
Rhinovirus Inhibitors: Including a New Target, the Viral RNA


Abstract
:1. Introduction
2. Targeting the Viral Capsid
3. Targeting Non-Structural Viral Proteins
4. The Rhinoviral RNA
5. RNA Encapsidation
6. RNA Uncoating
7. Access of Intercalating Compounds Is Limited by the Compactness of the RNA Core
8. Targeting the Viral RNA
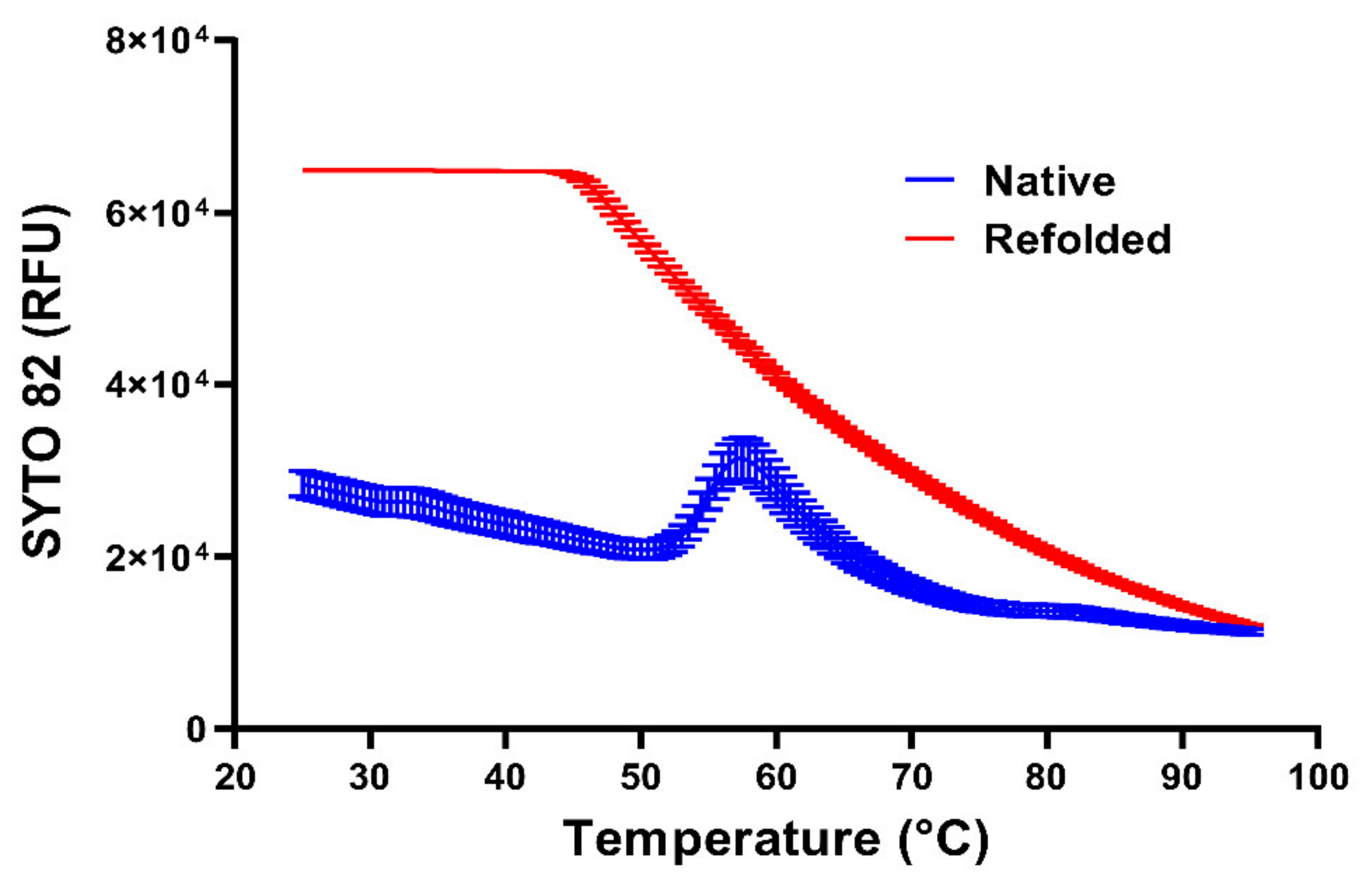
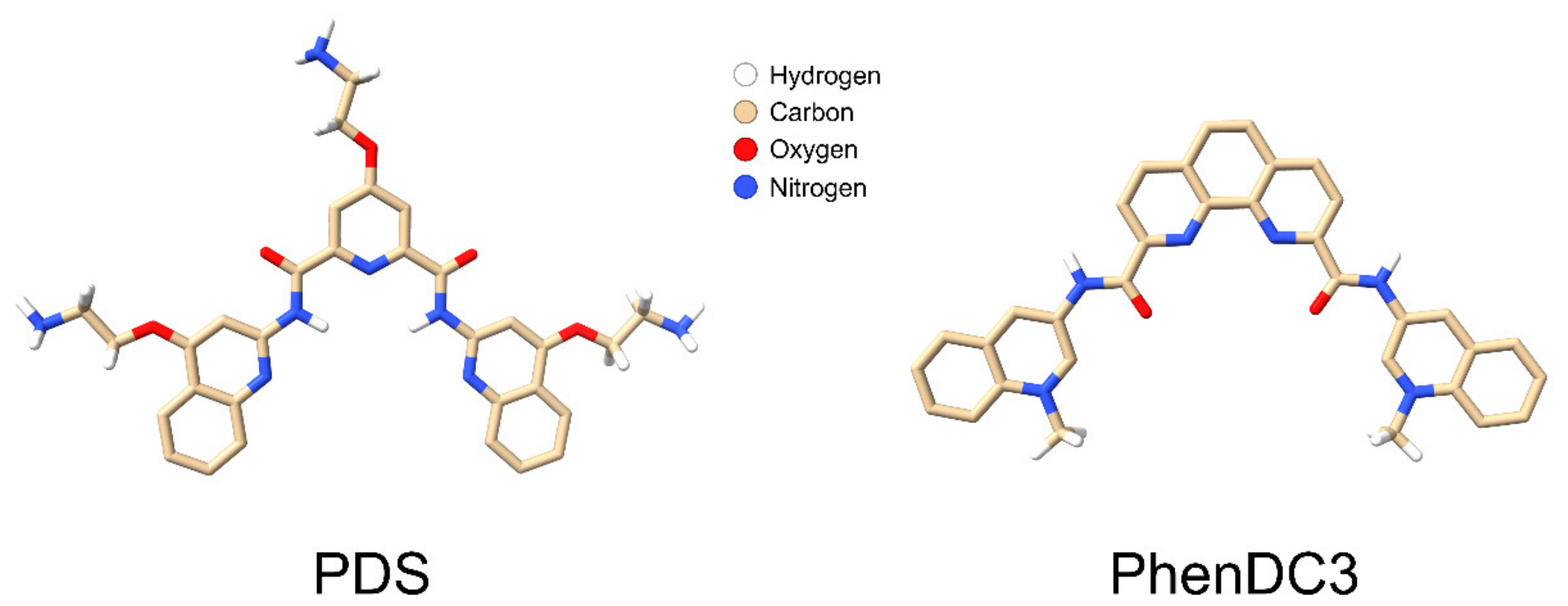
9. Stabilising GQs in Viral RNA
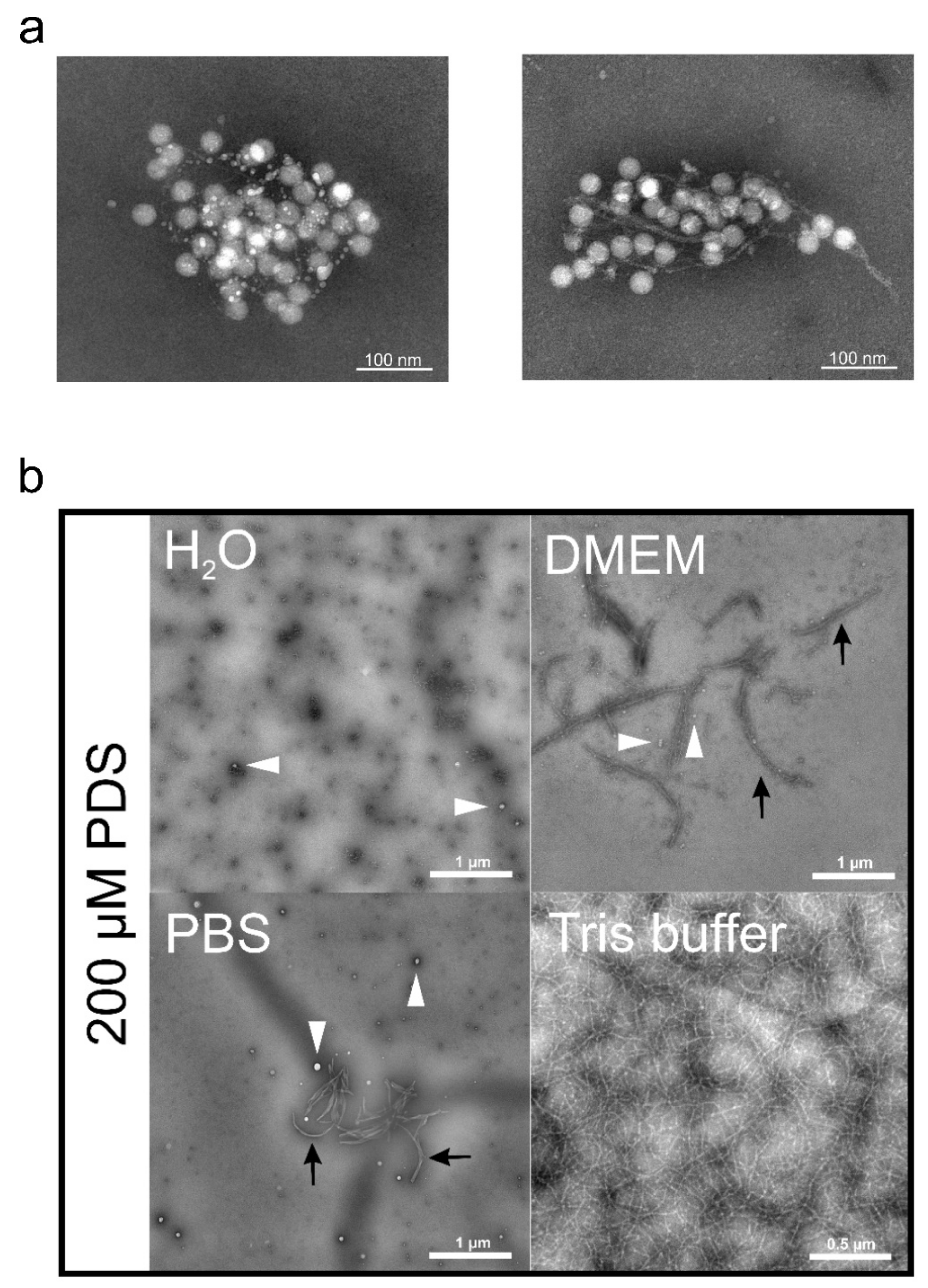
10. Inhibiting RNA Exit from Virions with PDS
11. Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palmenberg, A.C.; Gern, J.E. Classification and evolution of human rhinoviruses. Methods Mol. Biol. 2015, 1221, 1–10. [Google Scholar]
- Stott, E.J.; Killington, R.A. Rhinoviruses. Annu. Rev. Microbiol. 1972, 26, 503–524. [Google Scholar] [CrossRef] [PubMed]
- Gern, J.E.; Mosser, A.G.; Swenson, C.A.; Rennie, P.J.; England, R.J.; Shaffer, J.; Mizoguchi, H. Inhibition of rhinovirus replication in vitro and in vivo by acid-buffered saline. J. Infect. Dis. 2007, 195, 1137–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, S.E.; Lamson, D.M.; St George, K.; Walsh, T.J. Human rhinoviruses. Clin. Microbiol. Rev. 2013, 26, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fendrick, A.M.; Monto, A.S.; Nightengale, B.; Sarnes, M. The economic burden of non-influenza-related viral respiratory tract infection in the United States. Arch. Intern. Med. 2003, 163, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.L.; Turner, R.B.; Braciale, T.; Heymann, P.W.; Borish, L. Pathogenesis of rhinovirus infection. Curr. Opin. Virol. 2012, 2, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.P. Is a rhinovirus vaccine possible? Am. J. Epidemiol. 1976, 103, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.L. A, B & C Rhinoviruses—New Knowledge from an Impressive Consortium—A Step Forward for Rhinovirus Vaccine Efforts, or a Step Back? Am. J. Respir. Crit. Care Med. 2021, 203, 786–788. [Google Scholar]
- Choi, T.; Devries, M.; Bacharier, L.B.; Busse, W.; Camargo, C.A., Jr.; Cohen, R.; Demuri, G.P.; Evans, M.D.; Fitzpatrick, A.M.; Gergen, P.J.; et al. Enhanced Neutralizing Antibody Responses to Rhinovirus C and Age-Dependent Patterns of Infection. Am. J. Respir. Crit. Care Med. 2021, 203, 822–830. [Google Scholar] [CrossRef]
- Cooney, M.K.; Fox, J.P.; Kenny, G.E. Antigenic groupings of 90 rhinovirus serotypes. Infect. Immun. 1982, 37, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Edlmayr, J.; Niespodziana, K.; Popow-Kraupp, T.; Krzyzanek, V.; Focke-Tejkl, M.; Blaas, D.; Grote, M.; Valenta, R. Antibodies induced with recombinant VP1 from human rhinovirus exhibit cross-neutralisation. Eur. Respir. J. 2011, 37, 44–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katpally, U.; Fu, T.M.; Freed, D.C.; Casimiro, D.R.; Smith, T.J. Antibodies to the buried N terminus of rhinovirus VP4 exhibit cross-serotypic neutralization. J. Virol. 2009, 83, 7040–7048. [Google Scholar] [CrossRef] [Green Version]
- Hamory, B.H.; Hamparian, V.V.; Conant, R.M.; Gwaltney, J.M., Jr. Human responses to two decavalent rhinovirus vaccines. J. Infect. Dis. 1975, 132, 623–629. [Google Scholar] [CrossRef]
- McLean, G.R. Developing a vaccine for human rhinoviruses. J. Vaccines Immun. 2014, 2, 16–20. [Google Scholar] [PubMed] [Green Version]
- Bartlett, N.W.; Walton, R.P.; Edwards, M.R.; Aniscenko, J.; Caramori, G.; Zhu, J.; Glanville, N.; Choy, K.J.; Jourdan, P.; Burnet, J.; et al. Mouse models of rhinovirus-induced disease and exacerbation of allergic airway inflammation. Nat. Med. 2008, 14, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thibaut, H.J.; De Palma, A.M.; Neyts, J. Combating enterovirus replication: State-of-the-Art on antiviral research. Biochem. Pharmacol. 2012, 83, 185–192. [Google Scholar] [CrossRef]
- Van der Linden, L.; Wolthers, K.C.; van Kuppeveld, F.J. Replication and Inhibitors of Enteroviruses and Parechoviruses. Viruses 2015, 7, 4529–4562. [Google Scholar] [CrossRef] [Green Version]
- Staring, J.; von Castelmur, E.; Blomen, V.A.; van den Hengel, L.G.; Brockmann, M.; Baggen, J.; Thibaut, H.J.; Nieuwenhuis, J.; Janssen, H.; van Kuppeveld, F.J.; et al. PLA2G16 represents a switch between entry and clearance of Picornaviridae. Nature 2017, 541, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Elling, U.; Wimmer, R.A.; Leibbrandt, A.; Burkard, T.; Michlits, G.; Leopoldi, A.; Micheler, T.; Abdeen, D.; Zhuk, S.; Aspalter, I.M.; et al. A reversible haploid mouse embryonic stem cell biobank resource for functional genomics. Nature 2017, 550, 114–118. [Google Scholar] [CrossRef]
- Casanova, V.; Sousa, F.H.; Stevens, C.; Barlow, P.G. Antiviral therapeutic approaches for human rhinovirus infections. Future Virol. 2018, 13, 505–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, L.; Lyoo, H.; van der Schaar, H.M.; Strating, J.R.; van Kuppeveld, F.J. Direct-acting antivirals and host-targeting strategies to combat enterovirus infections. Curr. Opin. Virol. 2017, 24, 1–8. [Google Scholar] [CrossRef]
- Grassauer, A.; Weinmuellner, R.; Meier, C.; Pretsch, A.; Prieschl-Grassauer, E.; Unger, H. Iota-Carrageenan is a potent inhibitor of rhinovirus infection. Virol. J. 2008, 5, 107. [Google Scholar] [CrossRef] [Green Version]
- Cagno, V.; Andreozzi, P.; D’Alicarnasso, M.; Jacob Silva, P.; Mueller, M.; Galloux, M.; Le Goffic, R.; Jones, S.T.; Vallino, M.; Hodek, J.; et al. Broad-spectrum non-toxic antiviral nanoparticles with a virucidal inhibition mechanism. Nat. Mater. 2018, 17, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Santaquiteria, M.; Illescas, B.M.; Abdelnabi, R.; Boonen, A.; Mills, A.; Marti-Mari, O.; Noppen, S.; Neyts, J.; Schols, D.; Gago, F.; et al. Multivalent Tryptophan- and Tyrosine-Containing [60]Fullerene Hexa-Adducts as Dual HIV and Enterovirus A71 Entry Inhibitors. Chemistry 2021, 27, 10700–10710. [Google Scholar] [CrossRef]
- Real-Hohn, A.; Zhu, R.; Ganjian, H.; Ibrahim, N.; Hinterdorfer, P.; Kowalski, H.; Blaas, D. Catching Common Cold Virus with a Net: Pyridostatin Forms Filaments in Tris Buffer That Trap Viruses-A Novel Antiviral Strategy? Viruses 2020, 12, 723. [Google Scholar] [CrossRef] [PubMed]
- Diana, G.D.; McKinlay, M.A.; Brisson, C.J.; Zalay, E.S.; Miralles, J.V.; Salvador, U.J. Isoxazoles with antipicornavirus activity. J. Med. Chem. 1985, 28, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Pevear, D.C.; Fancher, M.J.; Felock, P.J.; Rossmann, M.G.; Miller, M.S.; Diana, G.; Treasurywala, A.M.; McKinlay, M.A.; Dutko, F.J. Conformational change in the floor of the human rhinovirus canyon blocks adsorption to HeLa cell receptors. J. Virol. 1989, 63, 2002–2007. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.J.; Kremer, M.J.; Luo, M.; Vriend, G.; Arnold, E.; Kamer, G.; Rossmann, M.G.; McKinlay, M.A.; Diana, G.D.; Otto, M.J. The site of attachment in human rhinovirus 14 for antiviral agents that inhibit uncoating. Science 1986, 233, 1286–1293. [Google Scholar] [CrossRef]
- Zhang, A.; Nanni, R.G.; Li, T.; Arnold, G.F.; Oren, D.A.; Jacobo-Molina, A.; Williams, R.L.; Kamer, G.; Rubenstein, D.A.; Li, Y.; et al. Structure determination of antiviral compound SCH 38057 complexed with human rhinovirus 14. J. Mol. Biol. 1993, 230, 857–867. [Google Scholar] [CrossRef]
- Woods, M.G.; Diana, G.D.; Rogge, M.C.; Otto, M.J.; Dutko, F.J.; McKinlay, M.A. In vitro and in vivo activities of WIN 54954, a new broad-spectrum antipicornavirus drug. Antimicrob. Agents Chemother. 1989, 33, 2069–2074. [Google Scholar] [CrossRef] [Green Version]
- Tsang, S.K.; Cheh, J.; Isaacs, L.; Joseph-McCarthy, D.; Choi, S.K.; Pevear, D.C.; Whitesides, G.M.; Hogle, J.M. A structurally biased combinatorial approach for discovering new anti-picornaviral compounds. Chem. Biol. 2001, 8, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Shepard, D.A.; Heinz, B.A.; Rueckert, R.R. WIN 52035-2 inhibits both attachment and eclipse of human rhinovirus 14. J. Virol. 1993, 67, 2245–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossmann, M.G. The structure of antiviral agents that inhibit uncoating when complexed with viral capsids. Antivir. Res. 1989, 11, 3–13. [Google Scholar] [CrossRef]
- Rosenwirth, B.; Oren, D.A.; Arnold, E.; Kis, Z.L.; Eggers, H.J. SDZ 35-682, a new picornavirus capsid-binding agent with potent antiviral activity. Antivir. Res. 1995, 26, 65–82. [Google Scholar] [CrossRef]
- Plevka, P.; Perera, R.; Yap, M.L.; Cardosa, J.; Kuhn, R.J.; Rossmann, M.G. Structure of human enterovirus 71 in complex with a capsid-binding inhibitor. Proc. Natl. Acad. Sci. USA 2013, 110, 5463–5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diana, G.D.; Otto, M.J.; McKinlay, M.A. Inhibitors of picornavirus uncoating as antiviral agents. Pharmacol. Ther. 1985, 29, 287–297. [Google Scholar] [CrossRef]
- Okun, V.M.; Nizet, S.; Blaas, D.; Kenndler, E. Kinetics of thermal denaturation of human rhinoviruses in the presence of anti-viral capsid binders analyzed by capillary electrophoresis. Electrophoresis 2002, 23, 896–902. [Google Scholar] [CrossRef]
- Phelps, D.K.; Post, C.B. Molecular dynamics investigation of the effect of an antiviral compound on human rhinovirus. Protein Sci. 1999, 8, 2281–2289. [Google Scholar] [CrossRef]
- Phelps, D.K.; Post, C.B. A novel basis of capsid stabilization by antiviral compounds. J. Mol. Biol. 1995, 254, 544–551. [Google Scholar] [CrossRef] [Green Version]
- Wald, J.; Pasin, M.; Richter, M.; Walther, C.; Mathai, N.; Kirchmair, J.; Makarov, V.A.; Goessweiner-Mohr, N.; Marlovits, T.C.; Zanella, I.; et al. Cryo-EM structure of pleconaril-resistant rhinovirus-B5 complexed to the antiviral OBR-5-340 reveals unexpected binding site. Proc. Natl. Acad. Sci. USA 2019, 116, 19109–19115. [Google Scholar] [CrossRef] [Green Version]
- Abdelnabi, R.; Geraets, J.A.; Ma, Y.; Mirabelli, C.; Flatt, J.W.; Domanska, A.; Delang, L.; Jochmans, D.; Kumar, T.A.; Jayaprakash, V.; et al. A novel druggable interprotomer pocket in the capsid of rhino- and enteroviruses. PLoS Biol. 2019, 17, e3000281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, Y.; He, W.; Sun, Y.; Guo, Y.; Zhong, W.; Gao, Q.; Liao, M.; Wang, X.; Cai, Y.; et al. Design, Synthesis, and Evaluation of Novel Enterovirus 71 Inhibitors as Therapeutic Drug Leads for the Treatment of Human Hand, Foot, and Mouth Disease. J. Med. Chem. 2020, 63, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sheng, J.; van Vliet, A.L.W.; Buda, G.; van Kuppeveld, F.J.M.; Rossmann, M.G. Molecular basis for the acid-initiated uncoating of human enterovirus D68. Proc. Natl. Acad. Sci. USA 2018, 115, E12209–E12217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garriga, D.; Pickl-Herk, A.; Luque, D.; Wruss, J.; Caston, J.R.; Blaas, D.; Verdaguer, N. Insights into minor group rhinovirus uncoating: The X-ray structure of the HRV2 empty capsid. PLoS Pathog. 2012, 8, e1002473. [Google Scholar] [CrossRef] [Green Version]
- Pickl-Herk, A.; Luque, D.; Vives-Adrian, L.; Querol-Audi, J.; Garriga, D.; Trus, B.L.; Verdaguer, N.; Blaas, D.; Caston, J.R. Uncoating of common cold virus is preceded by RNA switching as determined by X-ray and cryo-EM analyses of the subviral A-particle. Proc. Natl. Acad. Sci. USA 2013, 110, 20063–20068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, H.C.; Bostina, M.; Filman, D.J.; Hogle, J.M. Catching a virus in the act of RNA release: A novel poliovirus uncoating intermediate characterized by cryo-electron microscopy. J. Virol. 2010, 84, 4426–4441. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Sun, Y.; Fan, J.; Zhu, B.; Cao, L.; Gao, Q.; Zhang, Y.; Liu, H.; Rao, Z.; Wang, X. Structures of Coxsackievirus A10 unveil the molecular mechanisms of receptor binding and viral uncoating. Nat. Commun. 2018, 9, 4985. [Google Scholar] [CrossRef] [Green Version]
- Carson, S.D. Kinetic models for receptor-catalyzed conversion of coxsackievirus B3 to A-particles. J. Virol. 2014, 88, 11568–11575. [Google Scholar] [CrossRef] [Green Version]
- Lyu, K.; Ding, J.; Han, J.F.; Zhang, Y.; Wu, X.Y.; He, Y.L.; Qin, C.F.; Chen, R. Human enterovirus 71 uncoating captured at atomic resolution. J. Virol. 2014, 88, 3114–3126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Wang, X.; Hu, Z.; Gao, Q.; Sun, Y.; Li, X.; Porta, C.; Walter, T.S.; Gilbert, R.J.; Zhao, Y.; et al. Picornavirus uncoating intermediate captured in atomic detail. Nat. Commun. 2013, 4, 1929. [Google Scholar] [CrossRef] [Green Version]
- Casasnovas, J.M.; Springer, T.A. Pathway of rhinovirus disruption by soluble intercellular adhesion molecule 1 (ICAM-1): An intermediate in which ICAM-1 is bound and RNA is released. J. Virol. 1994, 68, 5882–5889. [Google Scholar] [CrossRef] [Green Version]
- Hoover-Litty, H.; Greve, J.M. Formation of rhinovirus-soluble ICAM-1 complexes and conformational changes in the virion. J. Virol. 1993, 67, 390–397. [Google Scholar] [CrossRef] [Green Version]
- Xing, L.; Casasnovas, J.M.; Cheng, R.H. Structural analysis of human rhinovirus complexed with ICAM-1 reveals the dynamics of receptor-mediated virus uncoating. J. Virol. 2003, 77, 6101–6107. [Google Scholar] [CrossRef] [Green Version]
- Konecsni, T.; Berka, U.; Pickl-Herk, A.; Bilek, G.; Khan, A.G.; Gajdzig, L.; Fuchs, R.; Blaas, D. Low pH-triggered beta-propeller switch of the low-density lipoprotein receptor assists rhinovirus infection. J. Virol. 2009, 83, 10922–10930. [Google Scholar] [CrossRef] [Green Version]
- Prchla, E.; Kuechler, E.; Blaas, D.; Fuchs, R. Uncoating of human rhinovirus serotype 2 from late endosomes. J. Virol. 1994, 68, 3713–3723. [Google Scholar] [CrossRef] [Green Version]
- Mukhamedova, L.; Fuzik, T.; Novacek, J.; Hrebik, D.; Pridal, A.; Marti, G.A.; Guerin, D.M.A.; Plevka, P. Virion structure and in vitro genome release mechanism of dicistrovirus Kashmir bee virus. J. Virol. 2021, 95, e01950-20. [Google Scholar] [CrossRef]
- Kalynych, S.; Fuzik, T.; Pridal, A.; de Miranda, J.; Plevka, P. Cryo-EM study of slow bee paralysis virus at low pH reveals iflavirus genome release mechanism. Proc. Natl. Acad. Sci. USA 2017, 114, 598–603. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Eugenia, R.; Durana, A.; Lopez-Marijuan, I.; Marti, G.A.; Guerin, D.M.A. X-ray structure of Triatoma virus empty capsid: Insights into the mechanism of uncoating and RNA release in dicistroviruses. J. Gen. Virol. 2016, 97, 2769–2779. [Google Scholar] [CrossRef] [PubMed]
- Buchta, D.; Fuzik, T.; Hrebik, D.; Levdansky, Y.; Sukenik, L.; Mukhamedova, L.; Moravcova, J.; Vacha, R.; Plevka, P. Enterovirus particles expel capsid pentamers to enable genome release. Nat. Commun. 2019, 10, 1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agirre, J.; Aloria, K.; Arizmendi, J.M.; Iloro, I.; Elortza, F.; Sanchez-Eugenia, R.; Marti, G.A.; Neumann, E.; Rey, F.A.; Guerin, D.M. Capsid protein identification and analysis of mature Triatoma virus (TrV) virions and naturally occurring empty particles. Virology 2011, 409, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Lonberg-Holm, K.; Noble-Harvey, J. Comparison of in vitro and cell-mediated alteration of a human Rhinovirus and its inhibition by sodium dodecyl sulfate. J. Virol. 1973, 12, 819–826. [Google Scholar] [CrossRef] [Green Version]
- Lonberg-Holm, K.; Yin, F.H. Antigenic determinants of infective and inactivated human rhinovirus type 2. J. Virol. 1973, 12, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snijder, J.; Uetrecht, C.; Rose, R.J.; Sanchez-Eugenia, R.; Marti, G.A.; Agirre, J.; Guerin, D.M.; Wuite, G.J.; Heck, A.J.; Roos, W.H. Probing the biophysical interplay between a viral genome and its capsid. Nat. Chem. 2013, 5, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Thibaut, H.J.; Lacroix, C.; De Palma, A.M.; Franco, D.; Decramer, M.; Neyts, J. Toward antiviral therapy/prophylaxis for rhinovirus-induced exacerbations of chronic obstructive pulmonary disease: Challenges, opportunities, and strategies. Rev. Med. Virol. 2016, 26, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Rotbart, H.A.; Webster, A.D.; Pleconaril Treatment Registry, G. Treatment of potentially life-threatening enterovirus infections with pleconaril. Clin. Infect. Dis. 2001, 32, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Radanovic, I.; Rkman, D.; Zekan, P.; Kutlesa, M.; Barsic, B. Chronic meningoencephalitis caused by Echo virus 6 in a patient with common variable immunodeficiency: Successful treatment with pleconaril. Wien. Klin. Wochenschr. 2018, 130, 70–72. [Google Scholar] [CrossRef]
- Liu, Y.; Sheng, J.; Fokine, A.; Meng, G.; Shin, W.H.; Long, F.; Kuhn, R.J.; Kihara, D.; Rossmann, M.G. Structure and inhibition of EV-D68, a virus that causes respiratory illness in children. Science 2015, 347, 71–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, F.G.; Herrington, D.T.; Coats, T.L.; Kim, K.; Cooper, E.C.; Villano, S.A.; Liu, S.; Hudson, S.; Pevear, D.C.; Collett, M.; et al. Pleconaril Respiratory Infection Study, G., Efficacy and safety of oral pleconaril for treatment of colds due to picornaviruses in adults: Results of 2 double-blind, randomized, placebo-controlled trials. Clin. Infect. Dis. 2003, 36, 1523–1532. [Google Scholar] [CrossRef] [Green Version]
- Feil, S.C.; Hamilton, S.; Krippner, G.Y.; Lin, B.; Luttick, A.; McConnell, D.B.; Nearn, R.; Parker, M.W.; Ryan, J.; Stanislawski, P.C.; et al. An Orally Available 3-Ethoxybenzisoxazole Capsid Binder with Clinical Activity against Human Rhinovirus. ACS Med. Chem. Lett. 2012, 3, 303–307. [Google Scholar] [CrossRef] [Green Version]
- Hiremath, C.N.; Filman, D.J.; Grant, R.A.; Hogle, J.M. Ligand-induced conformational changes in poliovirus-antiviral drug complexes. Acta Crystallogr. D Biol. Crystallogr. 1997, 53 Pt 5, 558–570. [Google Scholar] [CrossRef]
- Grant, R.A.; Hiremath, C.N.; Filman, D.J.; Syed, R.; Andries, K.; Hogle, J.M. Structures of poliovirus complexes with anti-viral drugs: Implications for viral stability and drug design. Curr. Biol. CB 1994, 4, 784–797. [Google Scholar] [CrossRef]
- Andries, K.; Dewindt, B.; Snoeks, J.; Willebrords, R.; van Eemeren, K.; Stokbroekx, R.; Janssen, P.A. In vitro activity of pirodavir (R 77975), a substituted phenoxy-pyridazinamine with broad-spectrum antipicornaviral activity. Antimicrob. Agents Chemother. 1992, 36, 100–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lentz, K.N.; Smith, A.D.; Geisler, S.C.; Cox, S.; Buontempo, P.; Skelton, A.; DeMartino, J.; Rozhon, E.; Schwartz, J.; Girijavallabhan, V.; et al. Structure of poliovirus type 2 Lansing complexed with antiviral agent SCH48973: Comparison of the structural and biological properties of three poliovirus serotypes. Structure 1997, 5, 961–978. [Google Scholar] [CrossRef] [Green Version]
- Hendry, E.; Hatanaka, H.; Fry, E.; Smyth, M.; Tate, J.; Stanway, G.; Santti, J.; Maaronen, M.; Hyypia, T.; Stuart, D. The crystal structure of coxsackievirus A9: New insights into the uncoating mechanisms of enteroviruses. Structure 1999, 7, 1527–1538. [Google Scholar] [CrossRef] [Green Version]
- Hiremath, C.N.; Grant, R.A.; Filman, D.J.; Hogle, J.M. Binding of the antiviral drug WIN51711 to the sabin strain of type 3 poliovirus: Structural comparison with drug binding in rhinovirus 14. Acta Crystallogr. D Biol. Crystallogr. 1995, 51 Pt 4, 473–489. [Google Scholar] [CrossRef]
- Hadfield, A.T.; Diana, G.D.; Rossmann, M.G. Analysis of three structurally related antiviral compounds in complex with human rhinovirus 16. Proc. Natl. Acad. Sci. USA 1999, 96, 14730–14735. [Google Scholar] [CrossRef] [Green Version]
- Hadfield, A.T.; Lee, W.; Zhao, R.; Oliveira, M.A.; Minor, I.; Rueckert, R.R.; Rossmann, M.G. The refined structure of human rhinovirus 16 at 2.15 A resolution: Implications for the viral life cycle. Structure 1997, 5, 427–441. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Simpson, A.A.; Ledford, R.M.; Bator, C.M.; Chakravarty, S.; Skochko, G.A.; Demenczuk, T.M.; Watanyar, A.; Pevear, D.C.; Rossmann, M.G. Structural and virological studies of the stages of virus replication that are affected by antirhinovirus compounds. J. Virol. 2004, 78, 11061–11069. [Google Scholar] [CrossRef] [Green Version]
- Jensen, L.M.; Walker, E.J.; Jans, D.A.; Ghildyal, R. Proteases of human rhinovirus: Role in infection. Methods Mol. Biol. 2015, 1221, 129–141. [Google Scholar]
- Arnold, E.; Luo, M.; Vriend, G.; Rossmann, M.G.; Palmenberg, A.C.; Parks, G.D.; Nicklin, M.J.; Wimmer, E. Implications of the picornavirus capsid structure for polyprotein processing. Proc. Natl. Acad. Sci. USA 1987, 84, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Richards, A.L.; Jackson, W.T. Intracellular vesicle acidification promotes maturation of infectious poliovirus particles. PLoS Pathog. 2012, 8, e1003046. [Google Scholar] [CrossRef] [Green Version]
- Haghighat, A.; Svitkin, Y.; Novoa, I.; Kuechler, E.; Skern, T.; Sonenberg, N. The eIF4G-eIF4E complex is the target for direct cleavage by the rhinovirus 2A proteinase. J. Virol. 1996, 70, 8444–8450. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.V.; Pelletier, J.; Meerovitch, K.; Sonenberg, N.; Kaufman, R.J. The effect of poliovirus proteinase 2Apro expression on cellular metabolism. Inhibition of DNA replication, RNA polymerase II transcription, and translation. J. Biol. Chem. 1991, 266, 14714–14720. [Google Scholar] [CrossRef]
- Lamphear, B.J.; Yan, R.; Yang, F.; Waters, D.; Liebig, H.D.; Klump, H.; Kuechler, E.; Skern, T.; Rhoads, R.E. Mapping the cleavage site in protein synthesis initiation factor eIF-4 gamma of the 2A proteases from human Coxsackievirus and rhinovirus. J. Biol. Chem. 1993, 268, 19200–19203. [Google Scholar] [CrossRef]
- Sommergruber, W.; Ahorn, H.; Zophel, A.; Maurer-Fogy, I.; Fessl, F.; Schnorrenberg, G.; Liebig, H.D.; Blaas, D.; Kuechler, E.; Skern, T. Cleavage specificity on synthetic peptide substrates of human rhinovirus 2 proteinase 2A. J. Biol. Chem. 1992, 267, 22639–22644. [Google Scholar] [CrossRef]
- Skern, T.; Sommergruber, W.; Auer, H.; Volkmann, P.; Zorn, M.; Liebig, H.D.; Fessl, F.; Blaas, D.; Kuechler, E. Substrate requirements of a human rhinoviral 2A proteinase. Virology 1991, 181, 46–54. [Google Scholar] [CrossRef]
- Sommergruber, W.; Zorn, M.; Blaas, D.; Fessl, F.; Volkmann, P.; Maurer-Fogy, I.; Pallai, P.; Merluzzi, V.; Matteo, M.; Skern, T.; et al. Polypeptide 2A of human rhinovirus type 2: Identification as a protease and characterization by mutational analysis. Virology 1989, 169, 68–77. [Google Scholar] [CrossRef]
- Seipelt, J.; Liebig, H.D.; Sommergruber, W.; Gerner, C.; Kuechler, E. 2A proteinase of human rhinovirus cleaves cytokeratin 8 in infected HeLa cells. J. Biol. Chem. 2000, 275, 20084–20089. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.; Zhang, J.; Yanagawa, B.; Luo, Z.; Yang, X.; Chang, J.; McManus, B.; Luo, H. Cleavage of serum response factor mediated by enteroviral protease 2A contributes to impaired cardiac function. Cell Res. 2012, 22, 360–371. [Google Scholar] [CrossRef]
- Krenn, B.M.; Gaudernak, E.; Holzer, B.; Lanke, K.; Van Kuppeveld, F.J.; Seipelt, J. Antiviral activity of the zinc ionophores pyrithione and hinokitiol against picornavirus infections. J. Virol. 2009, 83, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Gaudernak, E.; Seipelt, J.; Triendl, A.; Grassauer, A.; Kuechler, E. Antiviral effects of pyrrolidine dithiocarbamate on human rhinoviruses. J. Virol. 2002, 76, 6004–6015. [Google Scholar] [CrossRef] [Green Version]
- Molla, A.; Hellen, C.U.; Wimmer, E. Inhibition of proteolytic activity of poliovirus and rhinovirus 2A proteinases by elastase-specific inhibitors. J. Virol. 1993, 67, 4688–4695. [Google Scholar] [CrossRef] [Green Version]
- Deszcz, L.; Seipelt, J.; Vassilieva, E.; Roetzer, A.; Kuechler, E. Antiviral activity of caspase inhibitors: Effect on picornaviral 2A proteinase. FEBS Lett. 2004, 560, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Bauer, L.; Manganaro, R.; Zonsics, B.; Hurdiss, D.L.; Zwaagstra, M.; Donselaar, T.; Welter, N.G.E.; van Kleef, R.; Lopez, M.L.; Bevilacqua, F.; et al. Rational design of highly potent broad-spectrum enterovirus inhibitors targeting the nonstructural protein 2C. PLoS Biol. 2020, 18, e3000904. [Google Scholar] [CrossRef] [PubMed]
- Bauer, L.; Manganaro, R.; Zonsics, B.; Strating, J.; El Kazzi, P.; Lorenzo Lopez, M.; Ulferts, R.; van Hoey, C.; Mate, M.J.; Langer, T.; et al. Fluoxetine Inhibits Enterovirus Replication by Targeting the Viral 2C Protein in a Stereospecific Manner. ACS Infect. Dis. 2019, 5, 1609–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, B.A.; Vance, L.M. The antiviral compound enviroxime targets the 3A coding region of rhinovirus and poliovirus. J. Virol. 1995, 69, 4189–4197. [Google Scholar] [CrossRef] [Green Version]
- Dragovich, P.S.; Prins, T.J.; Zhou, R.; Webber, S.E.; Marakovits, J.T.; Fuhrman, S.A.; Patick, A.K.; Matthews, D.A.; Lee, C.A.; Ford, C.E.; et al. Structure-based design, synthesis, and biological evaluation of irreversible human rhinovirus 3C protease inhibitors. 4. Incorporation of P1 lactam moieties as L-glutamine replacements. J. Med. Chem. 1999, 42, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Hayden, F.G.; Turner, R.B.; Gwaltney, J.M.; Chi-Burris, K.; Gersten, M.; Hsyu, P.; Patick, A.K.; Smith, G.J., 3rd; Zalman, L.S. Phase II, randomized, double-blind, placebo-controlled studies of ruprintrivir nasal spray 2-percent suspension for prevention and treatment of experimentally induced rhinovirus colds in healthy volunteers. Antimicrob. Agents Chemother. 2003, 47, 3907–3916. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, C.; George, S.; Leyssen, P.; Hilgenfeld, R.; Neyts, J. The enterovirus 3C protease inhibitor SG85 efficiently blocks rhinovirus replication and is not cross-resistant with rupintrivir. Antimicrob. Agents Chemother. 2015, 59, 5814–5818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, D.O.; Murray, B.K.; Robins, R.K.; North, J.A. In vitro Antiviral Activity of Ribavirin against Picornaviruses. Antivir. Chem. Chemother. 1992, 3, 361–370. [Google Scholar] [CrossRef]
- Smee, D.F.; Evans, W.J.; Nicolaou, K.C.; Tarbet, E.B.; Day, C.W. Susceptibilities of enterovirus D68, enterovirus 71, and rhinovirus 87 strains to various antiviral compounds. Antivir. Res. 2016, 131, 61–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, Y.; Takahashi, K.; Fukuda, Y.; Kuno, M.; Kamiyama, T.; Kozaki, K.; Nomura, N.; Egawa, H.; Minami, S.; Watanabe, Y.; et al. In vitro and in vivo activities of anti-influenza virus compound T-705. Antimicrob. Agents Chemother. 2002, 46, 977–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nirwan, S.; Kakkar, R. Rhinovirus RNA Polymerase: Structure, Function, and Inhibitors. Viral Polym. 2019, 10, 301–331. [Google Scholar] [CrossRef]
- Scholle, M.D.; McLaughlin, D.; Gurard-Levin, Z.A. High-Throughput Affinity Selection Mass Spectrometry Using SAMDI-MS to Identify Small-Molecule Binders of the Human Rhinovirus 3C Protease. SLAS Discov. 2021, 26, 24725552211023211. [Google Scholar] [CrossRef]
- Yuan, S.; Fan, K.; Chen, Z.; Sun, Y.; Hou, H.; Zhu, L. Structure of the HRV-C 3C-Rupintrivir Complex Provides New Insights for Inhibitor Design. Virol Sin. 2020, 35, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Wanga, Q.M.; Chen, S.H. Human rhinovirus 3C protease as a potential target for the development of antiviral agents. Curr. Protein Pept. Sci. 2007, 8, 19–27. [Google Scholar] [CrossRef]
- Wang, Q.M. Protease inhibitors as potential antiviral agents for the treatment of picornaviral infections. In Progress in Drug Research; Jucker, E., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; Volume 52, p. 23. [Google Scholar]
- Matthews, D.A.; Smith, W.W.; Ferre, R.A.; Condon, B.; Budahazi, G.; Sisson, W.; Villafranca, J.E.; Janson, C.A.; McElroy, H.E.; Gribskov, C.L.; et al. Structure of human rhinovirus 3C protease reveals a trypsin-like polypeptide fold, RNA-binding site, and means for cleaving precursor polyprotein. Cell 1994, 77, 761–771. [Google Scholar] [CrossRef]
- Sommergruber, W.; Seipelt, J.; Fessl, F.; Skern, T.; Liebig, H.D.; Casari, G. Mutational analyses support a model for the HRV2 2A proteinase. Virology 1997, 234, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Van der Schaar, H.M.; Dorobantu, C.M.; Albulescu, L.; Strating, J.; van Kuppeveld, F.J.M. Fat(al) attraction: Picornaviruses Usurp Lipid Transfer at Membrane Contact Sites to Create Replication Organelles. Trends Microbiol. 2016, 24, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Belov, G.A. Dynamic lipid landscape of picornavirus replication organelles. Curr. Opin. Virol. 2016, 19, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Van der Linden, L.; Vives-Adrian, L.; Selisko, B.; Ferrer-Orta, C.; Liu, X.; Lanke, K.; Ulferts, R.; De Palma, A.M.; Tanchis, F.; Goris, N.; et al. The RNA template channel of the RNA-dependent RNA polymerase as a target for development of antiviral therapy of multiple genera within a virus family. PLoS Pathog. 2015, 11, e1004733. [Google Scholar] [CrossRef] [Green Version]
- Rightsel, W.A.; Dice, J.R.; Mc, A.R.; Timm, E.A.; Mc, L.I., Jr.; Dixon, G.J.; Schabel, F.M., Jr. Antiviral effect of guanidine. Science 1961, 134, 558–559. [Google Scholar] [CrossRef]
- Manganaro, R.; Zonsics, B.; Bauer, L.; Lorenzo Lopez, M.; Donselaar, T.; Zwaagstra, M.; Saporito, F.; Ferla, S.; Strating, J.; Coutard, B.; et al. Synthesis and antiviral effect of novel fluoxetine analogues as enterovirus 2C inhibitors. Antivir. Res. 2020, 178, 104781. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Xu, Z.; Jin, M.; Shu, T.; Chen, Y.; Feng, L.; Zhang, Q.; Lan, K.; Wu, S.; Zhou, H.B. Identification of dibucaine derivatives as novel potent enterovirus 2C helicase inhibitors: In vitro, in vivo, and combination therapy study. Eur. J. Med. Chem. 2020, 202, 112310. [Google Scholar] [CrossRef]
- Baggen, J.; Liu, Y.; Lyoo, H.; van Vliet, A.L.W.; Wahedi, M.; de Bruin, J.W.; Roberts, R.W.; Overduin, P.; Meijer, A.; Rossmann, M.G.; et al. Bypassing pan-enterovirus host factor PLA2G16. Nat. Commun. 2019, 10, 3171. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Mock, E.D.; Martella, A.; Kantae, V.; Di, X.; Burggraaff, L.; Baggelaar, M.P.; Al-Ayed, K.; Bakker, A.; Florea, B.I.; et al. Activity-Based Protein Profiling Identifies alpha-Ketoamides as Inhibitors for Phospholipase A2 Group XVI. ACS Chem. Biol. 2019, 14, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworski, K.; Ahmadian, M.; Duncan, R.E.; Sarkadi-Nagy, E.; Varady, K.A.; Hellerstein, M.K.; Lee, H.Y.; Samuel, V.T.; Shulman, G.I.; Kim, K.H.; et al. AdPLA ablation increases lipolysis and prevents obesity induced by high-fat feeding or leptin deficiency. Nat. Med. 2009, 15, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Tsou, Y.L.; Lin, Y.W.; Chang, H.W.; Lin, H.Y.; Shao, H.Y.; Yu, S.L.; Liu, C.C.; Chitra, E.; Sia, C.; Chow, Y.H. Heat shock protein 90: Role in enterovirus 71 entry and assembly and potential target for therapy. PLoS ONE 2013, 8, e77133. [Google Scholar] [CrossRef]
- Wang, R.Y.; Kuo, R.L.; Ma, W.C.; Huang, H.I.; Yu, J.S.; Yen, S.M.; Huang, C.R.; Shih, S.R. Heat shock protein-90-beta facilitates enterovirus 71 viral particles assembly. Virology 2013, 443, 236–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.P.; Shan, G.Z.; Peng, Z.G.; Zhu, J.H.; Meng, S.; Zhang, T.; Gao, L.Y.; Tao, P.Z.; Gao, R.M.; Li, Y.H.; et al. Synthesis and biological evaluation of heat-shock protein 90 inhibitors: Geldanamycin derivatives with broad antiviral activities. Antivir. Chem. Chemother. 2010, 20, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Shim, A.; Kim, Y.J.; Ahn, J.H.; Kwon, B.E.; Pham, T.T.; Lee, J.; Chang, S.Y.; Ko, H.J. Antiviral and Anti-Inflammatory Activities of Pochonin D, a Heat Shock Protein 90 Inhibitor, against Rhinovirus Infection. Biomol. Ther. 2018, 26, 576–583. [Google Scholar] [CrossRef]
- Bauer, L.; Ferla, S.; Head, S.A.; Bhat, S.; Pasunooti, K.K.; Shi, W.Q.; Albulescu, L.; Liu, J.O.; Brancale, A.; van Kuppeveld, F.J.M.; et al. Structure-activity relationship study of itraconazole, a broad-range inhibitor of picornavirus replication that targets oxysterol-binding protein (OSBP). Antivir. Res. 2018, 156, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Corbic Ramljak, I.; Stanger, J.; Real-Hohn, A.; Dreier, D.; Wimmer, L.; Redlberger-Fritz, M.; Fischl, W.; Klingel, K.; Mihovilovic, M.D.; Blaas, D.; et al. Cellular N-myristoyltransferases play a crucial picornavirus genus-specific role in viral assembly, virion maturation, and infectivity. PLoS Pathog. 2018, 14, e1007203. [Google Scholar] [CrossRef] [PubMed]
- Mousnier, A.; Bell, A.S.; Swieboda, D.P.; Morales-Sanfrutos, J.; Perez-Dorado, I.; Brannigan, J.A.; Newman, J.; Ritzefeld, M.; Hutton, J.A.; Guedan, A.; et al. Fragment-derived inhibitors of human N-myristoyltransferase block capsid assembly and replication of the common cold virus. Nat. Chem. 2018, 10, 599–606. [Google Scholar] [CrossRef]
- Majee, P.; Pattnaik, A.; Sahoo, B.R.; Shankar, U.; Pattnaik, A.K.; Kumar, A.; Nayak, D. Inhibition of Zika virus replication by G-quadruplex-binding ligands. Mol. Ther. Nucleic Acids 2021, 23, 691–701. [Google Scholar] [CrossRef]
- Artusi, S.; Ruggiero, E.; Nadai, M.; Tosoni, B.; Perrone, R.; Ferino, A.; Zanin, I.; Xodo, L.; Flamand, L.; Richter, S.N. Antiviral Activity of the G-Quadruplex Ligand TMPyP4 against Herpes Simplex Virus-1. Viruses 2021, 13, 196. [Google Scholar] [CrossRef]
- Shen, L.W.; Qian, M.Q.; Yu, K.; Narva, S.; Yu, F.; Wu, Y.L.; Zhang, W. Inhibition of Influenza A virus propagation by benzoselenoxanthenes stabilizing TMPRSS2 Gene G-quadruplex and hence down-regulating TMPRSS2 expression. Sci. Rep. 2020, 10, 7635. [Google Scholar] [CrossRef] [PubMed]
- Rohde, G. Drug targets in rhinoviral infections. Infect. Disord. Drug Targets 2009, 9, 126–132. [Google Scholar] [CrossRef]
- Norder, H.; De Palma, A.M.; Selisko, B.; Costenaro, L.; Papageorgiou, N.; Arnan, C.; Coutard, B.; Lantez, V.; De Lamballerie, X.; Baronti, C.; et al. Picornavirus non-structural proteins as targets for new anti-virals with broad activity. Antivir. Res. 2011, 89, 204–218. [Google Scholar] [CrossRef]
- Denani, C.B.; Real-Hohn, A.; de Carvalho, C.A.M.; Gomes, A.M.O.; Goncalves, R.B. Lactoferrin affects rhinovirus B-14 entry into H1-HeLa cells. Arch. Virol. 2021, 166, 1203–1211. [Google Scholar] [CrossRef]
- Weng, T.Y.; Chen, L.C.; Shyu, H.W.; Chen, S.H.; Wang, J.R.; Yu, C.K.; Lei, H.Y.; Yeh, T.M. Lactoferrin inhibits enterovirus 71 infection by binding to VP1 protein and host cells. Antivir. Res. 2005, 67, 31–37. [Google Scholar] [CrossRef]
- Seganti, L.; Di Biase, A.M.; Marchetti, M.; Pietrantoni, A.; Tinari, A.; Superti, F. Antiviral activity of lactoferrin towards naked viruses. Biometals 2004, 17, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Chu, C.; Chiu, C.H. Lactoferrin inhibits enterovirus 71 infection of human embryonal rhabdomyosarcoma cells in vitro. J. Infect. Dis. 2002, 186, 1161–1164. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, M.; Superti, F.; Ammendolia, M.G.; Rossi, P.; Valenti, P.; Seganti, L. Inhibition of poliovirus type 1 infection by iron-, manganese- and zinc-saturated lactoferrin. Med. Microbiol. Immunol. 1999, 187, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Eccles, R.; Winther, B.; Johnston, S.L.; Robinson, P.; Trampisch, M.; Koelsch, S. Efficacy and safety of iota-carrageenan nasal spray versus placebo in early treatment of the common cold in adults: The ICICC trial. Respir. Res. 2015, 16, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bichiri, D.; Rente, A.R.; Jesus, A. Safety and efficacy of iota-carrageenan nasal spray in treatment and prevention of the common cold. Med. Pharm. Rep. 2021, 94, 28–34. [Google Scholar] [PubMed]
- Geller, R.; Vignuzzi, M.; Andino, R.; Frydman, J. Evolutionary constraints on chaperone-mediated folding provide an antiviral approach refractory to development of drug resistance. Genes Dev. 2007, 21, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Brabec, M.; Blaas, D.; Fuchs, R. Wortmannin delays transfer of human rhinovirus serotype 2 to late endocytic compartments. Biochem. Biophys. Res. Commun. 2006, 348, 741–749. [Google Scholar] [CrossRef]
- Ganjian, H.; Zietz, C.; Mechtcheriakova, D.; Blaas, D.; Fuchs, R. ICAM-1 Binding Rhinoviruses Enter HeLa Cells via Multiple Pathways and Travel to Distinct Intracellular Compartments for Uncoating. Viruses 2017, 9, 68. [Google Scholar] [CrossRef] [Green Version]
- Albulescu, L.; Bigay, J.; Biswas, B.; Weber-Boyvat, M.; Dorobantu, C.M.; Delang, L.; van der Schaar, H.M.; Jung, Y.S.; Neyts, J.; Olkkonen, V.M.; et al. Uncovering oxysterol-binding protein (OSBP) as a target of the anti-enteroviral compound TTP-8307. Antivir. Res. 2017, 140, 37–44. [Google Scholar] [CrossRef]
- Blaas, D. Viral entry pathways: The example of common cold viruses. Wien. Med. Wochenschr. 2016, 166, 211–226. [Google Scholar] [CrossRef] [Green Version]
- Jurgeit, A.; McDowell, R.; Moese, S.; Meldrum, E.; Schwendener, R.; Greber, U.F. Niclosamide is a proton carrier and targets acidic endosomes with broad antiviral effects. PLoS Pathog. 2012, 8, e1002976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Yamaya, M.; Sekizawa, K.; Hosoda, M.; Yamada, N.; Ishizuka, S.; Nakayama, K.; Yanai, M.; Numazaki, Y.; Sasaki, H. Bafilomycin A(1) inhibits rhinovirus infection in human airway epithelium: Effects on endosome and ICAM-1. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 280, L1115–L1127. [Google Scholar] [CrossRef] [Green Version]
- Pfanzagl, B.; Andergassen, D.; Edlmayr, J.; Niespodziana, K.; Valenta, R.; Blaas, D. Entry of human rhinovirus 89 via ICAM-1 into HeLa epithelial cells is inhibited by actin skeleton disruption and by bafilomycin. Arch. Virol. 2014, 159, 125–140. [Google Scholar] [CrossRef]
- Nurani, G.; Lindqvist, B.; Casasnovas, J.M. Receptor priming of major group human rhinoviruses for uncoating and entry at mild low-pH environments. J. Virol. 2003, 77, 11985–11991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.R.; Song, J.H.; Ahn, J.H.; Lee, G.S.; Ahn, H.; Yoon, S.I.; Kang, S.G.; Kim, P.H.; Jeon, S.M.; Choi, E.J.; et al. Antiviral and anti-inflammatory activity of budesonide against human rhinovirus infection mediated via autophagy activation. Antivir. Res. 2018, 151, 87–96. [Google Scholar] [CrossRef]
- Real-Hohn, A.; Provance, D.W., Jr.; Goncalves, R.B.; Denani, C.B.; de Oliveira, A.C.; Salerno, V.P.; Oliveira Gomes, A.M. Impairing the function of MLCK, myosin Va or myosin Vb disrupts Rhinovirus B14 replication. Sci. Rep. 2017, 7, 17153. [Google Scholar] [CrossRef] [PubMed]
- Konecsni, T.; Kremser, L.; Snyers, L.; Rankl, C.; Kilar, F.; Kenndler, E.; Blaas, D. Twelve receptor molecules attach per viral particle of human rhinovirus serotype 2 via multiple modules. FEBS Lett. 2004, 568, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Crump, C.E.; Arruda, E.; Hayden, F.G. Comparative antirhinoviral activities of soluble intercellular adhesion molecule-1 (sICAM-1) and chimeric ICAM-1/immunoglobulin A molecule. Antimicrob. Agents Chemother. 1994, 38, 1425–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlin, A.; Hoover-Litty, H.; Sanderson, G.; Paessens, A.; Johnston, S.L.; Holgate, S.T.; Huguenel, E.; Greve, J.M. Spectrum of activity of soluble intercellular adhesion molecule-1 against rhinovirus reference strains and field isolates. Antimicrob. Agents Chemother. 1994, 38, 1413–1415. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.; Casanovas, J.M.; Staunton, D.E.; Springer, T.A. [Successful prevention of rhinovirus infections with chimeric ICAM-1 immunoglobulin in vitro]. Med. Klin. 1993, 88, 193–197. [Google Scholar]
- Rivero-Buceta, E.; Sun, L.; Martinez-Gualda, B.; Doyaguez, E.G.; Donckers, K.; Quesada, E.; Camarasa, M.J.; Delang, L.; San-Felix, A.; Neyts, J.; et al. Optimization of a Class of Tryptophan Dendrimers That Inhibit HIV Replication Leads to a Selective, Specific, and Low-Nanomolar Inhibitor of Clinical Isolates of Enterovirus A71. Antimicrob. Agents Chemother. 2016, 60, 5064–5067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualdoni, G.A.; Mayer, K.A.; Kapsch, A.M.; Kreuzberg, K.; Puck, A.; Kienzl, P.; Oberndorfer, F.; Fruhwirth, K.; Winkler, S.; Blaas, D.; et al. Rhinovirus induces an anabolic reprogramming in host cell metabolism essential for viral replication. Proc. Natl. Acad. Sci. USA 2018, 115, E7158–E7165. [Google Scholar] [CrossRef] [Green Version]
- Gielen, V.; Johnston, S.L.; Edwards, M.R. Azithromycin induces anti-viral responses in bronchial epithelial cells. Eur. Respir. J. 2010, 36, 646. [Google Scholar] [CrossRef] [Green Version]
- Sousa, F.H.; Casanova, V.; Findlay, F.; Stevens, C.; Svoboda, P.; Pohl, J.; Proudfoot, L.; Barlow, P.G. Cathelicidins display conserved direct antiviral activity towards rhinovirus. Peptides 2017, 95, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Real-Hohn, A.; Groznica, M.; Kontaxis, G.; Zhu, R.; Chaves, O.; Vazquez, L.; Hinterdorfer, P.; Blaas, D.; Kowalski, H. Targeting G-quadruplexes in the rhinovirus genome by Pyridostatin inhibits uncoating and highlights a critical role for sodium ions. Res. Square 2021. [Google Scholar] [CrossRef]
- Ahlquist, P.; Kaesberg, P. Determination of the length distribution of poly(A) at the 3′ terminus of the virion RNAs of EMC virus, poliovirus, rhinovirus, RAV-61 and CPMV and of mouse globin mRNA. Nucleic Acids Res. 1979, 7, 1195–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, C.; Mueller, S.; Paul, A.V.; Wimmer, E.; Jiang, P. Direct interaction between two viral proteins, the nonstructural protein 2C and the capsid protein VP3, is required for enterovirus morphogenesis. PLoS Pathog. 2010, 6, e1001066. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Liu, Y.; Wimmer, E.; Paul, A.V. Complete protein linkage map between the P2 and P3 non-structural proteins of poliovirus. J. Gen. Virol. 2007, 88 Pt 8, 2259–2267. [Google Scholar] [CrossRef]
- Li, J.P.; Baltimore, D. An intragenic revertant of a poliovirus 2C mutant has an uncoating defect. J. Virol. 1990, 64, 1102–1107. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.R.; Racaniello, V.R. Changes in rhinovirus protein 2C allow efficient replication in mouse cells. J. Virol. 2003, 77, 4773–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.H.; Wang, K.; Zhao, K.; Hua, S.C.; Du, J. The Structure, Function, and Mechanisms of Action of Enterovirus Non-structural Protein 2C. Front. Microbiol. 2020, 11, 615965. [Google Scholar] [CrossRef]
- Asare, E.; Mugavero, J.; Jiang, P.; Wimmer, E.; Paul, A.V. A Single Amino Acid Substitution in Poliovirus Nonstructural Protein 2CATPase Causes Conditional Defects in Encapsidation and Uncoating. J. Virol. 2016, 90, 6174–6186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfister, T.; Pasamontes, L.; Troxler, M.; Egger, D.; Bienz, K. Immunocytochemical localization of capsid-related particles in subcellular fractions of poliovirus-infected cells. Virology 1992, 188, 676–684. [Google Scholar] [CrossRef]
- Li, C.; Wang, J.C.; Taylor, M.W.; Zlotnick, A. In vitro assembly of an empty picornavirus capsid follows a dodecahedral path. J. Virol. 2012, 86, 13062–13069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, A.J.; Hu, Y.H.; Keinan, E. Chemical mimicry of viral capsid self-assembly. Proc. Natl. Acad. Sci. USA 2007, 104, 20731–20736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molla, A.; Paul, A.V.; Wimmer, E. Cell-free, de novo synthesis of poliovirus. Science 1991, 254, 1647–1651. [Google Scholar] [CrossRef]
- Marsian, J.; Fox, H.; Bahar, M.W.; Kotecha, A.; Fry, E.E.; Stuart, D.I.; Macadam, A.J.; Rowlands, D.J.; Lomonossoff, G.P. Plant-made polio type 3 stabilized VLPs-a candidate synthetic polio vaccine. Nat. Commun. 2017, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Shakeel, S.; Dykeman, E.C.; White, S.J.; Ora, A.; Cockburn, J.J.B.; Butcher, S.J.; Stockley, P.G.; Twarock, R. Genomic RNA folding mediates assembly of human parechovirus. Nat. Commun. 2017, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Chandler-Bostock, R.; Mata, C.P.; Bingham, R.J.; Dykeman, E.C.; Meng, B.; Tuthill, T.J.; Rowlands, D.J.; Ranson, N.A.; Twarock, R.; Stockley, P.G. Assembly of infectious enteroviruses depends on multiple, conserved genomic RNA-coat protein contacts. PLoS Pathog. 2020, 16, e1009146. [Google Scholar] [CrossRef]
- Twarock, R.; Stockley, P.G. RNA-Mediated Virus Assembly: Mechanisms and Consequences for Viral Evolution and Therapy. Annu. Rev. Biophys. 2019, 48, 495–514. [Google Scholar] [CrossRef] [PubMed]
- Twarock, R.; Bingham, R.J.; Dykeman, E.C.; Stockley, P.G. A modelling paradigm for RNA virus assembly. Curr. Opin. Virol. 2018, 31, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Wroblewski, E.; Leonov, G.; Phillips, S.E.V.; Tuma, R.; Twarock, R.; Stockley, P.G. Rewriting nature’s assembly manual for a ssRNA virus. Proc. Natl. Acad. Sci. USA 2017, 114, 12255–12260. [Google Scholar] [CrossRef] [Green Version]
- Blaas, D. Individual subunits of a rhinovirus causing common cold exhibit largely different protein-RNA contact site conformations. Commun. Biol. 2020, 3, 537. [Google Scholar] [CrossRef]
- Harutyunyan, S.; Kumar, M.; Sedivy, A.; Subirats, X.; Kowalski, H.; Kohler, G.; Blaas, D. Viral uncoating is directional: Exit of the genomic RNA in a common cold virus starts with the poly-(A) tail at the 3′-end. PLoS Pathog. 2013, 9, e1003270. [Google Scholar] [CrossRef] [PubMed]
- Harutyunyan, S.; Kowalski, H.; Blaas, D. The Rhinovirus subviral a-particle exposes 3′-terminal sequences of its genomic RNA. J. Virol. 2014, 88, 6307–6317. [Google Scholar] [CrossRef] [Green Version]
- Palmenberg, A.C.; Sgro, J.Y. Topological organization of picornaviral genomes: Statistical prediction of RNA structural signals. Semin. Virol. 1997, 8, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Strauss, M.; Levy, H.C.; Bostina, M.; Filman, D.J.; Hogle, J.M. RNA transfer from poliovirus 135S particles across membranes is mediated by long umbilical connectors. J. Virol. 2013, 87, 3903–3914. [Google Scholar] [CrossRef] [Green Version]
- Hrebík, D.; Füzik, T.; Gondová, M.; Šmerdová, L.; Adamopoulos, A.; Šedo, O.; Zdráhal, Z.; Plevka, P. ICAM-1 induced rearrangements of capsid and genome prime rhinovirus 14 for activation and uncoating. Proc. Natl. Acad. Sci. USA 2021, 118, e2024251118. [Google Scholar] [CrossRef]
- Real-Hohn, A.; Groznica, M.; Loffler, N.; Blaas, D.; Kowalski, H. nanoDSF: In vitro Label-Free Method to Monitor Picornavirus Uncoating and Test Compounds Affecting Particle Stability. Front. Microbiol. 2020, 11, 1442. [Google Scholar] [CrossRef]
- Jing, P.; Burris, B.; Zhang, R. Forces from the Portal Govern the Late-Stage DNA Transport in a Viral DNA Packaging Nanomotor. Biophys. J. 2016, 111, 162–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopal, A.; Egecioglu, D.E.; Yoffe, A.M.; Ben-Shaul, A.; Rao, A.L.; Knobler, C.M.; Gelbart, W.M. Viral RNAs are unusually compact. PLoS ONE 2014, 9, e105875. [Google Scholar] [CrossRef]
- Voss, N.R.; Gerstein, M. Calculation of standard atomic volumes for RNA and comparison with proteins: RNA is packed more tightly. J. Mol. Biol. 2005, 346, 477–492. [Google Scholar] [CrossRef]
- Kuznetsov, Y.G.; Daijogo, S.; Zhou, J.; Semler, B.L.; McPherson, A. Atomic force microscopy analysis of icosahedral virus RNA. J. Mol. Biol. 2005, 347, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madshus, I.H.; Olsnes, S.; Sandvig, K. Different pH requirements for entry of the two picornaviruses, human rhinovirus 2 and murine encephalomyocarditis virus. Virology 1984, 139, 346–357. [Google Scholar] [CrossRef]
- Crowther, D.; Melnick, J.L. The incorporation of neutral red and acridine orange into developing poliovirus particles making them photosensitive. Virology 1961, 14, 11–21. [Google Scholar] [CrossRef]
- Brandenburg, B.; Lee, L.Y.; Lakadamyali, M.; Rust, M.J.; Zhuang, X.; Hogle, J.M. Imaging poliovirus entry in live cells. PLoS Biol. 2007, 5, e183. [Google Scholar] [CrossRef] [PubMed]
- Kremser, L.; Petsch, M.; Blaas, D.; Kenndler, E. Labeling of capsid proteins and genomic RNA of human rhinovirus with two different fluorescent dyes for selective detection by capillary electrophoresis. Anal. Chem. 2004, 76, 7360–7365. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.G.; Lim, X.N.; Ng, W.C.; Sim, A.Y.L.; Poh, H.X.; Shen, Y.; Lim, S.Y.; Sundstrom, K.B.; Sun, X.; Aw, J.G.; et al. Structure mapping of dengue and Zika viruses reveals functional long-range interactions. Nat. Commun. 2019, 10, 1408. [Google Scholar] [CrossRef] [Green Version]
- Ziv, O.; Price, J.; Shalamova, L.; Kamenova, T.; Goodfellow, I.; Weber, F.; Miska, E.A. The Short- and Long-Range RNA-RNA Interactome of SARS-CoV-2. Mol. Cell 2020, 80, 1067–1077.e5. [Google Scholar] [CrossRef]
- Zhang, M.; Li, K.; Bai, J.; Velema, W.A.; Yu, C.; van Damme, R.; Lee, W.H.; Corpuz, M.L.; Chen, J.F.; Lu, Z. Optimized photochemistry enables efficient analysis of dynamic RNA structuromes and interactomes in genetic and infectious diseases. Nat. Commun. 2021, 12, 2344. [Google Scholar] [CrossRef]
- Broo, K.; Wei, J.; Marshall, D.; Brown, F.; Smith, T.J.; Johnson, J.E.; Schneemann, A.; Siuzdak, G. Viral capsid mobility: A dynamic conduit for inactivation. Proc. Natl. Acad. Sci. USA 2001, 98, 2274–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speir, J.A.; Johnson, J.E. Nucleic acid packaging in viruses. Curr. Opin. Struct. Biol. 2012, 22, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotecha, A.; Zhang, F.; Juleff, N.; Jackson, T.; Perez, E.; Stuart, D.; Fry, E.; Charleston, B.; Seago, J. Application of the thermofluor PaSTRy technique for improving foot-and-mouth disease virus vaccine formulation. J. Gen. Virol. 2016, 97, 1557–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez de Groot, N.; Armaos, A.; Grana-Montes, R.; Alriquet, M.; Calloni, G.; Vabulas, R.M.; Tartaglia, G.G. RNA structure drives interaction with proteins. Nat. Commun. 2019, 10, 3246. [Google Scholar] [CrossRef] [PubMed]
- Noller, H.F.; Hoffarth, V.; Zimniak, L. Unusual resistance of peptidyl transferase to protein extraction procedures. Science 1992, 256, 1416–1419. [Google Scholar] [CrossRef] [PubMed]
- Pascal, S.M.; Garimella, R.; Warden, M.S.; Ponniah, K. Structural Biology of the Enterovirus Replication-Linked 5′-Cloverleaf RNA and Associated Virus Proteins. Microbiol. Mol. Biol. Rev. 2020, 84, e00062-19. [Google Scholar] [CrossRef] [PubMed]
- Rohll, J.B.; Percy, N.; Ley, R.; Evans, D.J.; Almond, J.W.; Barclay, W.S. The 5′-untranslated regions of picornavirus RNAs contain independent functional domains essential for RNA replication and translation. J. Virol. 1994, 68, 4384–4391. [Google Scholar] [CrossRef] [Green Version]
- Lavezzo, E.; Berselli, M.; Frasson, I.; Perrone, R.; Palu, G.; Brazzale, A.R.; Richter, S.N.; Toppo, S. G-quadruplex forming sequences in the genome of all known human viruses: A comprehensive guide. PLoS Comput. Biol. 2018, 14, e1006675. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Qin, G.; Niu, J.; Wang, Z.; Wang, C.; Ren, J.; Qu, X. Targeting RNA G-Quadruplex in SARS-CoV-2: A Promising Therapeutic Target for COVID-19? Angew. Chem. Int. Ed. 2021, 60, 432–438. [Google Scholar] [CrossRef]
- Metifiot, M.; Amrane, S.; Litvak, S.; Andreola, M.L. G-quadruplexes in viruses: Function and potential therapeutic applications. Nucleic Acids Res. 2014, 42, 12352–12366. [Google Scholar] [CrossRef] [Green Version]
- Chaudhuri, R.; Bhattacharya, S.; Dash, J.; Bhattacharya, S. Recent Update on Targeting c-MYC G-Quadruplexes by Small Molecules for Anticancer Therapeutics. J. Med. Chem. 2021, 64, 42–70. [Google Scholar] [CrossRef] [PubMed]
- Dumas, L.; Herviou, P.; Dassi, E.; Cammas, A.; Millevoi, S. G-Quadruplexes in RNA Biology: Recent Advances and Future Directions. Trends Biochem. Sci. 2021, 46, 270–283. [Google Scholar] [CrossRef]
- Banco, M.T.; Ferre-D’Amare, A.R. The emerging structural complexity of G-quadruplex RNAs. RNA 2021, 27, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, E.; Richter, S.N. G-quadruplexes and G-quadruplex ligands: Targets and tools in antiviral therapy. Nucleic Acids Res. 2018, 46, 3270–3283. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Wu, Y.; Ou, T.M. RNA G-Quadruplex: The New Potential Targets for Therapy. Curr. Top. Med. Chem. 2015, 15, 1947–1956. [Google Scholar] [CrossRef]
- Schonrich, G.; Raftery, M.J. Neutrophil Extracellular Traps Go Viral. Front. Immunol. 2016, 7, 366. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Gorbatsevych, O.; Liu, Y.; Mugavero, J.; Shen, S.H.; Ward, C.B.; Asare, E.; Jiang, P.; Paul, A.V.; Mueller, S.; et al. Limits of variation, specific infectivity, and genome packaging of massively recoded poliovirus genomes. Proc. Natl. Acad. Sci. USA 2017, 114, E8731–E8740. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.R.; Papamichail, D.; Skiena, S.; Futcher, B.; Wimmer, E.; Mueller, S. Virus attenuation by genome-scale changes in codon pair bias. Science 2008, 320, 1784–1787. [Google Scholar] [CrossRef] [Green Version]
- Kikin, O.; D’Antonio, L.; Bagga, P.S. QGRS Mapper: A web-based server for predicting G-quadruplexes in nucleotide sequences. Nucleic Acids Res. 2006, 34, W676–W682. [Google Scholar] [CrossRef]
- Ogloblina, A.M.; Bannikova, V.A.; Khristich, A.N.; Oretskaya, T.S.; Yakubovskaya, M.G.; Dolinnaya, N.G. Parallel G-Quadruplexes Formed by Guanine-Rich Microsatellite Repeats Inhibit Human Topoisomerase I. Biochemistry 2015, 80, 1026–1038. [Google Scholar] [CrossRef] [PubMed]
- Piazza, A.; Cui, X.; Adrian, M.; Samazan, F.; Heddi, B.; Phan, A.T.; Nicolas, A.G. Non-Canonical G-quadruplexes cause the hCEB1 minisatellite instability in Saccharomyces cerevisiae. eLife 2017, 6, e26884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremser, L.; Okun, V.M.; Nicodemou, A.; Blaas, D.; Kenndler, E. Binding of fluorescent dye to genomic RNA inside intact human rhinovirus after viral capsid penetration investigated by capillary electrophoresis. Anal. Chem. 2004, 76, 882–887. [Google Scholar] [CrossRef]
- Draper, D.E. A guide to ions and RNA structure. RNA 2004, 10, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, D.; Mirihana Arachchilage, G.; Basu, S. Metal Cations in G-Quadruplex Folding and Stability. Front. Chem. 2016, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Wei, Y.; Mei, M.; Tang, L.; Sun, L.; Huang, W.; Zhou, J.; Zou, C.; Zhang, S.; Qin, C.F.; et al. Integrative Analysis of Zika Virus Genome RNA Structure Reveals Critical Determinants of Viral Infectivity. Cell Host Microbe 2018, 24, 875–886.e5. [Google Scholar] [CrossRef] [Green Version]
- Smyth, M.; Pettitt, T.; Symonds, A.; Martin, J. Identification of the pocket factors in a picornavirus. Arch. Virol. 2003, 148, 1225–1233. [Google Scholar] [CrossRef]
- Palika, A.; Armanious, A.; Rahimi, A.; Medaglia, C.; Gasbarri, M.; Handschin, S.; Rossi, A.; Pohl, M.O.; Busnadiego, I.; Gubeli, C.; et al. An antiviral trap made of protein nanofibrils and iron oxyhydroxide nanoparticles. Nat. Nanotechnol. 2021, 16, 1–8. [Google Scholar] [CrossRef]
- Graf, C.; Bernkop-Schnurch, A.; Egyed, A.; Koller, C.; Prieschl-Grassauer, E.; Morokutti-Kurz, M. Development of a nasal spray containing xylometazoline hydrochloride and iota-carrageenan for the symptomatic relief of nasal congestion caused by rhinitis and sinusitis. Int. J. Gen. Med. 2018, 11, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Neubauer, C.; Frasel, L.; Kuechler, E.; Blaas, D. Mechanism of entry of human rhinovirus 2 into HeLa cells. Virology 1987, 158, 255–258. [Google Scholar] [CrossRef]
- Berei, J.; Eckburg, A.; Miliavski, E.; Anderson, A.D.; Miller, R.J.; Dein, J.; Giuffre, A.M.; Tang, D.; Deb, S.; Racherla, K.S.; et al. Potential Telomere-Related Pharmacological Targets. Curr. Top. Med. Chem. 2020, 20, 458–484. [Google Scholar] [CrossRef] [PubMed]
- Kosiol, N.; Juranek, S.; Brossart, P.; Heine, A.; Paeschke, K. G-quadruplexes: A promising target for cancer therapy. Mol. Cancer 2021, 20, 40. [Google Scholar] [CrossRef]
- Cerofolini, L.; Amato, J.; Giachetti, A.; Limongelli, V.; Novellino, E.; Parrinello, M.; Fragai, M.; Randazzo, A.; Luchinat, C. G-triplex structure and formation propensity. Nucleic Acids Res. 2014, 42, 13393–13404. [Google Scholar] [CrossRef]
- Bonnat, L.; Dautriche, M.; Saidi, T.; Revol-Cavalier, J.; Dejeu, J.; Defrancq, E.; Lavergne, T. Scaffold stabilization of a G-triplex and study of its interactions with G-quadruplex targeting ligands. Org. Biomol. Chem. 2019, 17, 8726–8736. [Google Scholar] [CrossRef] [PubMed]
- Shyr, Z.A.; Cheng, Y.S.; Lo, D.C.; Zheng, W. Drug combination therapy for emerging viral diseases. Drug Discov. Today 2021. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | PDB (Virus–Inhibitor Complex) | Clinical Trials Register | Reference |
|---|---|---|---|
| BTA798 (Vapendavir) | 3VDD (RVA2) | NCT03024177 (w); NCT02367313 (nrp); 2014-001785-95 (cwr) | [69] |
| OBR-5-340 (LGQ) | 6SK5 (RVB5) | - | [40] |
| R77975 (Pirodavir) | 1PO2 (PV1); 1VBC (PV3) | - | [70,71,72] |
| SCH48973 (Pocapavir) | 1EAH (PV2L) | - | [73] |
| Win-51711 (Disoxaril) | 1D4M (CVA9); 1PIV (PV3); 3ZFE (EV71) | - | [35,74,75] |
| WIN-61209 (Isoxazole) | 1QJU (RVA16) | - | [76] |
| Win-63843 (Pleconaril) | 1C8M (RVA16); 1NCQ (RVB14), 1NCR (RVA16); 4WM7 (EVD68) | NCT00394914 (cwr) | [67,77,78] |
| Compound 17 | 6GZV (CVB3) | - | [41] |
| Target/Compound | Reference |
|---|---|
| 2Apro | |
| Hinokitiol | [90] |
| Pyrrolidine dithiocarbamate | [91] |
| Elastatinal | [92] |
| Z-VAD-FMK | [93] |
| 2CATPase | |
| (S)-fluoxetine | [94] |
| Dibucaine | [95] |
| Pirlindole | [95] |
| Zuclopenthixol | [95] |
| Guanidine hydrochloride | [17] |
| 3A | |
| Enviroxime | [96] |
| 3C(D)Pro | |
| Rupintrivir | [97,98] |
| SG85 | [99] |
| 3DPol | |
| Ribavirin | [100,101] |
| Favipiravir (T-705) | [102] |
| Gliotoxin | [103] |
| Compound | Proposed Mechanism | Clinical Trials Register | Reference |
|---|---|---|---|
| Lactoferrin | Sulfated glucosaminoglycan (sGAG) blocker | NCT01677702 (nrp); NCT01092039 (nrp) | [131,132,133,134,135] |
| Nanoparticles | Heparan sulfate proteoglycans mimetic Virion aggregation | - | [23] |
| PDS fibrils | Virion entrapment | [25] | |
| Carrageenans | Virion entrapment | NCT01944631 (cwr); NCT04533906 (nrp) | [136,137] |
| LEI110 (*) | PLA2G16 inhibitor | - | [117] |
| Geldanamycin | HSP90 inhibitor | - | [119,138] |
| Pochonin D | HSP90 inhibitor | - | [122] |
| Wortmannin | PI 3-kinase inhibition | - | [139] |
| Dynasore | Dynamin GTPase inhibitor | - | [140] |
| Blebbistatin | Myosin ATPase activity inhibitor | - | [140] |
| Chlorpromazine | Prevent the assembly/disassembly of clathrin lattices | - | [140] |
| Filipin | Cholesterol sequestering | - | [140] |
| Itraconazole | Oxysterol binding protein inhibitor | - | [141] |
| Monensin | Endosome acidification inhibitor | - | [142] |
| Niclosamide | Endosome acidification inhibitor | - | [143] |
| EIPA | Endosome acidification inhibitor | - | [144] |
| Bafilomycin A | Endosome acidification inhibitor | - | [145] |
| Ammonium Chloride | Endosome acidification inhibitor | - | [146] |
| Cytochalasin D (CytD) | Actin-polymerisation inhibitor | - | [145] |
| Budesonide | Autophagy activator | - | [147] |
| DDD85646 | N-myristoyltransferase inhibitor | - | [124] |
| ML-9 | Myosin light chain kinase inhibitor | - | [148] |
| sMBP-V33333 | Receptor mimetic | - | [149] |
| sICAM-1 | Receptor mimetic | - | [150,151,152] |
| Dendrimers | Blocking receptor attachment of EV-A71 and HIV | - | [24,153] |
| Deoxyglucose | Metabolism modifier | - | [154] |
| Azithromycin | Immunomodulatory | - | [155] |
| LL-37 | Cathelicidins | - | [156] |
| PDS | Targeting vRNA | - | § |
| PhenDC3 | Targeting vRNA | - | § |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Real-Hohn, A.; Blaas, D. Rhinovirus Inhibitors: Including a New Target, the Viral RNA. Viruses 2021, 13, 1784. https://doi.org/10.3390/v13091784
Real-Hohn A, Blaas D. Rhinovirus Inhibitors: Including a New Target, the Viral RNA. Viruses. 2021; 13(9):1784. https://doi.org/10.3390/v13091784
Chicago/Turabian StyleReal-Hohn, Antonio, and Dieter Blaas. 2021. "Rhinovirus Inhibitors: Including a New Target, the Viral RNA" Viruses 13, no. 9: 1784. https://doi.org/10.3390/v13091784
APA StyleReal-Hohn, A., & Blaas, D. (2021). Rhinovirus Inhibitors: Including a New Target, the Viral RNA. Viruses, 13(9), 1784. https://doi.org/10.3390/v13091784