner, T.; Paniskaki, K.; Yilmaz, S.; et al. Generation of Inhibitory Autoantibodies to ADAMTS13 in Coronavirus Disease 2019. medRxiv 2021. [Google Scholar] [CrossRef]
- Taha, M.; Samavati, L. Antiphospholipid antibodies in COVID-19: A meta-analysis and systematic review. RMD Open 2021, 7. [Google Scholar] [CrossRef]
- Pascolini, S.; Vannini, A.; Deleonardi, G.; Ciordinik, M.; Sensoli, A.; Carletti, I.; Veronesi, L.; Ricci, C.; Pronesti, A.; Mazzanti, L.; et al. COVID-19 and Immunological Dysregulation: Can Autoantibodies be Useful? Clin. Transl. Sci. 2021, 14, 502–508. [Google Scholar] [CrossRef]
- Sacchi, M.C.; Tamiazzo, S.; Stobbione, P.; Agatea, L.; De Gaspari, P.; Stecca, A.; Lauritano, E.C.; Roveta, A.; Tozzoli, R.; Guaschino, R.; et al. SARS-CoV-2 infection as a trigger of autoimmune response. Clin. Transl. Sci. 2020. [Google Scholar] [CrossRef]
- Zuo, Y.; Estes, S.K.; Ali, R.A.; Gandhi, A.A.; Yalavarthi, S.; Shi, H.; Sule, G.; Gockman, K.; Madison, J.A.; Zuo, M.; et al. Prothrombotic antiphospholipid antibodies in COVID-19. MedRxiv 2020. [Google Scholar] [CrossRef]
- Borghi, M.O.; Beltagy, A.; Garrafa, E.; Curreli, D.; Cecchini, G.; Bodio, C.; Grossi, C.; Blengino, S.; Tincani, A.; Franceschini, F.; et al. Anti-Phospholipid Antibodies in COVID-19 Are Different From Those Detectable in the Anti-Phospholipid Syndrome. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Gazzaruso, C.; Mariani, G.; Ravetto, C.; Malinverni, L.; Tondelli, E.; Cerrone, M.; Sala, V.; Bevilacqua, L.; Altavilla, T.; Coppola, A.; et al. Lupus anticoagulant and mortality in patients hospitalized for COVID-19. J. Thromb. Thrombolysis 2020, 1–7. [Google Scholar] [CrossRef]
- Vollmer, O.; Tacquard, C.; Dieudonné, Y.; Nespola, B.; Sattler, L.; Grunebaum, L.; Gies, V.; Radosavljevic, M.; Kaeuffer, C.; Hansmann, Y.; et al. Follow-up of COVID-19 patients: LA is transient but other aPLs are persistent. Autoimmun. Rev. 2021, 20, 102822. [Google Scholar] [CrossRef]
- Emmenegger, M.; Kumar, S.S.; Emmenegger, V.; Büttner, T.; Schierack, P.; Sprinzl, M.F.; Sommer, C.J.; Lackner, K.J.; Aguzzi, A.; Roggenbuck, D.; et al. Anti-prothrombin autoantibodies enriched after infection with SARS-CoV-2 and influenced by strength of antibody response against SARS-CoV-2 proteins. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zuniga, M.; Gomes, C.; Carsons, S.E.; Bender, M.T.; Cotzia, P.; Miao, Q.R.; Lee, D.C.; Rodriguez, A. Autoimmunity to the Lung Protective Phospholipid-Binding Protein Annexin A2 Predicts Mortality Among Hospitalized COVID-19 Patients. medRxiv 2021. [Google Scholar] [CrossRef]
- Pretorius, E.; Vlok, M.; Venter, C.; Bezuidenhout, J.A.; Laubscher, G.J.; Steenkamp, J.; Kell, D.B. Persistent clotting protein pathology in Long COVID/Post-Acute Sequelae of COVID-19 (PASC) is accompanied by increased levels of antiplasmin. medRxiv 2021. [Google Scholar] [CrossRef]
- Sarif, J.; Raychaudhuri, D.; D’Rozario, R.; Bandopadhyay, P.; Singh, P.; Mehta, P.; Hoque, M.A.; Sinha, B.P.; Kushwaha, M.; Sahni, S.; et al. Plasma gradient of soluble urokinase-type plasminogen activator receptor is linked to pathogenic plasma proteome and immune transcriptome and stratifies outcomes in severe COVID-19. medRxiv 2021. [Google Scholar] [CrossRef]
- Bye, A.P.; Hoepel, W.; Mitchell, J.L.; Jegouic, S.; Loureiro, S.; Sage, T.; de Taeye, S.; van Gils, M.; Kriek, N.; Cooper, N.; et al. Aberrant glycosylation of anti-SARS-CoV-2 IgG is a pro-thrombotic stimulus for platelets. Blood 2021. [Google Scholar] [CrossRef]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.-H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef]
- Focosi, D.; Franchini, M. Clinical predictors of SARS-CoV-2 neutralizing antibody titers in COVID-19 convalescents: Implications for convalescent plasma donor recruitment. Eur. J. Haematol. 2021, 107, 24–28. [Google Scholar] [CrossRef]
- Hensley, M.K.; Bain, W.G.; Jacobs, J.; Nambulli, S.; Parikh, U.; Cillo, A.; Staines, B.; Heaps, A.; Sobolewski, M.D.; Rennick, L.J.; et al. Intractable COVID-19 and Prolonged SARS-CoV-2 Replication in a CAR-T-cell Therapy Recipient: A Case Study. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2021. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Q.; Wang, Y.; Wang, G.; Wang, L.; Chen, H.; Jiao, T.; Hu, C.; Lei, X.; Guo, L.; et al. Analysis of the correlation between anti-MDA5 antibody and the severity of COVID-19: A retrospective study. medRxiv 2020. [Google Scholar] [CrossRef]
- Wilk, A.J.; Rustagi, A.; Zhao, N.Q.; Roque, J.; Martínez-Colón, G.J.; McKechnie, J.L.; Ivison, G.T.; Ranganath, T.; Vergara, R.; Hollis, T.; et al. A single-cell atlas of the peripheral immune response in patients with severe COVID-19. Nat. Med. 2020, 26, 1070–1076. [Google Scholar] [CrossRef]
- Gazzaruso, C.; Carlo Stella, N.; Mariani, G.; Nai, C.; Coppola, A.; Naldani, D.; Gallotti, P. High prevalence of antinuclear antibodies and lupus anticoagulant in patients hospitalized for SARS-CoV2 pneumonia. Clin. Rheumatol. 2020, 39, 2095–2097. [Google Scholar] [CrossRef]
- Woodruff, M.C.; Ramonell, R.P.; Lee, F.E.-H.; Sanz, I. Clinically identifiable autoreactivity is common in severe SARS-CoV-2 Infection. medRxiv 2020. [Google Scholar] [CrossRef]
- Gomes, C.; Zuniga, M.; Crotty, K.A.; Qian, K.; Hsu Lin, L.; Argyropoulos, K.; Li, H.; Cotzia, P.; Rodriguez, A. Autoimmune anti-DNA antibodies predict disease severity in COVID-19 patients. medRxiv 2021. [Google Scholar] [CrossRef]
- Casciola-Rosen, L.; Thiemann, D.R.; Andrade, F.; Trejo Zambrano, M.I.; Hooper, J.E.; Leonard, E.K.; Spangler, J.B.; Cox, A.L.; Machamer, C.E.; Sauer, L.; et al. IgM autoantibodies recognizing ACE2 are associated with severe COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Harrington, W.E.; Jiang, Y.; Duffy, F.; Hadlock, J.; Raappana, A.; Styrchak, S.; Beck, I.; Chour, W.; Houck, J.; Duvvuri, V.; et al. Angiotensin II receptor I auto-antibodies following SARS-CoV-2 infection. medRxiv 2021. [Google Scholar] [CrossRef]
- Frasca, D.; Reidy, L.; Romero, M.; Diaz, A.; Cray, C.; Kahl, K.; Blomberg, B. SARS-CoV-2 infection induces autoimmune antibody secretion more in lean than in obese COVID-19 patients. medRxiv 2021. [Google Scholar] [CrossRef]
- Franke, C.; Ferse, C.; Kreye, J.; Reincke, M.; Sanchez-Sendin, E.; Rocco, A.; Steinbrenner, M.; Angermair, S.; Treskatsch, S.; Zickler, D.; et al. High frequency of cerebrospinal fluid autoantibodies in COVID-19 patients with neurological symptoms. medRxiv 2020. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Focosi, D.; Franchini, M.; Pirofski, L.-a.; Burnouf, T.; Fairweather, D.; Joyner, M.J.; Casadevall, A. COVID-19 Convalescent Plasma Is More than Neutralizing Antibodies: A Narrative Review of Potential Beneficial and Detrimental Co-Factors. Viruses 2021, 13, 1594. https://doi.org/10.3390/v13081594
Focosi D, Franchini M, Pirofski L-a, Burnouf T, Fairweather D, Joyner MJ, Casadevall A. COVID-19 Convalescent Plasma Is More than Neutralizing Antibodies: A Narrative Review of Potential Beneficial and Detrimental Co-Factors. Viruses. 2021; 13(8):1594. https://doi.org/10.3390/v13081594
Chicago/Turabian StyleFocosi, Daniele, Massimo Franchini, Liise-anne Pirofski, Thierry Burnouf, DeLisa Fairweather, Michael J. Joyner, and Arturo Casadevall. 2021. "COVID-19 Convalescent Plasma Is More than Neutralizing Antibodies: A Narrative Review of Potential Beneficial and Detrimental Co-Factors" Viruses 13, no. 8: 1594. https://doi.org/10.3390/v13081594
APA StyleFocosi, D., Franchini, M., Pirofski, L.-a., Burnouf, T., Fairweather, D., Joyner, M. J., & Casadevall, A. (2021). COVID-19 Convalescent Plasma Is More than Neutralizing Antibodies: A Narrative Review of Potential Beneficial and Detrimental Co-Factors. Viruses, 13(8), 1594. https://doi.org/10.3390/v13081594