
Application of Plant Viruses in Biotechnology, Medicine, and Human Health

Abstract
:1. Introduction
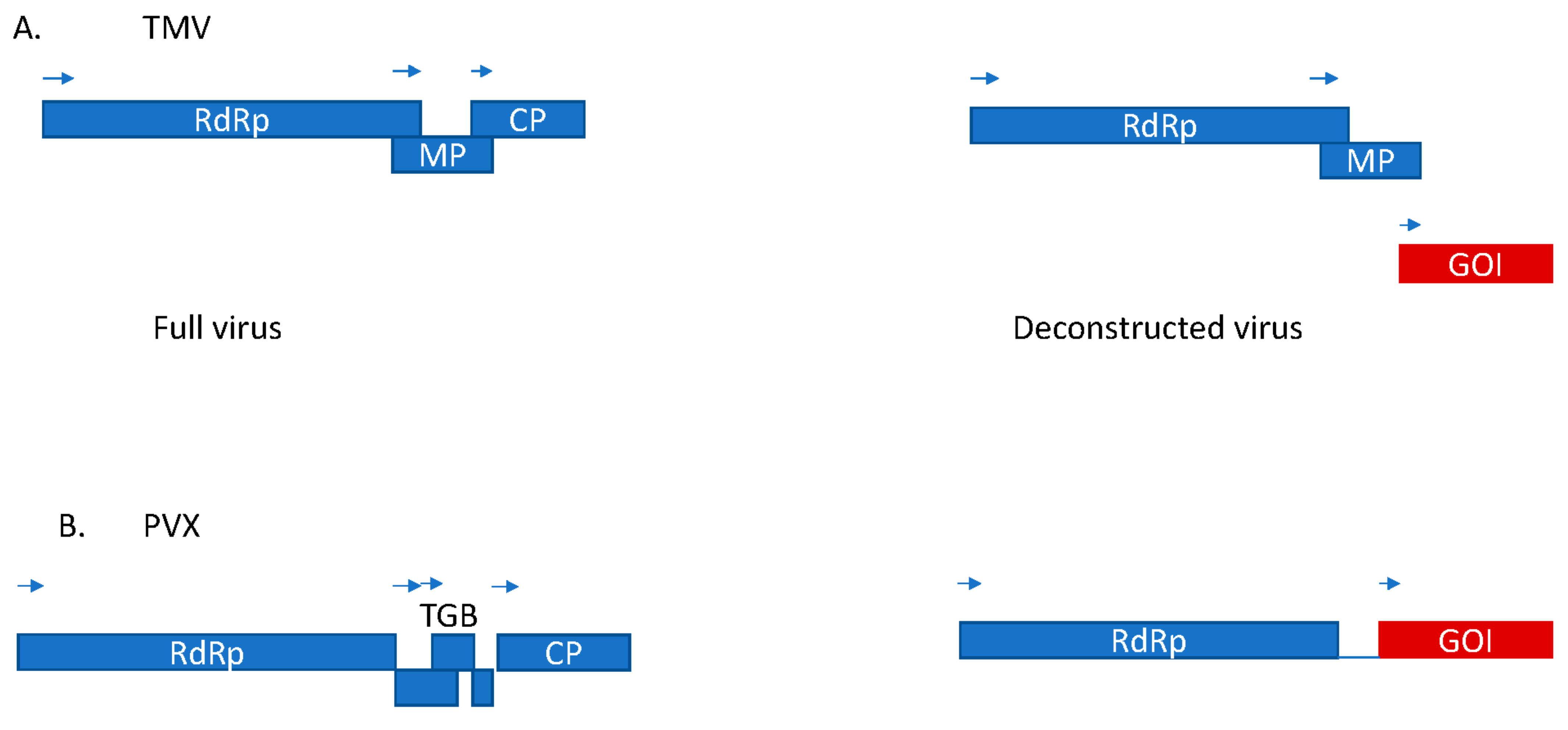
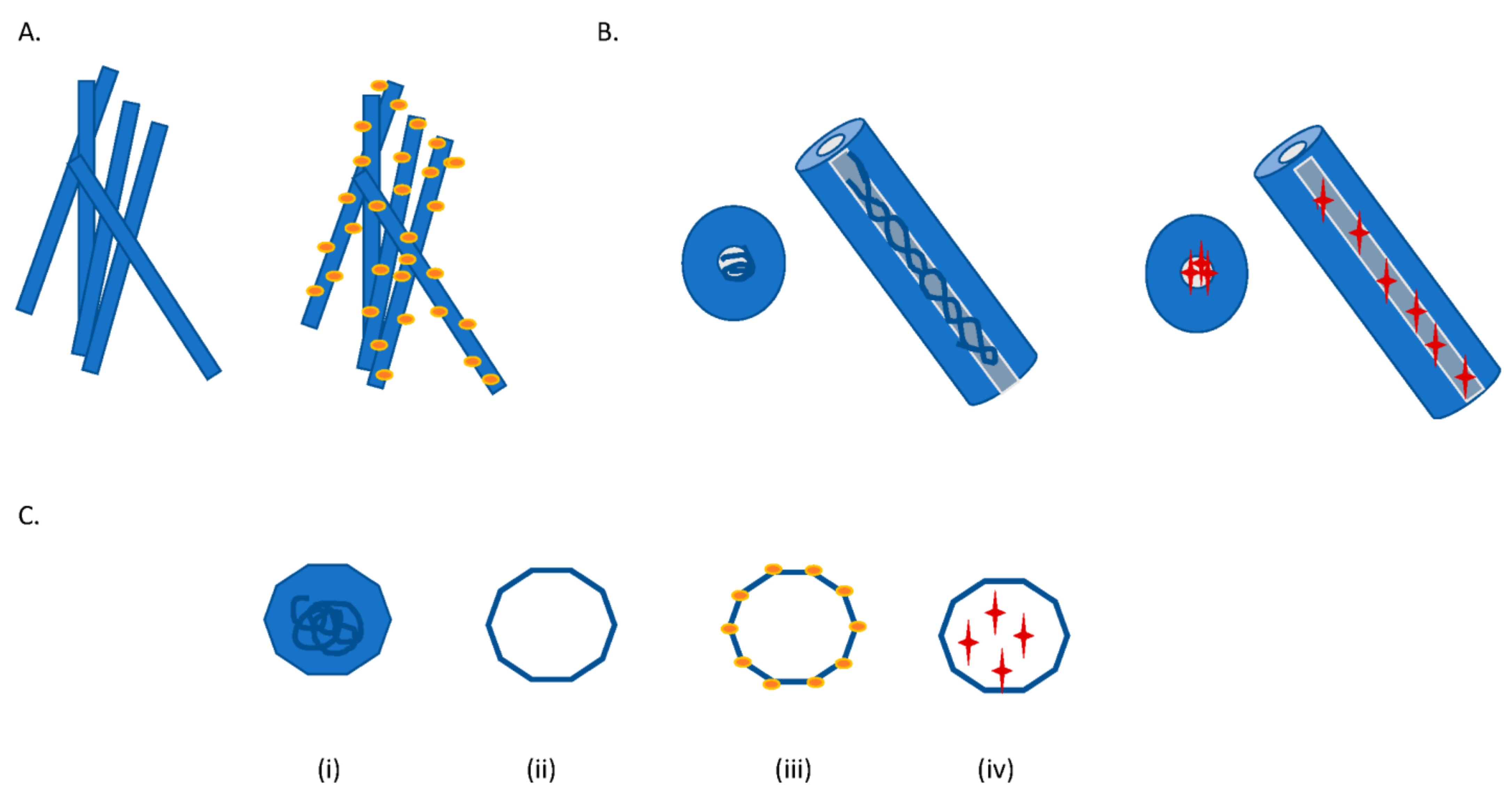
2. Molecular Characteristics of TMV Advantageous for Biotechnological Use
3. The Use of Genetically Engineered TMV in Biochemistry, Nanotechnology, and Plant Biotechnology
4. The Use of TMV in Medicine, Cancer, Imaging, and Theranostics
5. Molecular Characteristics of PVX Advantageous for Biotechnological Use
6. PVX as an Expression Vector and Repurposing PVX for Use in Medicine, Cancer, and Theranostics
7. Molecular Characteristics of CPMV Advantageous for Biotechnological Applications
8. Applications of Comoviruses CPMV and Cowpea Chlorotic Mottle Virus (CCMV) in Medical Biotechnology and Cancer
9. Molecular Features of Geminiviruses Advantageous for Biotechnological Use
10. The Use of Geminiviruses in Biotechnology and Medicine

{kind=link}
{kind=link}
| Virus | Application | References |
|---|---|---|
| Comovirus CPMV | Delays tumor growth using combination therapy | [137,138] |
| CPMV and cyclosposphamide | [140] | |
| Activation of monocytes, dendritic cells, macrophages | [141] | |
| Comovirus CCMV | mRNA vaccine delivery | [143] |
| Encapsulate CpG oligonucleotides, activated macrophages and inhibit growth | [144] | |
| Encapsulate drug-activating enymes to reduce side effects, increase targeting to tumor site | [145,146] | |
| Geminivirus BeYDV | Vaccines and monoclonal antibodies | [149] |
| Monoclonal antibodies to West Nile Virus, Ebola Virus | [149] | |
| RIC vaccines to ZIKV | [154,155] |
11. Viral Expression Vectors and the CRISPR/Cas9 Technology
12. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Acosta-Ramírez, E.; Pérez-Flores, R.; Majeau, N.; Pastelin-Palacios, R.; Gil-Cruz, C.; Ramírez-Saldaña, M.; Manjarrez-Orduño, N.; Cervantes-Barragan, L.; Santos-Argumedo, L.; Flores-Romo, L.; et al. Translating innate response into long-lasting antibody response by the intrinsic antigen-adjuvant properties of papaya mosaic virus. Immunology 2008, 124, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Yusibov, V.; Mett, V.; Davidson, C.; Musiychuk, K.; Gilliam, S.; Farese, A.; MacVittie, T.; Mann, D. Peptide-based candidate vaccine against respiratory syncytial virus. Vaccine 2005, 23, 2261–2265. [Google Scholar] [CrossRef] [PubMed]
- Kemnade, J.O.; Seethammagari, M.; Collinson-Pautz, M.; Kaur, H.; Spencer, D.M.; McCormick, A.A. Tobacco mosaic virus efficiently targets DC uptake, activation and antigen-specific T cell responses in vivo. Vaccine 2014, 32, 4228–4233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, F.R.; Bellaby, T.; Helliwell, S.M.; Jones, T.D.; Kamstrup, S.; Dalsgaard, K.; Flock, J.-I.; Hamilton, W.D.O. Chimeric plant virus particles administered nasally or orally induce systemic and mucosal immune responses in mice. J. Virol. 1999, 73, 930–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, F.; Jones, T.; Longstaff, M.; Chapman, S.; Bellaby, T.; Smith, H.; Xu, F.; Hamilton, W.; Flock, J.-I. Immunogenicity of peptides derived from a fibronectin-binding protein of S. aureus expressed on two different plant viruses. Vaccine 1999, 17, 1846–1857. [Google Scholar] [CrossRef]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Magnifection—A new platform for expressing recombinant vaccines in plants. Vaccine 2005, 23, 2042–2048. [Google Scholar] [CrossRef]
- Lindbo, J.A. TRBO: A High-Efficiency Tobacco Mosaic Virus RNA-Based Overexpression Vector. Plant Physiol. 2007, 145, 1232–1240. [Google Scholar] [CrossRef] [Green Version]
- Chapman, S.; Kavanagh, T.; Baulcombe, D. Potato virus X as a vector for gene expression in plants. Plant J. 1992, 2, 549–557. [Google Scholar]
- Bendandi, M.; Marillonnet, S.; Kandzia, R.; Thieme, F.; Nickstadt, A.; Herz, S.; Fröde, R.; Inogés, S.; de Cerio, A.L.-D.; Soria, E.; et al. Rapid, high-yield production in plants of individualized idiotype vaccines for non-Hodgkin’s lymphoma. Ann. Oncol. 2010, 21, 2420–2427. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, N.F. Viral nanoparticles as platforms for next-generation therapeutics and imaging devices. Nanomedicine 2010, 6, 634–641. [Google Scholar] [CrossRef] [Green Version]
- Young, M.; Debbie, W.; Uchida, M.; Douglas, T. Plant Viruses as Biotemplates for Materials and Their Use in Nanotechnology. Annu. Rev. Phytopathol. 2008, 46, 361–384. [Google Scholar] [CrossRef]
- Steinmetz, N.F.; Evans, D.J. Utilisation of plant viruses in bionanotechnology. Org. Biomol. Chem. 2007, 5, 2891–2902. [Google Scholar] [CrossRef]
- Chung, Y.H.; Cai, H.; Steinmetz, N.F. Viral nanoparticles for drug delivery, imaging, immunotherapy, and theranostic applications. Adv. Drug Deliv. Rev. 2020, 156, 214–235. [Google Scholar] [CrossRef] [PubMed]
- Namba, K.; Pattanayek, R.; Stubbs, G. Visualization of protein-nucleic acid interactions in a virus: Refined structure of intact tobacco mosaic virus at 2.9 Å resolution by X-ray fiber diffraction. J. Mol. Biol. 1989, 208, 307–325. [Google Scholar] [CrossRef]
- Parker, L.; Kendall, A.; Stubbs, G. Surface features of potato virus X from fiber diffraction. Virology 2002, 300, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Carette, N.; Engelkamp, H.; Akpa, E.; Pierre, S.J.; Cameron, N.R.; Christianen, P.C.M.; Maan, J.C.; Thies, J.C.; Weberskirch, R.; Rowan, A.E.; et al. A virus-based biocatalyst. Nat. Nanotechnol. 2007, 2, 226–229. [Google Scholar] [CrossRef]
- Yi, H.; Nisar, S.; Lee, S.-Y.; Powers, M.A.; Bentley, W.E.; Payne, G.F.; Ghodssi, R.; Rubloff, G.W.; Harris, M.T.; Culver, J.N. Patterned Assembly of Genetically Modified Viral Nanotemplates via Nucleic Acid Hybridization. Nano Lett. 2005, 5, 1931–1936. [Google Scholar] [CrossRef]
- Niehl, A.; Appaix, F.; Boscá, S.; Van Der Sanden, B.; Nicoud, J.-F.; Bolze, F.; Heinlein, M. Fluorescent Tobacco mosaic virus-Derived Bio-Nanoparticles for Intravital Two-Photon Imaging. Front. Plant Sci. 2016, 6, 1244. [Google Scholar] [CrossRef] [Green Version]
- Werner, S.; Marillonnet, S.; Hause, G.; Klimyuk, V.; Gleba, Y. Immunoabsorbent nanoparticles based on a tobamovirus displaying protein A. Proc. Natl. Acad. Sci. USA 2006, 103, 17678–17683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolenska, L.; Roberts, I.M.; Learmonth, D.; Porter, A.J.; Harris, W.J.; Wilson, T.; Cruz, S.S. Production of a functional single chain antibody attached to the surface of a plant virus. FEBS Lett. 1998, 441, 379–382. [Google Scholar] [CrossRef] [Green Version]
- Petukhova, N.; Gasanova, T.; Stepanova, L.; Rusova, O.; Potapchuk, M.; Korotkov, A.; Skurat, E.; Tsybalova, L.; Kiselev, O.; Ivanov, P.; et al. Immunogenicity and Protective Efficacy of Candidate Universal Influenza A Nanovaccines Produced in Plants by Tobacco Mosaic Virus-based Vectors. Curr. Pharm. Des. 2013, 19, 5587–5600. [Google Scholar] [CrossRef]
- Thérien, A.; Bédard, M.; Carignan, D.; Rioux, G.; Gauthier-Landry, L.; Laliberté-Gagné, M.-È.; Bolduc, M.; Savard, P.; Leclerc, D. A versatile papaya mosaic virus (PapMV) vaccine platform based on sortase-mediated antigen coupling. J. Nanobiotechnol. 2017, 15, 54. [Google Scholar] [CrossRef] [Green Version]
- Tseng, R.J.; Tsai, C.; Ma, L.; Ouyang, J.; Ozkan, C.S.; Yang, Y. Digital memory device based on tobacco mosaic virus conjugated with nanoparticles. Nat. Nanotechnol. 2006, 1, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Venuti, A.; Curzio, G.; Mariani, L.; Paolini, F. Immunotherapy of HPV-associated cancer: DNA/plant-derived vaccines and new orthotopic mouse models. Cancer Immunol. Immunother. 2015, 64, 1329–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Sánchez, L.; Cadena-Nava, R.D.; Palomares, L.A.; Ruiz-Garcia, J.; Koay, M.S.; Cornelissen, J.J.; Vazquez-Duhalt, R. Chemotherapy pro-drug activation by biocatalytic virus-like nanoparticles containing cytochrome P450. Enzym. Microb. Technol. 2014, 60, 24–31. [Google Scholar] [CrossRef]
- Phelps, J.P.; Dang, N.; Rasochova, L. Inactivation and purification of cowpea mosaic virus-like particles displaying peptide antigens from Bacillus anthracis. J. Virol. Methods 2007, 141, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azizgolshani, O.; Garmann, R.F.; Cadena-Nava, R.; Knobler, C.M.; Gelbart, W.M. Reconstituted plant viral capsids can release genes to mammalian cells. Virology 2013, 441, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Eiben, S.; Koch, C.; Altintoprak, K.; Southan, A.; Tovar, G.; Laschat, S.; Weiss, I.M.; Wege, C. Plant virus-based materials for biomedical applications: Trends and prospects. Adv. Drug Deliv. Rev. 2019, 145, 96–118. [Google Scholar] [CrossRef]
- Lomonossoff, G.P.; Wege, C. TMV Particles: The Journey from Fundamental Studies to Bionanotechnology Applications. Adv. Virus Res. 2018, 102, 149. [Google Scholar]
- Gamper, C.; Spenlé, C.; Boscá, S.; Van Der Heyden, M.; Erhardt, M.; Orend, G.; Bagnard, D.; Heinlein, M. Functionalized Tobacco Mosaic Virus Coat Protein Monomers and Oligomers as Nanocarriers for Anti-Cancer Peptides. Cancers 2019, 11, 1609. [Google Scholar] [CrossRef] [Green Version]
- Scholthof, K.-B.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- Lomonossoff, G.P. So what have plant viruses ever done for virology and molecular biology? Adv. Virus Res. 2018, 100, 145. [Google Scholar] [PubMed]
- Gallie, D.R.; Sleat, D.E.; Watts, J.W.; Turner, P.C.; Wilson, T.M.A. The 5′-leader sequence of tobacco mosaic virus RNA enhances the expression of foreign gene transcripts in vitro and in vivo. Nucleic Acids Res. 1987, 15, 3257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, T.M.A. Plant viruses: A tool-box for genetic engineering and crop protection. Bioessays 1989, 10, 179. [Google Scholar] [PubMed]
- Peyret, H.; Lomonossoff, G.P. When plant virology met Agrobacterium: The rise of the deconstructed clones. Plant Biotechnol. J. 2015, 13, 1121–1135. [Google Scholar] [CrossRef] [Green Version]
- Golemboski, D.B.; Lomonossoff, G.P.; Zaitlin, M. Plants transformed with a tobacco mosaic virus nonstructural gene sequence are resistant to the virus. Proc. Natl. Acad. Sci. USA 1990, 87, 6311–6315. [Google Scholar] [CrossRef] [Green Version]
- Abel, P.P.; Nelson, R.S.; De, B.; Hoffmann, N.; Rogers, S.G.; Fraley, R.T.; Beachy, R.N. Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene. Science 1986, 232, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Fromm, S.; Bharat, T.A.; Jakobi, A.J.; Hagen, W.; Sachse, C. Seeing tobacco mosaic virus through direct electron detectors. J. Struct. Biol. 2015, 189, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Sleat, D.; Turner, P.; Finch, J.; Butler, P.; Wilson, T. Packaging of recombinant RNA molecules into pseudovirus particles directed by the origin-of-assembly sequence from tobacco mosaic virus RNA. Virology 1986, 155, 299–308. [Google Scholar] [CrossRef]
- Bruckman, M.; Hern, S.; Jiang, K.; Flask, C.A.; Yu, X.; Steinmetz, N.F. Tobacco mosaic virus rods and spheres as supramolecular high-relaxivity MRI contrast agents. J. Mater. Chem. B 2013, 1, 1482–1490. [Google Scholar] [CrossRef] [Green Version]
- Fraenkel-Conrat, H.; Williams, R.C. Reconstitution of active tobacco mosaic virus from its inactive protein and nucleic acid components. Proc. Natl. Acad. Sci. USA 1955, 41, 690–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.L.; Fitzmaurice, W.P.; Turpen, T.H.; Palmer, K.E. Display of Peptides on the Surface of Tobacco Mosaic Virus Particles. Curr. Top. Microbiol. Immunol. 2009, 332, 13–31. [Google Scholar] [CrossRef]
- Moore, L.; Hamorsky, K.; Matoba, N. Production of Recombinant Cholera Toxin B Subunit in Nicotiana benthamiana Using GENEWARE® Tobacco Mosaic Virus Vector. Methods Protoc. 2016, 1385, 129–137. [Google Scholar] [CrossRef]
- McCormick, A.A.; Reinl, S.J.; Cameron, T.I.; Vojdani, F.; Fronefield, M.; Levy, R.; Tusé, D. Individualized human scFv vaccines produced in plants: Humoral anti-idiotype responses in vaccinated mice confirm relevance to the tumor Ig. J. Immunol. Methods 2003, 278, 95–104. [Google Scholar] [CrossRef]
- Kumagai, M.H.; Donson, J.; Dellacioppa, G.R.; Grill, L.K. Rapid, high-level expression of glycosylated rice α-amylase in transfected plants by an RNA viral vector. Gene 2000, 245, 169–174. [Google Scholar] [CrossRef]
- Giritch, A.; Marillonnet, S.; Engler, C.; van Eldik, G.; Botterman, J.; Klimyuk, V.; Gleba, Y. Rapid high-yield expression of full-size IgG antibodies in plants coinfected with noncompeting viral vectors. Proc. Natl. Acad. Sci. USA 2006, 103, 14701–14706. [Google Scholar] [CrossRef] [Green Version]
- Gils, M.; Kandzia, R.; Marillonnet, S.; Klimyuk, V.; Gleba, Y. High-yield production of authentic human growth hormone using a plant virus-based expression system. Plant Biotechnol. J. 2005, 3, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Czapar, A.E.; Zheng, Y.; Riddell, I.A.; Shukla, S.; Awuah, S.G.; Lippard, S.J.; Steinmetz, N.F. Tobacco Mosaic Virus Delivery of Phenanthriplatin for Cancer therapy. ACS Nano 2016, 10, 4119–4126. [Google Scholar] [CrossRef] [Green Version]
- Sainsbury, F.; Lomonossoff, G.P. Extremely High-Level and Rapid Transient Protein Production in Plants without the Use of Viral Replication. Plant Physiol. 2008, 148, 1212–1218. [Google Scholar] [CrossRef] [Green Version]
- Porta, C.; Spall, V.E.; Loveland, J.; Johnson, J.E.; Barker, P.J.; Lomonossoff, G.P. Development of Cowpea Mosaic Virus as a High-Yielding System for the Presentation of Foreign Peptides. Virology 1994, 202, 949–955. [Google Scholar] [CrossRef]
- Aljabali, A.A.A.; Sainsbury, F.; Lomonossoff, G.P.; Evans, D.J. Cowpea Mosaic Virus Unmodified Empty Viruslike Particles Loaded with Metal and Metal Oxide. Small 2010, 6, 818–821. [Google Scholar] [CrossRef]
- Uhde-Holzem, K.; Schlösser, V.; Viazov, S.; Fischer, R.; Commandeur, U. Immunogenic properties of chimeric potato virus X particles displaying the hepatitis C virus hypervariable region I peptide R9. J. Virol. Methods 2010, 166, 12–20. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, Q.; Hjelm, B.; Arntzen, C.; Mason, H. A DNA replicon system for rapid high-level production of virus-like particles in plants. Biotechnol. Bioeng. 2009, 103, 706–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, A.; Odon, V.; Kormelink, R. Plant Viruses in Plant Molecular Pharming: Toward the Use of Enveloped Viruses. Front. Plant Sci. 2019, 10, 803. [Google Scholar] [CrossRef] [PubMed]
- Finbloom, J.A.; Han, K.; Aanei, I.L.; Hartman, E.C.; Finley, D.T.; Dedeo, M.T.; Fishman, M.; Downing, K.H.; Francis, M.B. Stable Disk Assemblies of a Tobacco Mosaic Virus Mutant as Nanoscale Scaffolds for Applications in Drug Delivery. Bioconj. Chem. 2016, 27, 2480–2485. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Eber, F.J.; Nagarajan, A.S.; DiFranco, N.A.; Schmidt, N.; Wen, A.M.; Eiben, S.; Twyman, R.M.; Wege, C.; Steinmetz, N.F. The Impact of Aspect Ratio on the Biodistribution and Tumor Homing of Rigid Soft-Matter Nanorods. Adv. Healthc. Mater. 2015, 4, 874–882. [Google Scholar] [CrossRef] [Green Version]
- Bazzini, A.A.; Hopp, H.E.; Beachy, R.N.; Asurmendi, S. Infection and coaccumulation of tobacco mosaic virus proteins alter microRNA levels, correlating with symptom and plant development. Proc. Natl. Acad. Sci. USA 2007, 104, 12157–12162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, S.; Dickmeis, C.; Nagarajan, A.S.; Fischer, R.; Commandeur, U.; Steinmetz, N.F. Molecular farming of fluorescent virus-based nanoparticles for optical imaging in plants, human cells and mouse models. Biomater. Sci. 2014, 2, 784–797. [Google Scholar] [CrossRef]
- Wang, Q.; Kaltgrad, E.; Lin, T.; Johnson, J.; Finn, M. Natural Supramolecular Building Blocks: Wild-Type Cowpea Mosaic Virus. Chem. Biol. 2002, 9, 805–811. [Google Scholar] [CrossRef] [Green Version]
- Huynh, N.T.; Hesketh, E.L.; Saxena, P.; Meshcheriakova, Y.; Ku, Y.-C.; Hoang, L.T.; Johnson, J.E.; Ranson, N.; Lomonossoff, G.P.; Reddy, V.S. Crystal Structure and Proteomics Analysis of Empty Virus-like Particles of Cowpea Mosaic Virus. Structure 2016, 24, 567–575. [Google Scholar] [CrossRef] [Green Version]
- Sokullu, E.; Abyaneh, H.S.; Gauthier, M.A. Plant/Bacterial Virus-Based Drug Discovery, Drug Delivery, and Therapeutics. Pharmaceutics 2019, 11, 211. [Google Scholar] [CrossRef] [Green Version]
- Koch, C.; Poghossian, A.; Schöning, M.J.; Wege, C. Penicillin Detection by Tobacco Mosaic Virus-Assisted Colorimetric Biosensors. Nanotheranostics 2018, 2, 184–196. [Google Scholar] [CrossRef] [Green Version]
- Poghossian, A.; Jablonski, M.; Koch, C.; Bronder, T.S.; Rolka, D.; Wege, C.; Schoning, M.J. Field-effect biosensor using virus particles as scaffolds for enzyme immobilization. Biosens. Bioelectron. 2018, 110, 168. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.; Wabbel, K.; Eber, F.J.; Krolla-Sidenstein, P.; Azucena, C.; Gliemann, H.; Eiben, S.; Geiger, F.; Wege, C. Modified TMV particles as beneficial scaffolds to present sensor enzymes. Front. Plant Sci. 2015, 6, 1137. [Google Scholar] [CrossRef]
- Bäcker, M.; Koch, C.; Eiben, S.; Geiger, F.; Eber, F.; Gliemann, H.; Poghossian, A.; Wege, C.; Schoening, M.J. Tobacco mosaic virus as enzyme nanocarrier for electrochemical biosensors. Sens. Actuators B Chem. 2017, 238, 716–722. [Google Scholar] [CrossRef]
- Tiu, B.D.B.; Kernan, D.L.; Tiu, S.B.; Wen, A.M.; Zheng, Y.; Pokorski, J.K.; Advincula, R.C.; Steinmetz, N.F. Electrostatic layer-by-layer construction offibrous TMV biofilms. Nanoscale 2017, 9, 1580. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Wang, C.; Sun, J.; Wang, Q. The synergistic effects of multivalent ligand display and nanotopography on osteogenic differentiation of rat bone marrow stem cells. Biomaterials 2010, 31, 5813–5824. [Google Scholar] [CrossRef] [PubMed]
- Sitasuwan, P.; Lee, L.A.; Li, K.; Nguyen, H.G.; Wang, Q. RGD-conjugated rod-like viral nanoparticles on 2D scaffold improve bone differentiation of mesenchymal stem cells. Front. Chem. 2014, 2, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zang, J.; Lee, L.A.; Niu, Z.; Horvatha, G.C.; Braxtona, V.; Wibowo, A.C.; Bruckman, M.A.; Ghoshroy, S.; Loye, H.-C.Z.; et al. Electrospinning fabrication, structural and mechanical characterization of rod-like virus-based composite nanofibers. J. Mater. Chem. 2011, 21, 8550–8557. [Google Scholar] [CrossRef] [Green Version]
- Beachy, R.N. Coat–protein–mediated resistance to tobacco mosaic virus: Discovery mechanisms and exploitation. Philos. Trans. R. Soc. B Biol. Sci. 1999, 354, 659–664. [Google Scholar] [CrossRef] [Green Version]
- Mascia, T.; Nigro, F.; Abdallah, A.; Ferrara, M.; De Stradis, A.; Faedda, R.; Palukaitis, P.; Gallitelli, D. Gene silencing and gene expression in phytopathogenic fungi using a plant virus vector. Proc. Natl. Acad. Sci. USA 2014, 111, 4291–4296. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Ablack, A.L.; Wen, A.M.; Lee, K.L.; Lewis, J.D.; Steinmetz, N.F. Increased Tumor Homing and Tissue Penetration of the Filamentous Plant Viral Nanoparticle Potato virus X. Mol. Pharm. 2013, 10, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.L.; Hubbard, L.C.; Hern, S.; Yildiz, I.; Gratzl, M.; Steinmetz, N.F. Shape matters: The diffusion rates of TMV rods and CPMV icosahedrons in a spheroid model of extracellular matrix are distinct. Biomater. Sci. 2013, 1, 581–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, J.; Niu, Z.; Lee, L.A.; Wang, Q. Self-assembly of viral particles. Curr. Opin. Colloid Interface Sci. 2011, 16, 441–450. [Google Scholar] [CrossRef]
- Park, J.; Gao, H.; Wang, Y.; Hu, H.; Simon, D.I.; Steinmetz, N.F. S100A9-targeted tobacco mosaic virus nanoparticles exhibit high specificity toward atherosclerotic lesions in ApoE−/− mice. J. Mater. Chem. B 2019, 7, 1842–1846. [Google Scholar] [CrossRef]
- Grasso, S.; Santi, L. Viral nanoparticles as macromolecular devices for new therapeutic and pharmaceutical approaches. Int. J. Physiol. Pathophysiol. Pharmacol. 2010, 2, 161–178. [Google Scholar]
- Chapman, S.; Faulkner, C.; Kaiserli, E.; Garcia-Mata, C.; Savenkov, E.I.; Roberts, A.G.; Oparka, K.J.; Christie, J.M. The photoreversible fluorescent protein iLOV outperforms GFP as a reporter of plant virus infection. Proc. Natl. Acad. Sci. USA 2008, 105, 20038–20043. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Jiang, L.; Zhou, Z.; Fan, J.; Zhang, Q.; Zhu, H.; Han, Q.; Xu, Z. Expression of foot-and-mouth disease virus epitopes in tobacco by a tobacco mosaic virus-based vector. Vaccine 2003, 21, 4390. [Google Scholar] [CrossRef]
- Koo, M.; Bendahmane, M.; Lettieri, G.A.; Paoletti, A.D.; Lane, T.E.; Fitchen, J.H.; Buchmeier, M.J.; Beachy, R.N. Protective immunity against murine hepatitis virus (MHV) induced by intranasal or subcutaneous administration of hybrids of tobacco mosaic virus that carries an MHV epitope. Proc. Natl. Acad. Sci. USA 1999, 96, 7774–7779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staczek, J.; Bendahmane, M.; Gilleland, L.B.; Beachy, R.N.; Gilleland, H. Immunization with a chimeric tobacco mosaic virus containing an epitope of outer membrane protein F of Pseudomonas aeruginosa provides protection against challenge with P. aeruginosa. Vaccine 2000, 18, 2266–2274. [Google Scholar] [CrossRef]
- Pitek, A.S.; Park, J.; Wang, Y.; Gao, H.; Hu, H.; Simon, D.I.; Steinmetz, N.F. Delivery of thrombolytic therapy using rod-shaped plant viral nanoparticles decreases the risk of hemorrhage. Nanoscale 2018, 10, 16547–16555. [Google Scholar] [CrossRef]
- Yin, Z.; Nguyen, H.G.; Chowdhury, S.; Bentley, P.; Bruckman, M.A.; Miermont, A.; Gildersleeve, J.C.; Wang, Q.; Huang, X. Tobacco mosaic virus as a new carrier for tumor associated carbohydrate antigens. Bioconjug. Chem. 2012, 23, 1694–1703. [Google Scholar] [CrossRef] [Green Version]
- Finbloom, J.A.; Aanei, I.L.; Bernard, J.M.; Klass, S.H.; Elledge, S.K.; Han, K.; Ozawa, T.; Nicolaides, T.P.; Berger, M.S.; Francis, M.B. Evaluation of Three Morphologically Distinct Virus-Like Particles as Nanocarriers for Convection-Enhanced Drug Delivery to Glioblastoma. Nanomaterials 2018, 8, 1007. [Google Scholar] [CrossRef] [Green Version]
- Pitek, A.S.; Hu, H.; Shukla, S.; Steinmetz, N.F. Cancer Theranostic Applications of Albumin-Coated Tobacco Mosaic Virus Nanoparticles. ACS Appl. Mater. Interfaces 2018, 10, 39468–39477. [Google Scholar] [CrossRef]
- Franke, C.E.; Czapar, A.E.; Patel, R.; Steinmetz, N.F. Tobacco Mosaic Virus-Delivered Cisplatin Restores Efficacy in Platinum-Resistant Ovarian Cancer Cells. Mol. Pharm. 2017, 15, 2922–2931. [Google Scholar] [CrossRef]
- Vernekar, A.; Berger, G.; Czapar, A.E.; Veliz, F.A.; Wang, D.I.; Steinmetz, N.F.; Lippard, S.J. Speciation of Phenanthriplatin and Its Analogs in the Core of Tobacco Mosaic Virus. J. Am. Chem. Soc. 2018, 140, 4279–4287. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Liu, B.; Gao, S.; Wang, Z.; Tian, Y.; Wu, M.; Jiang, S.; Niu, Z. Glyco-decorated tobacco mosaic virus as a vector for cisplatin delivery. J. Mater. Chem. B 2017, 5, 2078–2085. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Liu, X.; Wang, Z.; Jiang, S.; Wu, M.; Tian, Y.; Niu, Z. Fluorous interaction induced self-assembly of tobacco mosaic virus coat protein for cisplatin delivery. Nanoscale 2018, 10, 11732–11736. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.D.; Steinmetz, N.F. Tobacco mosaic virus delivery of mitoxantrone for cancer therapy. Nanoscale 2018, 10, 16307–16313. [Google Scholar] [CrossRef] [PubMed]
- Kernan, D.L.; Wen, A.M.; Pitek, A.S.; Steinmetz, N.F. Featured Article: Delivery of chemotherapeutic vcMMAE using tobacco mosaic virus nanoparticles. Exp. Biol. Med. 2017, 242, 1405–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Zhou, M.; Shi, H.; Gao, S.; Xie, G.; Zhu, M.; Wu, M.; Chen, J.; Niu, Z. Integration of Cell-Penetrating Peptides with Rod-like Bionanoparticles: Virus-Inspired Gene-Silencing Technology. Nano Lett. 2018, 18, 5453–5460. [Google Scholar] [CrossRef] [PubMed]
- Chariou, P.L.; Wang, L.; Desai, C.; Park, J.; Robbins, L.K.; von Recum, H.A.; Ghiladi, R.A.; Steinmetz, N.F. Let There Be Light: Targeted Photodynamic Therapy Using High Aspect Ratio Plant Viral Nanoparticles. Macromol. Biosci. 2019, 19, e1800407. [Google Scholar] [CrossRef] [PubMed]
- Luzuriaga, M.A.; Welch, R.P.; Dharmarwardana, M.; Benjamin, C.E.; Li, S.; Shahrivarkevishahi, A.; Popal, S.; Tuong, L.H.; Creswell, C.T.; Gassensmith, J.J. Enhanced Stability and Controlled Delivery of MOF-Encapsulated Vaccines and Their Immunogenic Response In Vivo. ACS Appl. Mater. Interfaces 2019, 11, 9740–9746. [Google Scholar] [CrossRef] [PubMed]
- Bruckman, M.; Jiang, K.; Simpson, E.J.; Randolph, L.N.; Luyt, L.; Yu, X.; Steinmetz, N.F. Dual-Modal Magnetic Resonance and Fluorescence Imaging of Atherosclerotic Plaques in Vivo Using VCAM-1 Targeted Tobacco Mosaic Virus. Nano Lett. 2014, 14, 1551–1558. [Google Scholar] [CrossRef]
- Dharmarwardana, M.; Martins, A.F.; Chen, Z.; Palacios, P.M.; Nowak, C.M.; Welch, R.P.; Li, S.; Luzuriaga, M.A.; Bleris, L.; Pierce, B.S.; et al. Nitroxyl Modified Tobacco Mosaic Virus as a Metal-Free High-Relaxivity MRI and EPR Active Superoxide Sensor. Mol. Pharm. 2018, 15, 2973–2983. [Google Scholar] [CrossRef]
- Lee, H.; Shahrivarkevishahi, A.; Lumata, J.L.; Luzuriaga, M.A.; Hagge, L.M.; Benjamin, C.E.; Brohlin, O.R.; Parish, C.R.; Firouzi, H.R.; Nielsen, S.O.; et al. Supramolecular and biomacromolecular enhancement of metal-free magnetic resonance imaging contrast agents. Chem. Sci. 2020, 11, 2045–2050. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Black, K.C.L.; Luehmann, H.; Li, W.; Zhang, Y.S.; Cai, X.; Wan, D.; Liu, S.-Y.; Li, M.; Kim, P.; et al. Comparison Study of Gold Nanohexapods, Nanorods, and Nanocages for Photothermal Cancer Treatment. ACS Nano 2013, 7, 2068–2077. [Google Scholar] [CrossRef] [PubMed]
- Vankayala, R.; Huang, Y.-K.; Kalluru, P.; Chiang, C.-S.; Hwang, K.C. First Demonstration of Gold Nanorods-Mediated Photodynamic Therapeutic Destruction of Tumors via Near Infra-Red Light Activation. Small 2014, 10, 1612–1622. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.-B.G. Tobaccomosaic Virus: A Model System for Plant Biology. Annu. Rev. Phytopathol. 2004, 42, 13–34. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Yang, Q.; Baroni, S.; Yang, H.; Aime, S.; Steinmetz, N.F. Polydopamine-decorated tobacco mosaic virus for photoacoustic/magnetic resonance bimodal imaging and photothermal cancer therapy. Nanoscale 2019, 11, 9760–9768. [Google Scholar] [CrossRef]
- Yu, X.-Q.; Jia, J.-L.; Zhang, C.-L.; Li, X.-D.; Wang, Y.-J. Phylogenetic analyses of an isolate obtained from potato in 1985 revealed potato virus X was introduced to China via multiple events. Virus Genes 2010, 40, 447–451. [Google Scholar] [CrossRef]
- Wang, Y.; Cong, Q.-Q.; Lan, Y.-F.; Geng, C.; Li, X.-D.; Liang, Y.-C.; Yang, Z.-Y.; Zhu, X.-P.; Li, X.-D. Development of new potato virus X-based vectors for gene over-expression and gene silencing assay. Virus Res. 2014, 191, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Lacomme, C.; Chapman, S. Use of potato virus X (PVX)-based vectors for geneexpression and virus-induced gene silencing (VIGS). Curr. Protoc. Microbiol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Plchova, H.; Moravec, T.; Hoffmeisterova, H.; Folwarczna, J.; Čeřovská, N. Expression of Human papillomavirus 16 E7ggg oncoprotein on N- and C-terminus of Potato virus X coat protein in bacterial and plant cells. Protein Expr. Purif. 2011, 77, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Hefferon, K. Plant Virus Expression Vectors: A Powerhouse for Global Health. Biomedicines 2017, 5, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhde-Holzem, K.; McBurney, M.; Tiu, B.D.; Advincula, R.C.; Fischer, R.; Commandeur, U.; Steinmetz, N.F. Production of Immunoabsorbent Nanoparticles by Displaying Single-Domain Protein A on Potato Virus X. Macromol. Biosci. 2016, 16, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Biswas, M.E.; Chen, P. Study of Binding between Protein A and Immunoglobulin G Using a Surface Tension Probe. Biophys. J. 2003, 84, 509–522. [Google Scholar] [CrossRef] [Green Version]
- Arkhipenko, M.V.; Petrova, E.K.; Nikitin, N.A.; Protopopova, A.D.; Dubrovin, E.V.; Yaminskii, I.V.; Rodionova, N.P.; Karpova, O.; Atabekov, J.G.; Center, A.T. Characteristics of Artificial Virus-like Particles Assembled in vitro from Potato Virus X Coat Protein and Foreign Viral RNAs. Acta Nat. 2011, 3. [Google Scholar] [CrossRef]
- Tyulkina, L.G.; Skurat, E.V.; Frolova, O.Y.; Komarova, T.V.; Karger, E.M.; Atabekov, I.G. New Viral Vector for Superproduction of Epitopes of Vaccine Proteins in Plants. Acta Nat. 2011, 3, 11. [Google Scholar] [CrossRef]
- Shukla, S.; DiFranco, N.A.; Wen, A.M.; Commandeur, U.; Steinmetz, N.F. To Target or Not to Target: Active vs. Passive Tumor Homing of Filamentous Nanoparticles Based on Potato virus X. Cell. Mol. Bioeng. 2015, 8, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Röder, J.; Dickmeis, C.; Fischer, R.; Commandeur, U. Systemic infection of nicotiana benthamianawith potato virus X nanoparticles presenting a fluorescent iLOV polypeptide fused directly to the coat protein. Biomed. Res. Int. 2018, e932867. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Dickmeis, C.; Fischer, R.; Commandeur, U.; Steinmetz, N.F. In Planta Production of Fluorescent Filamentous Plant Virus-Based Nanoparticles. Methods Mol. Biol. 2018, 1776, 61–84. [Google Scholar] [CrossRef]
- Li, P.; Jing, C.; Ren, H.; Jia, Z.; Ghanem, H.; Wu, G.; Li, M.; Qing, L.; Li, P.; Jing, C.; et al. Analysis of Pathogenicity and Virulence Factors of Ageratum leaf curl Sichuan virus. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Shi, J.; Zhu, Y.; Li, M.; Ma, Y.; Liu, H.; Zhang, P.; Fang, D.; Guo, Y.; Xu, P.; Qiao, Y. Establishment of a novel virus-induced virulence effector assay for the identification of virulence effectors of plant pathogens using a PVX-based expression vector. Mol. Plant Pathol. 2020, 21, 1654–1661. [Google Scholar] [CrossRef]
- Wang, X.; Luo, C.; Xu, Y.; Zhang, C.; Bao, M.; Dou, J.; Wang, Q.; Cheng, Y. Expression of the p24 silencing suppressor of Grapevine leafroll-associated virus 2 from Potato virus X or Barley stripe mosaic virus vector elicits hypersensitive responses in Nicotiana benthamiana. Plant Physiol Biochem. 2019, 142, 34–42. [Google Scholar] [CrossRef]
- Wieczorek, P.; Wrzesińska, B.; Frąckowiak, P.; Przybylska, A.; Obrępalska-Stęplowska, A. Contribution of Tomato torrado virus Vp26 coat protein subunit to systemic necrosis induction and virus infectivity in Solanum lycopersicum. Virol. J. 2019, 16, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faivre-Rampant, O.; Gilroy, E.M.; Hrubikova, K.; Hein, I.; Millam, S.; Loake, G.J.; Birch, P.; Taylor, M.; Lacomme, C. Potato virus X-induced gene silencing in leavesand tubers of potato. Plant Physiol. 2004, 134, 1308–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopertekh, L.; Meyer, T.; Freyer, C.; Hust, M. Transient plant production of Salmonella Typhimurium diagnostic antibodies. Biotechnol. Rep. 2018, 20, e00314. [Google Scholar] [CrossRef]
- Demurtas, O.C.; Massa, S.; Illiano, E.; De Martinis, D.; Chan, P.; Di Bonito, P.; Franconi, R. Antigen Production in Plant to Tackle Infectious Diseases Flare Up: The Case of SARS. Front. Plant Sci. 2016, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Mardanova, E.S.; Kotlyarov, R.Y.; Kuprianov, V.V.; Stepanova, L.A.; Tsybalova, L.M.; Lomonosoff, G.P.; Ravin, N.V. Rapid high-yield expression of a candidate influenza vaccine based on the ectodomain of M2 protein linked to flagellin in plants using viral vectors. BMC Biotechnol. 2015, 15, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadzadeh, S.; Roohvand, F.; Memarnejadian, A.; Jafari, A.; Ajdary, S.; Salmanian, A.H.; Ehsani, P. Co-expression of hepatitis C virus polytope-HBsAg and p19-silencing suppressor protein in tobacco leaves. Pharm. Biol. 2016, 54, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Khezri, G.; Rouz, B.B.K.; Ofoghi, H.; Davarpanah, S.J. Heterologous expression of biologically active Mambalgin-1 peptide as a new potential anticancer, using a PVX-based viral vector in Nicotiana benthamiana. Plant Cell Tissue Organ Cult. 2020, 142, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Esfandiari, N.; Arzanani, M.K.; Soleimani, M.; Kohi-Habibi, M.; Svendsen, W.E. A new application of plant virus nanoparticles as drug delivery in breast cancer. Tumor Biol. 2016, 37, 1229–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demurtas, O.C.; Massa, S.; Ferrante, P.; Venuti, A.; Franconi, R.; Giuliano, G. A Chlamydomonas-Derived Human Papillomavirus 16 E7 Vaccine Induces Specific Tumor Protection. PLoS ONE 2013, 8, e61473. [Google Scholar] [CrossRef] [Green Version]
- Le, D.H.T.; Lee, K.L.; Shukla, S.; Commandeur, U.; Steinmetz, N.F. Potato virus X, a filamentous plant viral nanoparticle for doxorubicin delivery in cancer therapy. Nanoscale 2017, 9, 2348–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.L.; Murray, A.A.; Le, D.H.T.; Sheen, M.R.; Shukla, S.; Commandeur, U.; Fiering, S.; Steinmetz, N.F. Combination of Plant Virus Nanoparticle-Based in Situ Vaccination with Chemotherapy Potentiates Antitumor Response. Nano Lett. 2017, 17, 4019–4028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, D.H.T.; Commandeur, U.; Steinmetz, N.F. Presentation and Delivery of Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand via Elongated Plant Viral Nanoparticle Enhances Antitumor Efficacy. ACS Nano 2019, 13, 2501–2510. [Google Scholar] [CrossRef]
- Jobsri, J.; Allen, A.; Rajagopal, D.; Shipton, M.; Kanyuka, K.; Lomonossoff, G.P.; Ottensmeier, C.; Diebold, S.S.; Stevenson, F.; Savelyeva, N. Plant Virus Particles Carrying Tumour Antigen Activate TLR7 and Induce High Levels of Protective Antibody. PLoS ONE 2015, 10, e0118096. [Google Scholar] [CrossRef] [PubMed]
- Kruse, I.; Peyret, H.; Saxena, P.; Lomonossoff, G.P. Encapsidation of Viral RNA in Picornavirales: Studies on Cowpea Mosaic Virus Demonstrate Dependence on Viral Replication. J. Virol. 2019, 93, e01520-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Cañizares, M.; Monger, W.; Perrin, Y.; Tsakiris, E.; Porta, C.; Shariat, N.; Nicholson, L.; Lomonossoff, G.P. Cowpea mosaic virus-based systems for the production of antigens and antibodies in plants. Vaccine 2005, 23, 1788–1792. [Google Scholar] [CrossRef]
- Sainsbury, F.; Cañizares, M.C.; Lomonossoff, G.P. Cowpea mosaicVirus: The Plant Virus–Based Biotechnology Workhorse. Annu. Rev. Phytopathol. 2010, 48, 437–455. [Google Scholar] [CrossRef] [Green Version]
- Sainsbury, F.; Sack, M.; Stadlmann, J.; Quendler, H.; Fischer, R.; Lomonossoff, G.P. Rapid Transient Production in Plants by Replicating and Non-Replicating Vectors Yields High Quality Functional Anti-HIV Antibody. PLoS ONE 2010, 5, e13976. [Google Scholar] [CrossRef] [Green Version]
- Meshcheriakova, Y.; Durrant, A.; Hesketh, E.L.; Ranson, N.; Lomonossoff, G.P. Combining high-resolution cryo-electron microscopy and mutagenesis to develop cowpea mosaic virus for bionanotechnology. Biochem. Soc. Trans. 2017, 45, 1263–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Beiss, V.; Steinmetz, N.F. Cowpea Mosaic Virus Nanoparticles and Empty Virus-Like Particles Show Distinct but Overlapping Immunostimulatory Properties. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Dent, M.; Matoba, N. Cancer biologics made in plants. Curr. Opin. Biotechnol. 2020, 61, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, N.F.; Cho, C.-F.; Ablack, A.; Lewis, J.D.; Manchester, M. Cowpea mosaic virus nanoparticles target surface vimentin on cancer cells. Nanomedicine 2011, 6, 351–364. [Google Scholar] [CrossRef] [Green Version]
- Lizotte, P.; Wen, A.M.; Sheen, M.R.; Fields, J.; Rojanasopondist, P.; Steinmetz, N.F.; Fiering, S. In situ vaccination with cowpea mosaic virus nanoparticles suppresses metastatic cancer. Nat. Nanotechnol. 2016, 11, 295–303. [Google Scholar] [CrossRef]
- Patel, R.; Czapar, A.E.; Fiering, S.; Oleinick, N.L.; Steinmetz, N.F. Radiation Therapy Combined with Cowpea Mosaic Virus Nanoparticle in Situ Vaccination Initiates Immune-Mediated Tumor Regression. ACS Omega 2018, 3, 3702–3707. [Google Scholar] [CrossRef]
- Wang, C.; Steinmetz, N.F. CD47 Blockade and Cowpea Mosaic Virus Nanoparticle In Situ Vaccination Triggers Phagocytosis and Tumor Killing. Adv. Healthc. Mater. 2019, 8, e1801288. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Wang, C.; Shukla, S.; Steinmetz, N.F. Cowpea Mosaic Virus Immunotherapy Combined with Cyclophosphamide Reduces Breast Cancer Tumor Burden and Inhibits Lung Metastasis. Adv. Sci. 2019, 6, 1802281. [Google Scholar] [CrossRef] [Green Version]
- Albakri, M.; Veliz, F.A.; Fiering, S.N.; Steinmetz, N.F.; Sieg, S.F. Endosomal toll-like receptors play a key role in activation of primary human monocytes by cowpea mosaic virus. Immunology 2019, 159, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Wang, C.; Beiss, V.; Cai, H.; Washington, T., II; Murray, A.A.; Gong, X.; Zhao, Z.; Masarapu, H.; Zlotnick, A.; et al. The unique potency of Cowpea mosaic virus (CPMV) in situ cancer vaccine. Biomater. Sci. 2020, 8, 5489–5503. [Google Scholar] [CrossRef] [PubMed]
- Biddlecome, A.; Habte, H.H.; McGrath, K.M.; Sambanthamoorthy, S.; Wurm, M.; Sykora, M.M.; Knobler, C.M.; Lorenz, I.C.; Lasaro, M.; Elbers, K.; et al. Delivery of self-amplifying RNA vaccines in in vitro reconstituted virus-like particles. PLoS ONE 2019, 14, e0215031. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Shukla, S.; Steinmetz, N.F. The Antitumor Efficacy of CpG Oligonucleotides is Improved by Encapsulation in Plant Virus-Like Particles. Adv. Funct. Mater. 2020, 30, 1908743. [Google Scholar] [CrossRef] [PubMed]
- Koyani, R.D.; Pérez-Robles, J.; Cadena-Nava, R.D.; Vazquez-Duhalt, R. Biomaterial-based nanoreactors, an alternative for enzyme delivery. Nanotechnol. Rev. 2017, 6, 405–419. [Google Scholar] [CrossRef]
- Strable, E.; Finn, M.G. Chemical Modification of Viruses and Virus-Like Particles. In Viruses and Nanotechnology; Manchester, M., Steinmetz, N.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–21. [Google Scholar] [CrossRef]
- Diamos, A.G.; Mason, H.S. High-level expression and enrichment of norovirus virus-like particles in plants using modified geminiviral vectors. Protein Expr. Purif. 2018, 151, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E.P.; Martin, D.P. Virus-Derived ssDNA Vectors for the Expression of Foreign Proteins in Plants. Curr. Top. Microbiol. Immunol. 2011, 375, 19–45. [Google Scholar] [CrossRef]
- Hefferon, K.L. DNA Virus Vectors for Vaccine Production in Plants: Spotlight on Geminiviruses. Vaccines 2014, 2, 642–653. [Google Scholar] [CrossRef]
- Dickey, A.; Wang, N.; Cooper, E.; Tull, L.; Breedlove, D.; Mason, H.; Liu, D.; Wang, K.Y. Transient Expression of Lumbrokinase (PI239) in Tobacco (Nicotiana tabacum) Using a Geminivirus-Based Single Replicon System Dissolves Fibrin and Blood Clots. Evid. Based Complement. Altern. Med. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Bañuelos-Hernández, B.; Monreal-Escalante, E.; Gonzalez-Ortega, O.; Angulo, C.; Rosales-Mendoza, S. Algevir: An Expression System for Microalgae Based on Viral Vectors. Front. Microbiol. 2017, 8, 1100. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Durán, R. Geminiviruses for biotechnology: The art of parasite taming. New Phytol. 2016, 210, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.-Y.; Ding, B.; Zhou, X.-P. Geminiviruses and their application in biotechnology. J. Integr. Agric. 2017, 16, 2761–2771. [Google Scholar] [CrossRef]
- Diamos, A.G.; Pardhe, M.D.; Sun, H.; Hunter, J.G.L.; Kilbourne, J.; Chen, Q.; Mason, H.S. A Highly Expressing, Soluble, and Stable Plant-Made IgG Fusion Vaccine Strategy Enhances Antigen Immunogenicity in Mice Without Adjuvant. Front. Immunol. 2020, 11, 576012. [Google Scholar] [CrossRef] [PubMed]
- Diamos, A.G.; Pardhe, M.D.; Sun, H.; Hunter, J.G.; Mor, T.; Meador, L.; Kilbourne, J.; Chen, Q.; Mason, H.S. Codelivery of improved immune complex and virus-like particle vaccines containing Zika virus envelope domain III synergistically enhances immunogenicity. Vaccine 2020, 38, 3455–3463. [Google Scholar] [CrossRef]
- Čermák, T.; Curtin, S.J.; Gil-Humanes, J.; Čegan, R.; Kono, T.J.Y.; Konečná, E.; Belanto, J.J.; Starker, C.G.; Mathre, J.W.; Greenstein, R.L.; et al. A Multipurpose Toolkit to Enable Advanced Genome Engineering in Plants. Plant Cell. 2017, 29, 1196–1217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zheng, Q.; Yi, X.; An, H.; Zhao, Y.; Ma, S.; Zhou, G. Establishing RNA virus resistance in plants by harnessing CRISPR immune system. Plant Biotechnol. J. 2018, 16, 1415–1423. [Google Scholar] [CrossRef] [Green Version]
- Ariga, H.; Toki, S.; Ishibashi, K. Potato Virus X Vector-Mediated DNA-Free Genome Editing in Plants. Plant Cell Physiol. 2020, 61, 1946–1953. [Google Scholar] [CrossRef]
- Ludman, M.; Burgyán, J.; Fátyol, K. Crispr/Cas9 Mediated Inactivation of Argonaute 2 Reveals its Differential Involvement in Antiviral Responses. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Brisson, N.; Paszkowski, J.; Penswick, J.R.; Gronenborn, B.; Potrykus, I.; Hohn, T.M. Expression of a bacterial gene in plants by using a viral vector. Nat. Cell Biol. 1984, 310, 511–514. [Google Scholar] [CrossRef]
- French, R.; Janda, M.; Ahlquist, P. Bacterial Gene Inserted in an Engineered RNA Virus: Efficient Expression in Monocotyledonous Plant Cells. Science 1986, 231, 1294–1297. [Google Scholar] [CrossRef]
- Palmer, K.; Gleba, Y. Plant Viral Vectors. Curr. Top. Microbiol. Immunol. 2014, 375, 1–194. [Google Scholar] [CrossRef]
- Baulcombe, D.; Chapman, S.; Cruz, S.S. Jellyfish green fluorescent protein as a reporter for virus infections. Plant J. 1995, 7, 1045–1053. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Hunter, D.; Voogd, C.; Joyce, N.; Davies, K. A Narcissus mosaic viral vector system for protein expression and flavonoid production. Plant Methods 2013, 9, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takamatsu, N.; Ishikawa, M.; Meshi, T.; Okada, Y. Expression of bacterial chloramphenicol acetyltransferase gene in tobacco plants mediated by TMV-RNA. EMBO J. 1987, 6, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Yusibov, V.; Shivprasad, S.; Turpen, T.H.; Dawson, W.; Koprowski, H. Plant Viral Vectors Based on Tobamoviruses. Plant Biotechnol. 2000, 240, 81–94. [Google Scholar] [CrossRef]
- Jarugula, S.; Charlesworth, S.R.; Qu, F.; Stewart, L.R. Soil-borne wheat mosaic virus infectious clone and manipulation for gene-carrying capacity. Arch. Virol. 2016, 161, 2291–2297. [Google Scholar] [CrossRef] [PubMed]
- Lellis, A.D.; Kasschau, K.D.; Whitham, S.A.; Carrington, J.C. Loss-ofsusceptibility mutants of Arabidopsis thaliana reveal an essential role for eIF (iso)4E during potyvirus infection. Curr. Biol. 2002, 12, 1046–1051. [Google Scholar] [CrossRef] [Green Version]
- Majer, E.; Navarro, J.-A.; Daròs, J.-A. A potyvirus vector efficiently targets recombinant proteins to chloroplasts, mitochondria and nuclei in plant cells when expressed at the amino terminus of the polyprotein. Biotechnol. J. 2015, 10, 1792–1802. [Google Scholar] [CrossRef]
- Seo, J.-K.; Choi, H.-S.; Kim, K.-H. Engineering of soybean mosaic virus as a versatile tool for studying protein–protein interactions in soybean. Sci. Rep. 2016, 6, 22436. [Google Scholar] [CrossRef] [Green Version]
- Stanley, J. Geminiviruses: Plant viral vectors. Curr. Opin. Genet. Dev. 1993, 3, 91–96. [Google Scholar] [CrossRef]
- Sainsbury, F.; Lavoie, P.O.; D’Aoust, M.A.; Vezina, L.P.; Lomonossoff, G.P. Expression of multiple proteins using full-length and deleted versions of Cowpea mosaic virus RNA-2. Plant Biotechnol. J. 2008, 6, 82–92. [Google Scholar] [CrossRef]
- Zhang, C.; Bradshaw, J.D.; Whitham, S.A.; Hill, J.H. The Development of an Efficient Multipurpose Bean Pod Mottle Virus Viral Vector Set for Foreign Gene Expression and RNA Silencing. Plant Physiol. 2010, 153, 52–65. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Zhang, Y.; Wang, X.-B.; Dong, J.; Wang, B.; Han, C.; Yu, J.; Li, D. Oral administration of plant-based rotavirus VP6 induces antigen-specific IgAs, IgGs and passive protection in mice. Vaccine 2010, 28, 6021–6027. [Google Scholar] [CrossRef]
- Bouton, C.; King, R.C.; Chen, H.; Azhakanandam, K.; Bieri, S.; Hammond-Kosack, K.E.; Kanyuka, K. Foxtail mosaic virus: A Viral Vector for Protein Expression in Cereals. Plant Physiol. 2018, 177, 1352–1367. [Google Scholar] [CrossRef] [Green Version]
- Cheuk, A.; Houde, M. A New Barley Stripe Mosaic Virus Allows Large Protein Overexpression for Rapid Function Analysis. Plant Physiol. 2018, 176, 1919–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.-S.; Hammond-Kosack, K.; Kanyuka, K. Barley Stripe Mosaic Virus-Mediated Tools for Investigating Gene Function in Cereal Plants and Their Pathogens: Virus-Induced Gene Silencing, Host-Mediated Gene Silencing, and Virus-Mediated Overexpression of Heterologous Protein. Plant Physiol. 2012, 160, 582–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Li, C.; Yan, L.; Jackson, A.O.; Liu, Z.; Han, C.; Yu, J.; Li, D. A High Throughput Barley Stripe Mosaic Virus Vector for Virus Induced Gene Silencing in Monocots and Dicots. PLoS ONE 2011, 6, e26468. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.-R.; Stenger, D.C.; Morris, T.J.; French, R. A plant virus vector for systemic expression of foreign genes in cereals. Plant J. 2000, 23, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Elzoghby, A.; Samy, W.; Elgindy, N.A. Protein-based nanocarriers as promising drug and gene delivery systems. J. Control. Release 2012, 161, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Nolte, R.; Cornelissen, J.J. Virus-based nanocarriers for drug delivery. Adv. Drug Deliv. Rev. 2012, 64, 811–825. [Google Scholar] [CrossRef]
- Ferrer-Miralles, N.; Rodríguez-Carmona, E.; Corchero, J.L.; García-Fruitós, E.; Vázquez, E.; Villaverde, A. Engineering protein self-assembling in protein-based nanomedicines for drug delivery and gene therapy. Crit. Rev. Biotechnol. 2015, 35, 209–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Recombinant Protein or Vaccine or VLP | Viral Vector |
|---|---|
| Cholera toxin b subunit | TMV [43] |
| Human anti-non-Hodgkin’s lymphoma single-chain Fv (scFv) immunoglobulins | Hybrid TMV and odontoglossum ringspot virus (ORSV) [44] |
| Rice a-amylase | Hybrid TMV and tomato mosaic virus (ToMV) [45] |
| Assembled full-size monoclonal antibody | Combination of non-competing viral vectors TMV and PVX [46] |
| Human growth hormone | Hybrid crucifer-infecting TMV (cr-TMV) and turnip vein-clearing virus (TVCV) [47] |
| Plant-produced VLP developed for drug delivery | TMV [48] |
| Plant-produced chimaeric virus vaccine for influenza virus | TMV [21] |
| Assembled full-size monoclonal antibody | CPMV [49] |
| Plant-produced chimaeric virus vaccine for human rhinovirus 14 and human immunodeficiency virus | CPMV [50] |
| Plant-produced VLP developed for encapsulation of metals | CPMV [51] |
| Plant-produced chimaeric virus vaccine for hepatitis C virus | PVX [52] |
| Hepatitis B core Norwalk virus capsid protein (NVCP) | BeYDV [53] |
| Virus | Symmetry | Family | Locations within the CP Amenable to Genetic Modification |
|---|---|---|---|
| TMV | Rod-like | Tombusviridae | Threonine 104/158, serine 123, N/C-terminal of coat protein [55,56,57] |
| PVX | Rod-like | Potexviridae | N-terminal of coat protein [58] |
| CPMV | Icosahedral | Comoviridae | βB-βC loop of the small subunit/βE-βF loop of the large subunit [59,60] |
| (a) Applications of TMV in medicine | ||
| Engineered Modifications | Effects | Reference |
| The extreme C-terminus of the TMV CP fused to the 11 amino acid epitope of the foot and mouth disease virus (FMDV) VP1 protein | This nanoparticle protected animals against FMDV challenge | [78] |
| Peptides from the coronavirus murine hepatitis virus spike protein displayed on the surface of TMV particles | Increased antibody titers and protected mice against murine hepatitis virus challenge | [79] |
| An epitope from Pseudomonas aeruginosa outer membrane protein F fused to the C-terminus of the TMV CP | Demonstrated immunity to Pseudomonas aeruginosa | [80] |
| The influenza virus M2e epitope displayed by fusion near the C-terminus of the TMV CP | Afforded protective anti-influenza immune response in mice | [21] |
| TMV conjugated to the thrombolytic tissue plasminogen activator tPA | Functioned efficiently equivalent to free tPA and enhanced safety profile as shown by diminished average bleeding times and therefore applicable for cardiovascular therapy | [81] |
| (b) Applications of TMV in cancer treatment | ||
| Engineered Modifications | Effects | Reference |
| TMV employed to display a weakly immunogenic tumor-associated carbohydrate antigen, the Tn antigen (GalNAc-α-O-Ser/Thr) | Potent immune responses were observed when the Tn antigen was conjugated to Tyr 139 of TMV | [82] |
| TMV CP used as nanocarrier for a highly hydrophobic, insoluble peptide that binds to the neuropilin (NRP1) receptor transmembrane domain in cancer cells | Shown to be anti-angiogenic by reducing cancer cell growth and migration | [30] |
| Doxorubicin (DOX) loaded onto TMV disks | Increased rates of survival of mice bearing intracranial glioblastoma | [83] |
| DOX loaded onto TMV VNPs coated with albumin | Antitumor effects | [84] |
| Cisplatin and phenanthriplatin loaded into the cavity of TMV by formation of stable covalent adduct or by charge-based reaction | Enhanced absorption by cancer cells and improved cytotoxicity | [85,86] |
| TMV VNPs loaded with cisplatin modified using lactose and mannose moieties on their external surface | This construction assisted the VNP’s recognition by the asialoglycoprotein receptor that is present on cell membranes and demonstrated augmented cytotoxicity in cancer cell lines | [87] |
| Modification of the TMV coat protein with a molecular fluorous ponytail incorporated at specific sites which resulted in self-assembly of the virus into spherical VNPs | These spherical VNP’s conferred greater stability of for the cisplatin-VNP complexes formed via metal-ligated coordination | [88] |
| Mitoxanthrone (MTO) loaded onto TMV VNPs by a charge-driven mechanism | Increased antitumor effects in mice | [89] |
| Antimitotic drug, valine-citrulline monomethyl auristatin E loaded onto external surface of TMV VNPs | Effective targeting and cytotoxicity in non-Hodgkin’s lymphoma cell line, Karpas 299; internal entry of TMV VNPs into endolysosomal components accompanied by protease-encoded release of the drug | [90] |
| Transacting activation transduction (TAT) peptide fused to the external surface of TMV | The engineered TAT-tagged TMV was internalized; this delivered RNA silencing in nude mice hepatocellular carcinoma tumors upon intravenous and intratumoral delivery | [91] |
| Zn-EpPor (5-(4-ethynylphenyl)-10,15,20-tris(4-methylpyridin-4-ium-1-yl)porphyrin-zinc(II) triiodide), a photosensitizer drug loaded onto the interior of the TMV particles | Demonstrated high stability and shelf-life; drug was released into endolysosomes and showed augmented cell-killing efficiency | [61] |
| Zn-Por+3 loaded TMV conjugated to F3 peptide | Targeted the nucleolin shuttle protein overexpressed on Hela cells; drug accumulated on cell membranes along with increased cell-killing efficiency likely due to disruption of the cell membrane through light activation followed by drug release and cellular uptake | [92] |
| (c) Applications of TMV in theranostics | ||
| Engineered Modifications | Effects | Reference |
| A near infrared fluorescent (NIR) dye as well as a peptide targeting S100A9 (a myeloid-related protein 14 present in atherosclerotic lesions and a molecular marker for acute myocardial infarctions) were conjugated to TMV | These targeted TMV particles were able to identify atherosclerotic lesions in apolipoprotein E-deficient (ApoE-/-) mice upon intravenous injection, showing that TMV can be used as a platform to detect at-risk lesions | [75] |
| A TMV-MOF (metal-organic framework) hybrid nanoparticle engineered | Increased retention of the TMV VNPs observed in mice | [93] |
| A Cy5-encapsulated TMV coated with zeolitic imidazolate framework-8 (Cy5-TMV@ZIF) | Improved the fluorescence retention time by 2.5 times more than that of the Cy5-TMV alone; this TMV@ZIF was recalcitrant to harsh conditions and proved to be highly stable and non-toxic | [93] |
| Gd-dodecane tetraacetic acid (Gd-DOTA) loaded onto TMV particles altered to target the vascular cell adhesion molecule, VCAM-1 | Facilitated the sensitive identification and depiction of atherosclerotic plaques in ApoE-/- mice, using low doses of the contrast agent wherein the augmented relaxivity and slower tumbling of the Gd-DOTA coupled with the TMV carrier improved the signal-to-noise ratio; also, this coupling afforded greater sensitivity of imaging, allowing 40× decrease in Gd dose in comparison with the standard clinical doses | [94] |
| Packing of a dysprosium (Dy3+) complex within the interior cavity of TMV | Enhanced T2 relaxivity towards MRI; this enabled NIR fluorescent dye delivery, which facilitated dual optical-MR imaging. The exterior surface of TMV was labeled with an Asp-Gly-Glu-Ala peptide that enabled target specificity to integrin α2β1 molecules on prostate cancer cells | [13] |
| A metal-free paramagnetic nitroxide organic radical contrast agent (ORCA) loaded onto TMV particles to generate electron paramagnetic resonance and MRI probes towards the detection of superoxide | This augmented in vitro r1 and r2 relaxivities and these probes worked as both T1 as well as T2 contrast agents, facilitating their suitability for preclinical and clinical MRI scanning | [95] |
| TMV conjugated to a derivative of the aminoxyl radical TEMPO (tetramethylpiperidin-1-oxyl, coined Compound 6) by means of a copper catalyzed azide-alkyne cyclo-addition reaction | Subsequent interaction with cucurbit [8] uril (CB [8]) generated an aminoxyl-based ORCA (semitroxane) that was silent for MRI; the r1 (relaxivity) values for TMV-6 emulated that of Gd-DOTA | [96] |
| TMV nanorods loaded with Gd and coated with polydopamine (PDA) | The PDA enhanced the MRI properties and provided PDA contrast, while simultaneously facilitating photothermal therapy (PTT); strong in vitro NIR absorption was observed along with increased photothermal conversion efficiency, compared to that of gold nanocages [97] and nanorods [98]; also, these VNPs demonstrated potent efficiency with lowered cytotoxicity in treating 4T1 breast and PC-3 prostate cancer cells in vitro | [99,100] |
| (a) Applications of PVX in identifying pathogenicity determinants and in VIGS | ||
| Engineered Modifications | Effects | Reference |
| PVX used as an expression vector for the production of V2, C1, and C4 proteins of a novel monopartite begomovirus, the Ageratum leaf curl Sichuan virus in N. benthamiana | Deletion and mutational analysis of the C4 protein using this PVX-derived vector showed that C4 is the major pathogenicity determinant which impacted symptom expression and virus accumulation | [113] |
| Phytophthora sojae virulence effector Avh148 expressed in plants using a PVX-based vector and a virus-induced virulence effector (VIVE) assay to detect putative effectors encoded by various plant pathogens | This PVX-Avh148 vector infected plants with strong viral symptoms and led to elevated levels of Avh148 effector and viral RNA accumulation; Avh148 was found to be essential for full pathogenic virulence; this VIVE assay could detect putative effectors encoded by various plant pathogens including even unculturable pathogens using this PVX-based expression vector | [114] |
| Grapevine leafroll-associated virus 2 (GLRaV-2) encodes a p24 polypeptide (a suppressor of RNA-silencing) that was expressed in a PVX-based vector | p24 causes systemic necrosis in N. benthamiana wherein a cytoplasmic Zn2+-binding protein, NbRAR1 is involved and the symptoms are characteristic of a hypersensitive response; the essential role of p24 in GLRaV-2 pathogenesis was elucidated using the PVX expression vector wherein both silencing suppression and p24 self-interaction are critical for the pathogenic activity of p24 | [115] |
| Tomato torrado virus (ToTV) capsid protein subunits Vp23, Vp26, and Vp35 expressed transiently from a PVX-derived vector in Solanum lycopersicum | Of these, Vp26 protein was shown to be the necrosis and pathogenicity determinant responsible for severe systemic necrosis of the plants accompanied by increased ribonuclease and oxidative activities | [116] |
| PVX has been developed as a VIGS vector in potatoes wherein VIGS mediates silencing of endogenous plant genes, thus helping to investigate the functions of the silenced genes | This caused the silencing of the endogenous phytoene desaturase gene in potato plants which led to characteristic photobleaching symptoms in the leaves by interference of the carotenoid biosynthetic pathway | [117] |
| (b) Applications of PVX in the diagnosis, prophylaxis, and therapy of infectious diseases | ||
| Engineered Modifications | Effects | Reference |
| The scFv-TM43-E10 and scFv-Fc-TM43-E10 antibody derivatives specific for the recognition of the Salmonella typhimurium Omp D protein expressed in a deconstructed PVX vector deficient for virus movement | These PVX vector-based antibodies exhibited similar antigen-binding specificities as that of their mammalian/microbial cell-generated counterparts and were able to successfully recognize the S. typhimurium Omp D antigen; therefore showed great promise as new diagnostic tools for the detection of S. typhimurium infection | [118] |
| The Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) N and M proteins expressed using PVX | The presence of antibodies specific to the SARS-CoV N protein could be detected in SARS-CoV patient sera using the plant-derived N protein | [119] |
| The M2e peptide of H1N1 Influenza virus was fused to bacterial flagellin to augment immunogenicity and then expressed in a PVX vector | The yield of the fusion protein was as high as 30% of the total soluble protein and mice inoculated with the PVX-derived protein exhibited protection against Influenza virus infection | [120] |
| The hyper variable region 1 (HVR-1) epitope of Hepatitis C Virus (HCV) expressed in a PVX Vector and administered parenterally | This elicited IgG immune response and the PVX-HVR1 epitope reacted positively with the serum of chronic HCV patients | [53] |
| A second capsid protein promoter of PVX used to express a chimaeric protein derived from fusion of the HCV core antigen with the hepatitis B virus (HBV) surface antigen (HBsAg) | This PVX-based polytopic HCVpc-HBsAg construct could be a potential plant-derived HCV vaccine | [121] |
| (c).Applications of PVX in cancer | ||
| Engineered Modifications | Effects | Reference |
| PVX used as an expression vector for Mambalgin-1, a peptide that functions as a potent analgesic by obstructing acid-sensing ion channels (ASIC) in nerve cells wherein the ASIC is involved in the growth and proliferation of cancer cells | This resulted in the production of Mambalgin-1 which exhibited cytotoxicity towards nervous (SH-SY5Y) cancer cells, inhibited ASIC channels and potentiated anticancer effects | [122] |
| Monoclonal antibodies of Herceptin or Trastuzumab loaded onto PVX nanofilaments | This successfully induced apoptosis in breast cancer cell lines | [123] |
| PVX used as an expression vector for a mutant form of the HPV16 E7 oncoprotein, by fusing it with lichenase | This elicited protection against tumor progression in mice by inducing robust cytotoxic T-cell response | [124] |
| The filamentous PVX used to deliver DOX | These DOX-loaded PVX VNPs greatly diminished the growth of tumors in athymic mice harboring breast cancer xenografts | [125] |
| PVX-DOX combination | Prolonged mouse survival and stimulated chemokine/cytokine levels in mouse intradermal melanoma models | [126] |
| PVX used to display tumor necrosis factor-related apoptosis inducing ligand (TRAIL) | Multivalent display of TRAIL enabled increased recruitment and stimulation of death receptors expressed on cancer cell lines and successfully suppressed tumor growth in mice breast cancer models | [127] |
| PVX conjugated to an idiotypic (Id) tumor-associated antigen (TAA) recombinant through a biotin/streptavidin linker | This elicited a 7 times higher anti-Id IgG response as compared with Id alone in a mouse B-cell lymphoma model; IFN-α and IL-12 were induced; also TLR7 was found to be essential for viral RNA recognition | [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venkataraman, S.; Hefferon, K. Application of Plant Viruses in Biotechnology, Medicine, and Human Health. Viruses 2021, 13, 1697. https://doi.org/10.3390/v13091697
Venkataraman S, Hefferon K. Application of Plant Viruses in Biotechnology, Medicine, and Human Health. Viruses. 2021; 13(9):1697. https://doi.org/10.3390/v13091697
Chicago/Turabian StyleVenkataraman, Srividhya, and Kathleen Hefferon. 2021. "Application of Plant Viruses in Biotechnology, Medicine, and Human Health" Viruses 13, no. 9: 1697. https://doi.org/10.3390/v13091697
APA StyleVenkataraman, S., & Hefferon, K. (2021). Application of Plant Viruses in Biotechnology, Medicine, and Human Health. Viruses, 13(9), 1697. https://doi.org/10.3390/v13091697