
Impact of Molecular Modification on the Efficiency of Recombinant Baculovirus Vector Invasion to Mammalian Cells and Its Immunogenicity in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, Viral Bacmid, Reagents, Cells, and Mice
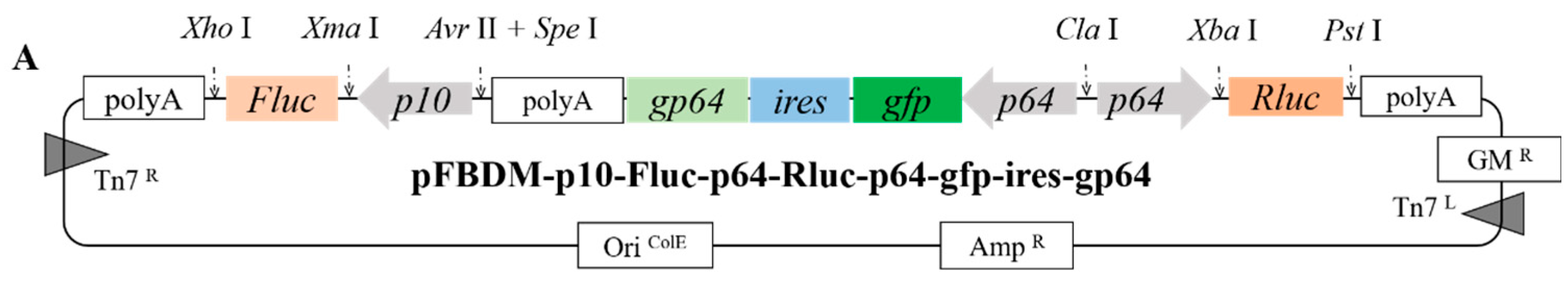
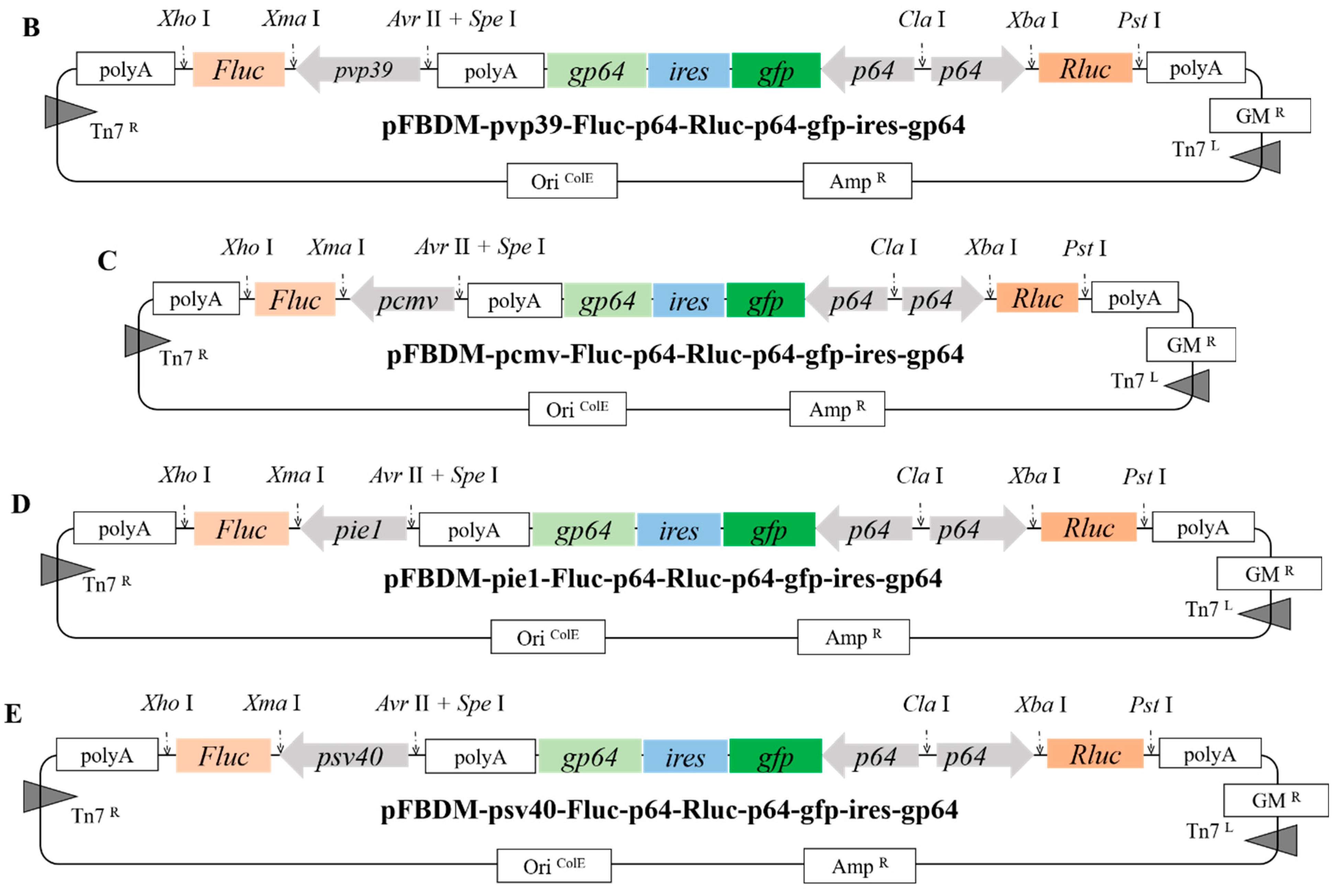
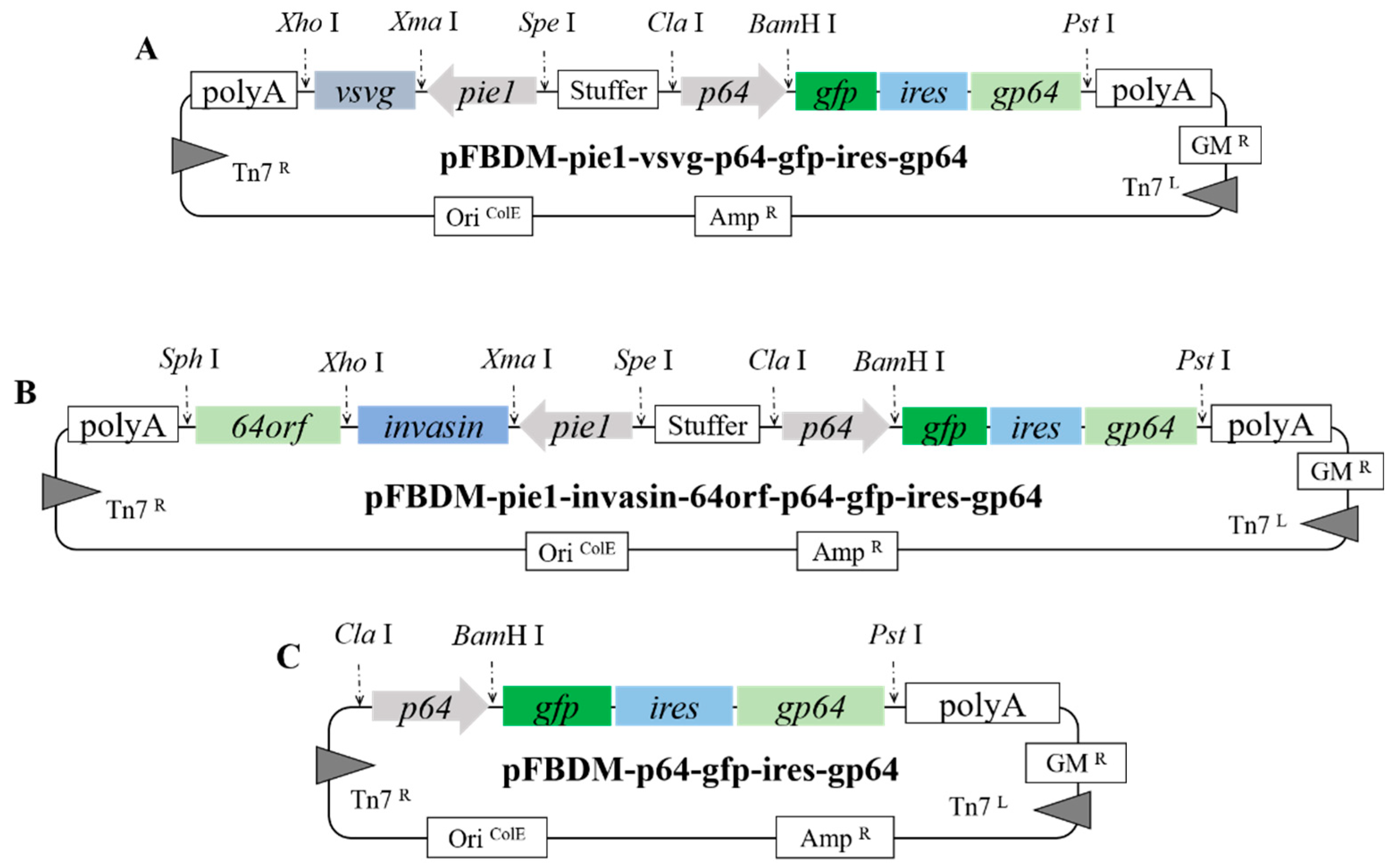
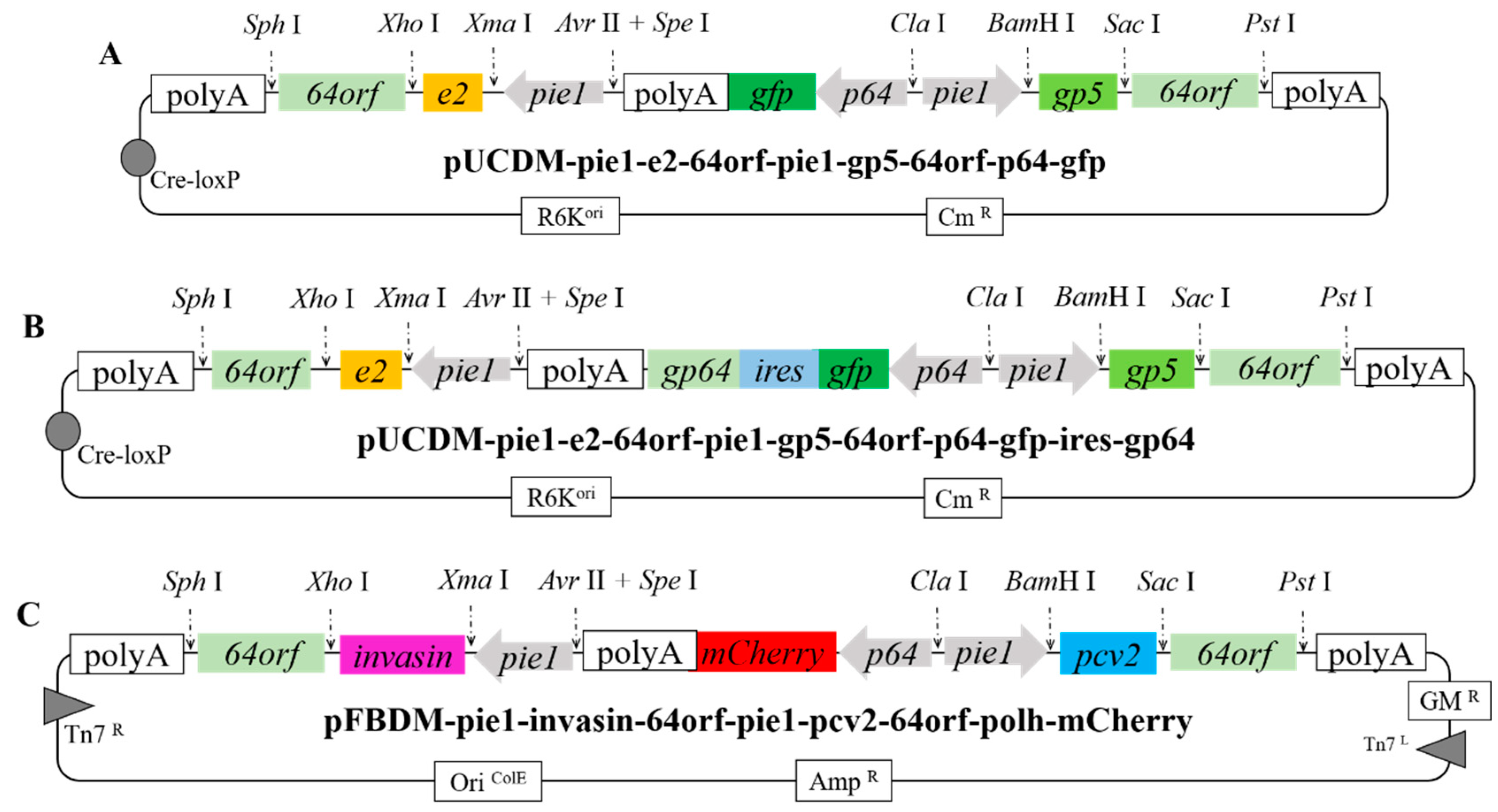
2.2. Construction of Donor Vectors
2.3. Introduction of Foreign Genes into rBacmid
2.4. Production of Recombinant Baculovirus
2.5. Determination of Recombinant Baculovirus Titer
2.6. Baculovirus Purification and Transmission Electron Microscopy (TEM)
2.7. Dual-Glo Luciferase Assay System
2.8. ELISA
2.9. Statistical Analysis
3. Results
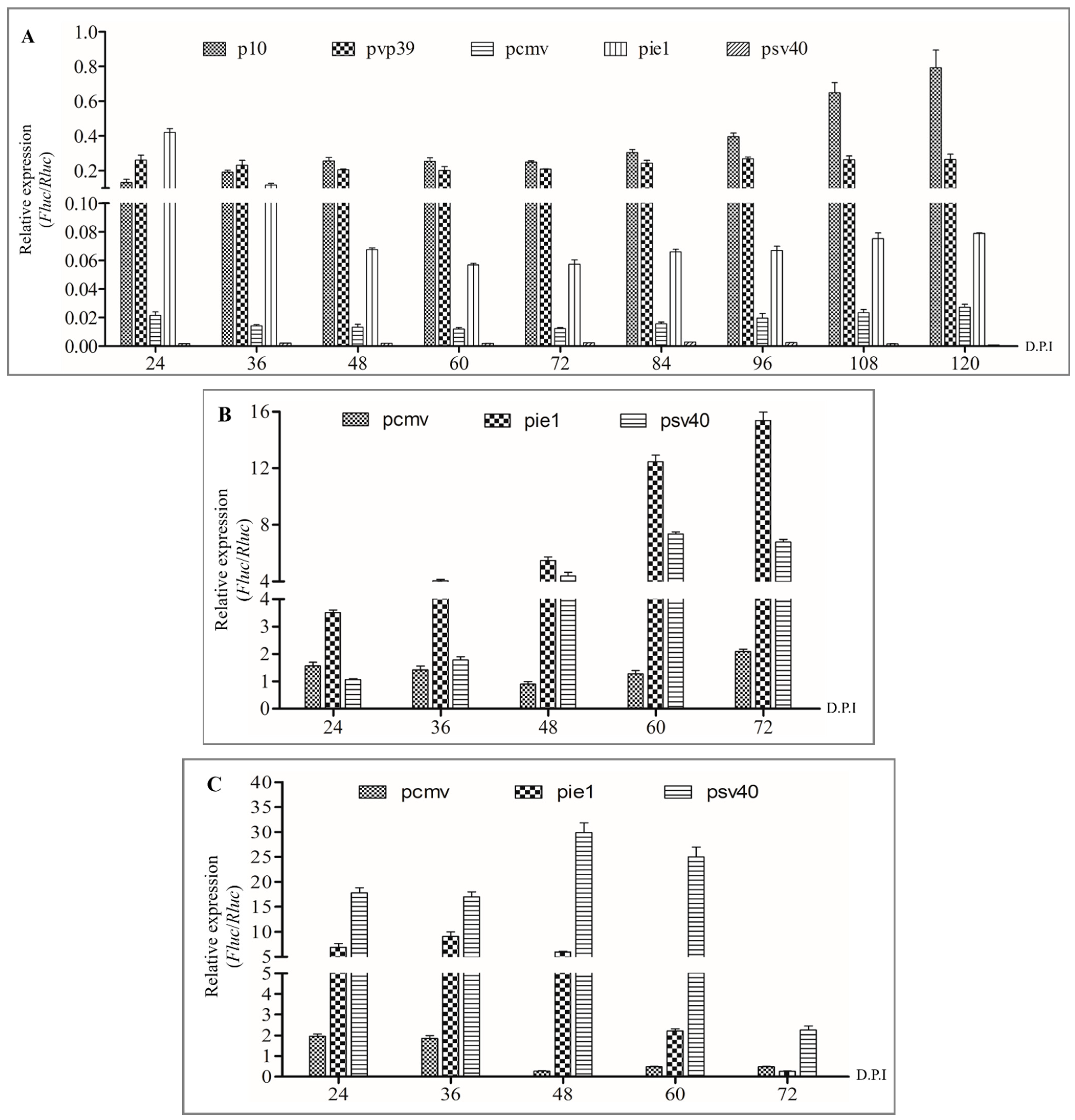
3.1. Determination of Promoter Expression in Insect and Mammalian Cells
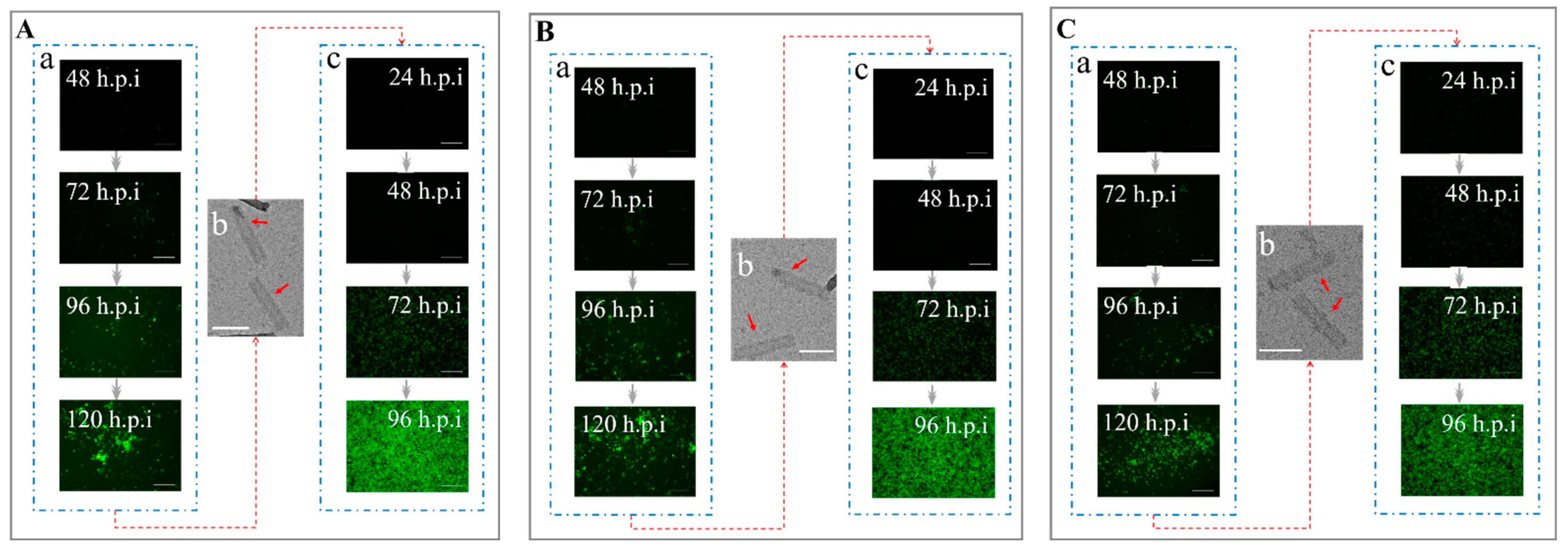
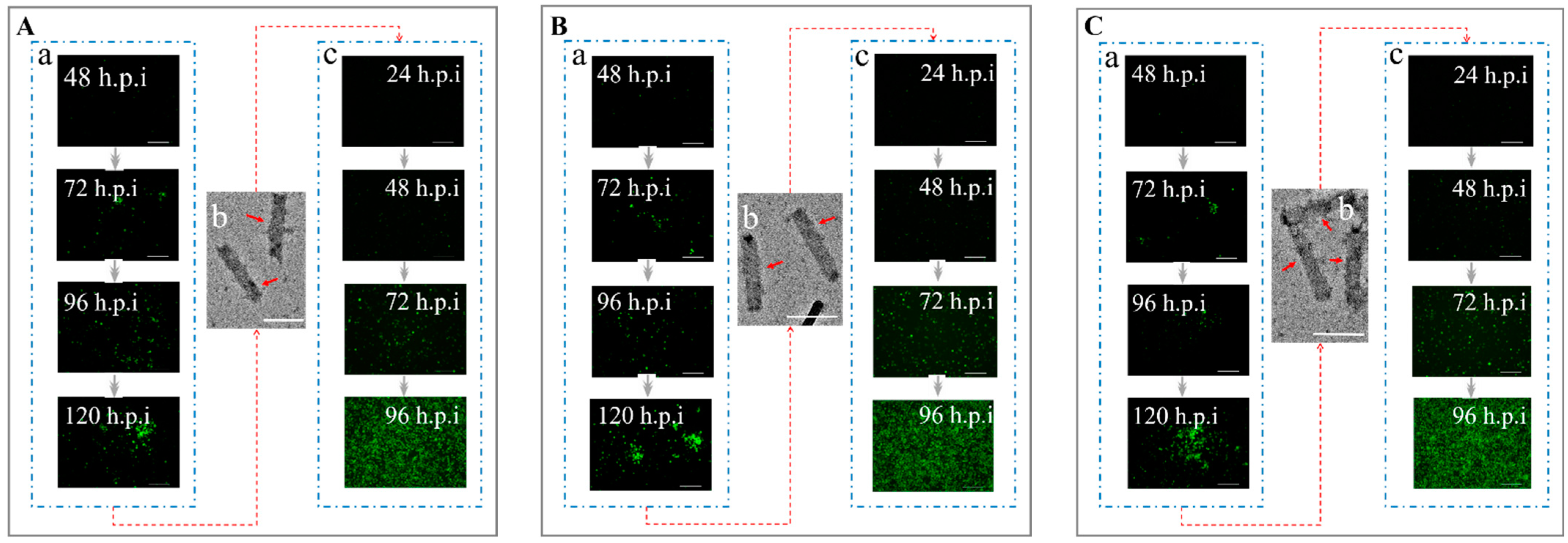
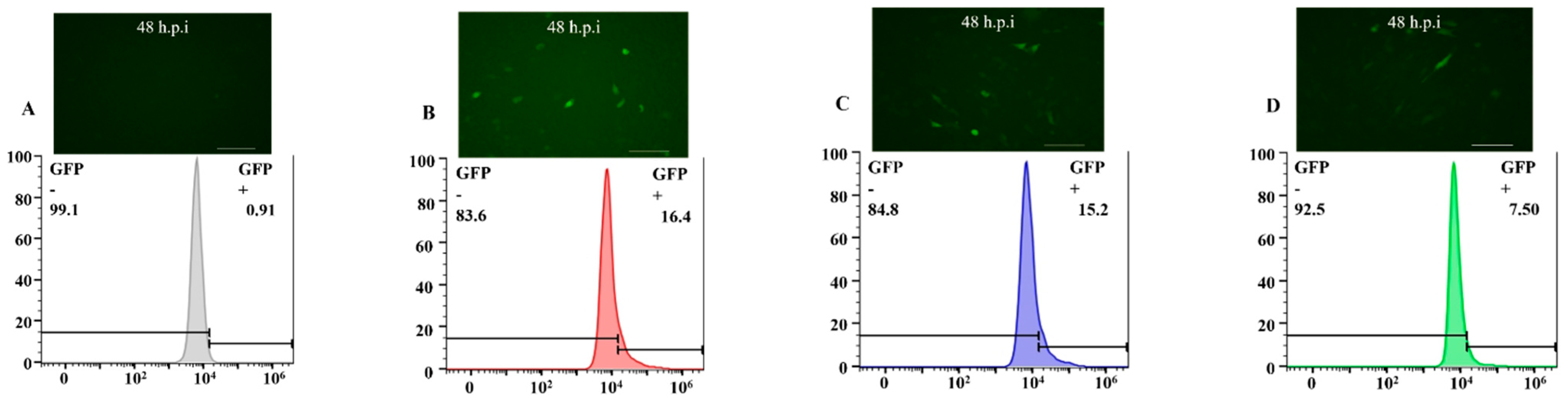
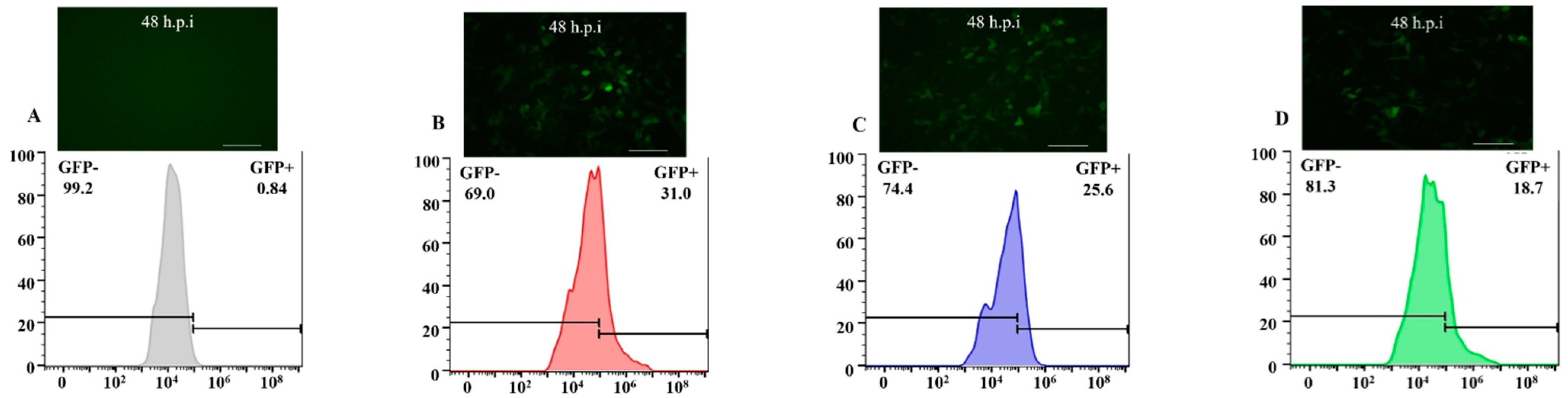
3.2. The Efficiency of Transducing Mammalian Cells by BV with Envelope Modification
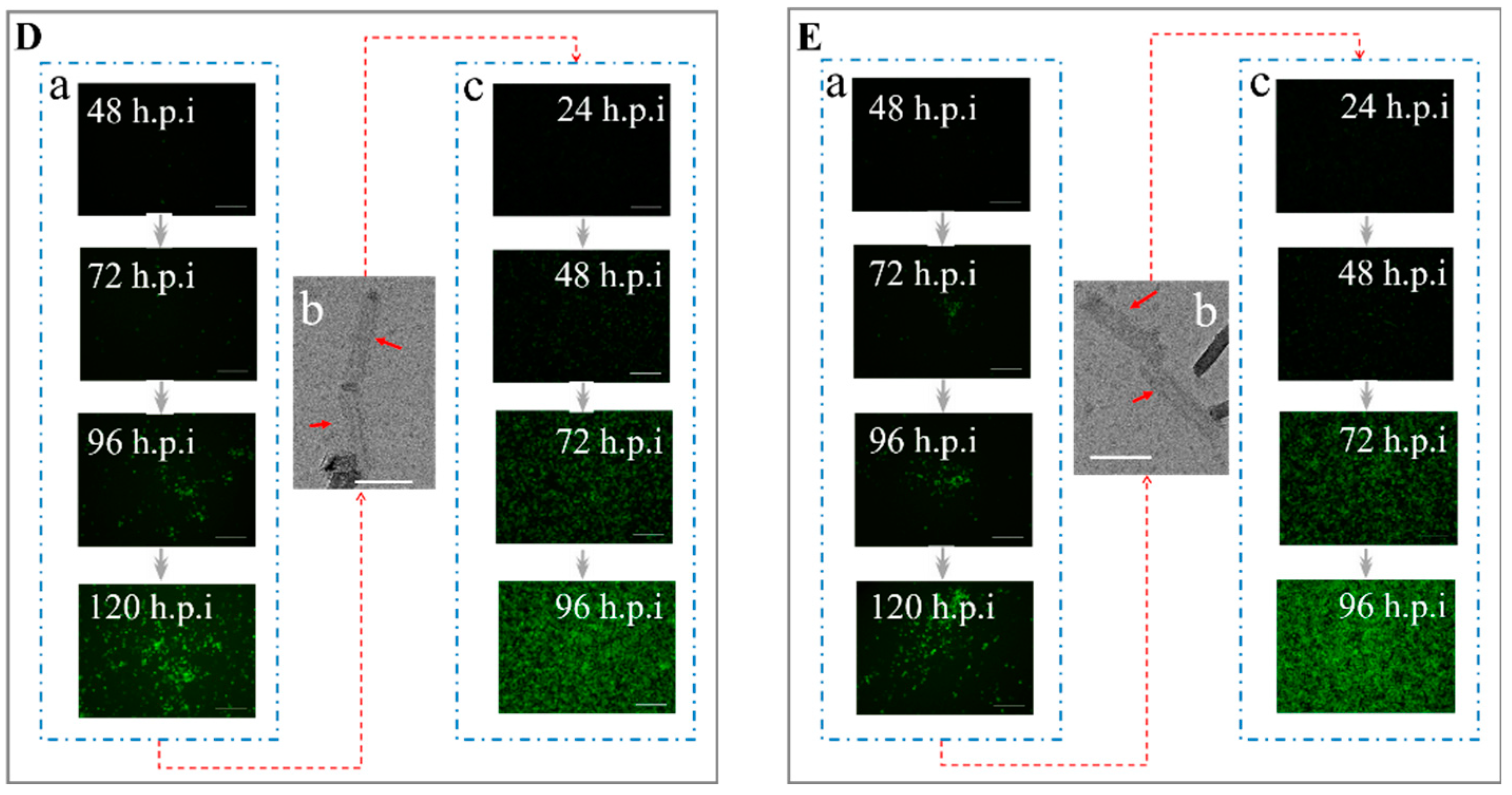
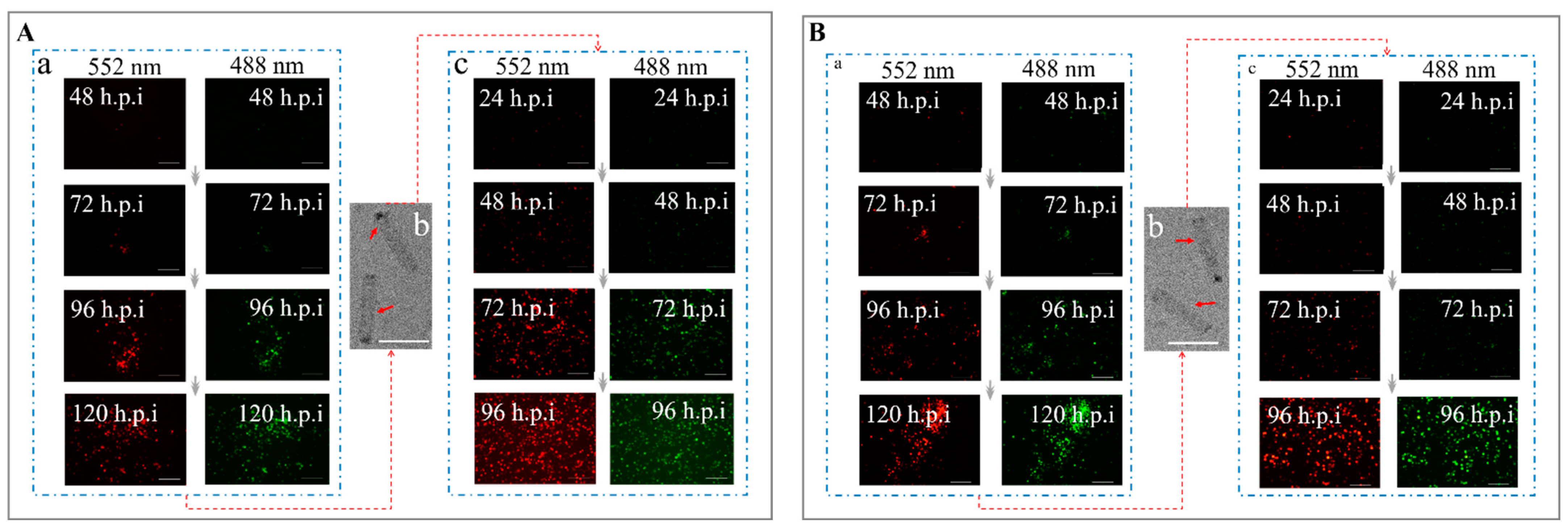
3.3. Determination of the Co-Display Efficiency of Multiple Bacmid-Δgp64 Genes
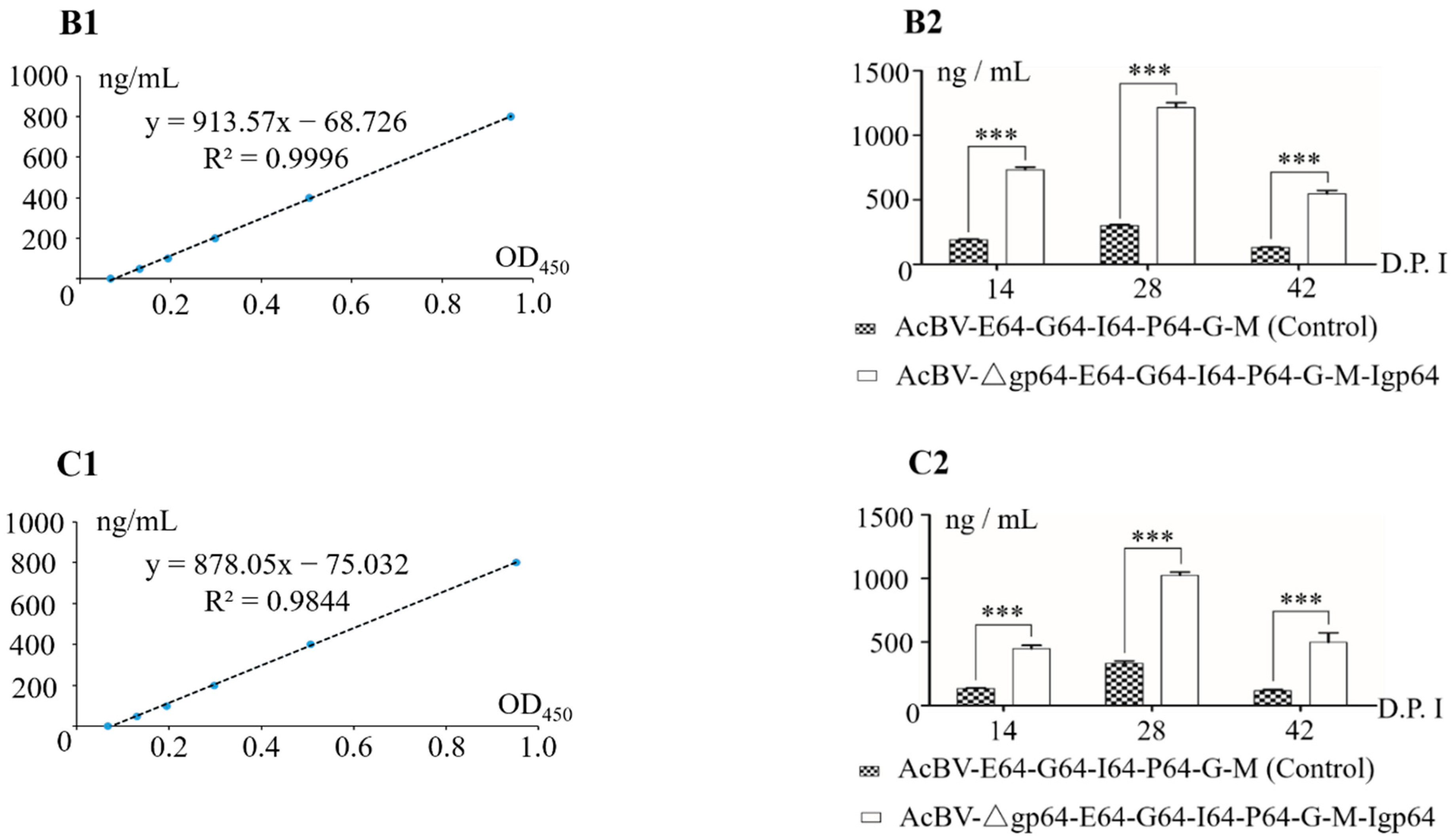
3.4. Determination of Immunogenicity of Multiple-Vaccines Encoded by Bacmid-Δgp64
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdulrahman, W.; Radu, L.; Garzoni, F.; Kolesnikova, O.; Gupta, K.; Osz-Papai, J.; Berger, I.; Poterszman, A. The production of multiprotein complexes in insect cells using the baculovirus expression system. Methods Mol. Biol. 2015, 1261, 91–114. [Google Scholar]
- Lynn, D.E. Comparative susceptibilities of insect cell lines to infection by the occlusion-body derived phenotype of baculoviruses. J. Invertebr. Pathol. 2003, 83, 215–222. [Google Scholar] [CrossRef]
- Shin, H.; Choi, H.; Kim, N.; Park, N.; Kim, H.; Kim, J.; Kim, Y. Unraveling the genome-wide impact of recombinant baculovirus infection in mammalian cells for gene delivery. Genes 2020, 11, 1306. [Google Scholar] [CrossRef]
- Altmannova, V.; Blaha, A.; Astrinidis, S.; Reichle, H.; Weir, J.R. InteBac: An integrated bacterial and baculovirus expression vector suite. Protein Sci. 2021, 30, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Miao, Y.; Ke, X.; Tan, Z.; Hu, C.; Li, P.; Wang, T.; Zhang, Y.; Sun, J.; Liu, Y.; et al. Baculovirus surface display of zika virus envelope protein protects against virus challenge in mouse model. Virol. Sin. 2020, 35, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Jian, C.; Na, L.; Wang, X.; Dai, Y.; Li, D. Production and characterization of Hantaan virus-like particles from baculovirus expression system. Biochem. Eng. J. 2019, 152, 107373. [Google Scholar] [CrossRef]
- Scholefield, J.A.; Shikano, I.; Lowenberger, C.A.; Cory, J.S. The impact of baculovirus challenge on immunity: The effect of dose and time after infection. J. Invertebr. Pathol. 2019, 167, 107232. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, X.; Ren, F.; Zou, S.; Feng, M.; Xu, L.; Yao, L.; Sun, J. Construction of a highly efficient display system for baculovirus and its application on multigene co-display. Mol. Genet. Genom. 2018, 293, 1265–1277. [Google Scholar] [CrossRef] [PubMed]
- Molinari, P.; Molina, G.N.; Tavarone, E.; Zajac, M.P.D.M.; Morón, G.; Taboga, O. Baculovirus capsid display in vaccination schemes: Effect of a previous immunity against the vector on the cytotoxic response to delivered antigens. Appl. Microbiol. Biotechnol. 2018, 102, 10139–10146. [Google Scholar] [CrossRef]
- Premanand, B.; Wee, P.Z.; Prabakaran, M. Baculovirus surface display of immunogenic proteins for vaccine development. Viruses 2018, 10, 298. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Blissard, G.W.; Liu, T.-X.; Li, Z. Autographa californica multiple nucleopolyhedrovirus GP64 protein: Analysis of domain I and V amino acid interactions and membrane fusion activity. Virology 2016, 488, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Yin, F.; Shen, S.; Tan, Y.; Deng, F.; Vlak, J.M.; Hu, Z.; Wang, H. partial functional rescue of helicoverpa armigera single nucleocapsid nucleopolyhedrovirus infectivity by replacement of F protein with GP64 from autographa californica multicapsid nucleopolyhedrovirus. J. Virol. 2010, 84, 11505–11514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.Y.; Chen, Y.H.; Liu, H.J. Baculovirus as a vaccine vector. Bioengineered 2012, 3, 271–274. [Google Scholar] [CrossRef]
- Charlton, C.A.; Volkman, L.E. Penetration of autographa californica nuclear polyhedrosis virus nucleocapsids into IPLB Sf 21 cells induces actin cable formation. Virology 1993, 197, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, C.; Kaname, Y.; Taguwa, S.; Abe, T.; Fukuhara, T.; Tani, H.; Moriishi, K.; Matsuura, Y. Baculovirus GP64-mediated entry into mammalian cells. J. Virol. 2011, 86, 2610–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hefferon, K.; Oomens, A.; Monsma, S.; Finnerty, C.; Blissard, G. Host cell receptor binding by baculovirus gp64 and kinetics of virion entry. Virology 1999, 258, 455–468. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Cao, L.; Miao, Y.; Wu, S.; Xu, S.; Wang, R.; Du, J.; Liang, A.; Fu, Y. Transcriptome analysis of Spodoptera frugiperda 9 (Sf9) cells infected with baculovirus, AcMNPV or AcMNPV-BmK IT. Biotechnol. Lett. 2017, 39, 1129–1139. [Google Scholar] [CrossRef]
- Sun, X.; Yau, V.K.; Briggs, B.J.; Whittaker, G.R. Role of clathrin-mediated endocytosis during vesicular stomatitis virus entry into host cells. Virology 2005, 338, 53–60. [Google Scholar] [CrossRef]
- Hu, L.; Li, Y.; Deng, F.; Hu, Z.; Wang, H.; Wang, M. Improving baculovirus transduction of mammalian cells by incorporation of thogotovirus glycoproteins. Virol. Sin. 2019, 34, 454–466. [Google Scholar] [CrossRef]
- Carneiro, F.A.; Vandenbussche, G.; Juliano, M.A.; Juliano, L.; Ruysschaert, J.-M.; Da Poian, A.T. Charged residues are involved in membrane fusion mediated by a hydrophilic peptide located in vesicular stomatitis virus G protein. Mol. Membr. Biol. 2006, 23, 396–406. [Google Scholar] [CrossRef]
- Sun, J.; Yao, L.; Yao, N.; Xu, H.; Jin, P.; Kan, Y. Production of recombinant Bombyx mori nucleopolyhedrovirus in silkworm by intrahaemocoelic injection with invasive diaminopimelate auxotrophic Escherichia coli containing BmNPV-Bacmid. Biotechnol. Appl. Biochem. 2010, 57, 117–125. [Google Scholar] [CrossRef]
- Yao, L.; Wang, S.; Su, S.; Yao, N.; He, J.; Peng, L.; Sun, J. Construction of a baculovirus-silkworm multigene expression system and its application on producing virus-like particles. PLoS ONE 2012, 7, e32510. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.G.; Sun, J.C.; Xu, H.; Kan, Y.C.; Zhang, X.M.; Yan, H.C. A novel economic method for high throughput production of recombinant baculovirus by infecting insect cells with bacmid-containing diminopimelate-auxotrophic Escherichia coli. J. Biotechnol. 2010, 145, 23–29. [Google Scholar] [CrossRef]
- Sun, J.C.; Zhang, E.H.; Yao, L.G.; Zhang, H.L.; Jin, P.F. A high efficient method of constructing recombinant Bombyx mori(silkworm) multiple nucleopolyhedrovirus based on zero-background Tn7-mediated transposition in Escherichia coli. Biotechnol. Prog. 2009, 25, 524–529. [Google Scholar] [CrossRef]
- Liu, Z.C.; Yao, L.G.; Zhang, X.M.; Kan, Y.C.; Zhou, J.-J. A highly efficient method for the generation of a recombinant Bombyx mori nuclear-polyhedrosis-virus Bacmid and large-scale expression of foreign proteins in silkworm (B. mori) larvae. Biotechnol. Appl. Biochem. 2007, 48, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, H.; Xu, Z.; Ishii-Watabe, A.; Uchida, E.; Hayakawa, T. IRES-dependent second gene expression is significantly lower than cap-dependent first gene expression in a bicistronic vector. Mol. Ther. 2000, 1, 376–382. [Google Scholar] [CrossRef]
- Nagai, S.; Alves, C.A.F.; Kobayashi, M.; Ikeda, M. Comparative transient expression assay analysis of hycu-hr6- and IE1-dependent regulation of baculovirus gp64 early promoters in three insect cell lines. Virus Res. 2011, 155, 83–90. [Google Scholar] [CrossRef]
- Liu, W.-J.; Chang, Y.-S.; Wang, C.-H.; Kou, G.-H.; Lo, C.-F. Microarray and RT-PCR screening for white spot syndrome virus immediate-early genes in cycloheximide-treated shrimp. Virology 2005, 334, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-J.; Chang, Y.-S.; Wang, A.H.-J.; Kou, G.-H.; Lo, C.-F. White spot syndrome virus annexes a shrimp stat to enhance expression of the immediate-early gene ie1. J. Virol. 2007, 81, 1461–1471. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; McCormick, C.; Arthur, M.J.P.; Ruddell, R.; Oakley, F.; Smart, D.; Murphy, F.R.; Harris, M.P.G.; Mann, D.A. High efficiency gene transfer into cultured primary rat and human hepatic stellate cells using baculovirus vectors. Liver Int. 2002, 22, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Long, G.; Pan, X.; Kormelink, R.; Vlak, J.M. Functional entry of baculovirus into insect and mammalian cells is dependent on clathrin-mediated endocytosis. J. Virol. 2006, 80, 8830–8833. [Google Scholar] [CrossRef] [Green Version]
- Tani, H.; Nishijima, M.; Ushijima, H.; Miyamura, T.; Matsuura, Y. Characterization of cell-surface determinants important for baculovirus infection. Virology 2001, 279, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Wang, M.; Qiu, Z.; Deng, F.; Vlak, J.M.; Hu, Z.; Wang, H. Autographa californica multicapsid nucleopolyhedrovirus efficiently infects Sf9 cells and transduces mammalian cells via direct fusion with the plasma membrane at low pH. J. Virol. 2010, 84, 5351–5359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yali, C.; Yang, Y.; Xu, C.; Shi, L. Vesicular stomatitis virus G protein transmembrane region is crucial for the hemi-fusion to full fusion transition. Sci. Rep. 2018, 8, 10669. [Google Scholar]
- Ghandehari, F.; Behbahani, M.; Pourazar, A.; Nourmohammadi, Z. Producing vesicular stomatitis virus g (VSVG) protein and assessment of its cytotoxic activity against breast cancer cells. J. Isfahan Med. Sch. 2015, 33, 221–230. [Google Scholar]
- Yang, D.G.; Chung, Y.C.; Lai, Y.K.; Lai, C.W.; Liu, H.J.; Hu, Y.C. Avian influenza virus hemagglutinin display on baculovirus envelope: Cytoplasmic domain affects virus properties and vaccine potential. Mol. Ther. 2007, 15, 989–996. [Google Scholar] [CrossRef]
- Chapple, S.; Jones, I.M. Non-polar distribution of green fluorescent protein on the surface of Autographa californica nucleopolyhedrovirus using a heterologous membrane anchor. J. Biotechnol. 2002, 95, 269–275. [Google Scholar] [CrossRef]
- Tao, Y.; Li, G.; Zheng, W.; Shu, J.; Chen, J.; Yang, F.; Wu, Y.; He, Y. Development of a combined genetic engineering vaccine for porcine circovirus type 2 and mycoplasma hyopneumoniae by a baculovirus expression system. Int. J. Mol. Sci. 2019, 20, 4425. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Li, Z.; Li, J.; Yi, Y.; Zhang, Y.; Li, X.; Li, B.; Yang, B.; Lan, X.; Li, Y.; et al. Rabies virus nucleoprotein expressed in silkworm pupae at high-levels and evaluation of immune responses in mice. J. Biotechnol. 2013, 163, 333–338. [Google Scholar] [CrossRef]
- Prabakaran, M.; Madhan, S.; Prabhu, N.; Geng, G.Y.; New, R.; Kwang, J. Reverse micelle-encapsulated recombinant baculovirus as an oral vaccine against H5N1 infection in mice. Antivir. Res. 2010, 86, 180–187. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, Y.; Zhang, Y.; Yang, Y.; Ren, J.; Zhang, X.; Du, E. Surface displaying of swine IgG1 Fc enhances baculovirus-vectored vaccine efficacy by facilitating viral complement escape and mammalian cell transduction. Vet. Res. 2017, 48, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, C.; Strauss, M. Baculovirus-mediated gene transfer in the presence of human serum or blood facilitated by inhibition of the complement system. Gene Ther. 1998, 5, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Georgopoulos, L.J.; Elgue, G.; Sanchez, J.; Dussupt, V.; Magotti, P.; Lambris, J.D.; Tötterman, T.H.; Maitland, N.J.; Nilsson, B. Preclinical evaluation of innate immunity to baculovirus gene therapy vectors in whole human blood. Mol. Immunol. 2009, 46, 2911–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaname, Y.; Tani, H.; Kataoka, C.; Shiokawa, M.; Taguwa, S.; Abe, T.; Moriishi, K.; Kinoshita, T.; Matsuura, Y. Acquisition of complement resistance through incorporation of CD55/decay-accelerating factor into viral particles bearing baculovirus GP64. J. Virol. 2010, 84, 3210–3219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martyn, J.C.; Cardin, A.J.; Wines, B.D.; Cendron, A.; Li, S.; MacKenzie, J.; Powell, M.; Gowans, E.J. Surface display of IgG Fc on baculovirus vectors enhances binding to antigen-presenting cells and cell lines expressing Fc receptors. Arch. Virol. 2009, 154, 1129–1138. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | Primer Sequences | Genes |
|---|---|---|
| Rluc | F: 5′-tctagaatgatgtacatagaagatt-3′ (Xba I) R: 5′-ctgcagttacacttgatacatgc-3′ (Pst I) | Renilla-luciferase |
| Fluc | F: 5′-cccgggatggaagacgccaaaaacataaagaa-3′ (Xma I) R: 5′-ctcgagttacacggcgatctttccgcccttctt-3′ (Xho I) | Firefly-luciferas |
| p10 | F: 5′-actagtatacggacctttaattcaacccaac-3′ (Spe I) R: 5′-cccgggtgatcaagtcttcgtcgagtgatt-3′ (Xma I) | promoter of baculovirus |
| pvp39 | F: 5′-actagttcgcgagttttgcagcgtctctgaa-3′ (Spe I) R: 5′-cccgggggatccttgttgccgttataaatatg-3′ (Xma I) | promoter of baculovirus |
| pcmv | F: 5′-actagtgttgacattgattattgtctagtta-3′(Spe I) R: 5′-cccgggtctgcttatatagacctcccaccgta-3′ (Xma I) | promoter of baculovirus |
| pie1 | F: 5′-actagttcaattttatgtggctaatggagaat-3′ (Spe I) R: 5′-cccgggcttgagtggagagagagagctagtta-3′ (Xma I) | promoter of white spot syndrome virus |
| psv40 | F: 5′-actagtattcaaatatgtatccgctcatgagac-3′ (Spe I) R: 5′-cccgggcctccaaaaaagcctcctcactactt-3′ (Xma I) | promoter of Simian vacuolating virus 40 |
| vsvg | F: 5′-cccgggatggtaagcgctattgttttatatgtgcttttggcggcggcggcgcattctgcctttgc ggcgggatctaagttcaccatagttt-3′ (Xma I) R: 5′-ctcgagttactttccaagtcggttcatctcta-3′ (Xho I) | envelope gene of vesicular stomatitis virus |
| invasin | F: 5′-cccgggatggtaagcgctattgttttatatgtgcttttggcggcggcggcgcattctgcctttgc ggcgatggataacgatgttgctaataata-3′ (Xma I) R: 5′-ctcgagtattgccagcgcacagagcgggaacg-3′ (Xho I) | infection gene of Yersinia pseudotuberculosis |
| E2 | F: 5′-cccgggatggtaagcgctattgttttatatgtgcttttggcggcggcggcgcattctgcctttgc ggcgcggctagcctgcaaggaaga-3′ (Xma Ⅰ) R: 5′-ctcgaggtagaatagatcttcattttccact-3′ (Xho Ⅰ) | envelope gene of classical swine fever virus |
| gp5 | F: 5′-ggatccatggtaagcgctattgttttatatgtgcttttggcggcggcggcgcattctgcctttgc ggcgttggggaagtgcttgaccgcgt-3′ (BamH Ⅰ) R: 5′-gagctcgagacgaccccattgttccgctgaaa-3′ (Sac I) | envelope gene of porcine reproductive and respiratory syndrome virus |
| pcv2 | F: 5′-ggatccatggtaagcgctattgttttatatgtgcttttggcggcggcggcgcattctgcctttgc ggcgacgtatccaaggaggcgttacc-3′ (BamH I) R: 5′-gagctcttcattaagggttaagttgggggtct-3′ (Sac Ⅰ) | envelope gene of porcine circovirus type 2 |
| 64ORF | F: 5′-gagctcctcgaggagcactgcaacgcgcaaatgaag-3′ (Sac I + Xho I) R: 5′-ctgcaggcatgcttaatattgtctattacggtttctaa-3′ (Pst I + Sph I) | baculoviral envelope gene |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.; Pan, Y.; Wang, X.; Tian, W.; Yao, L.; Sun, J. Impact of Molecular Modification on the Efficiency of Recombinant Baculovirus Vector Invasion to Mammalian Cells and Its Immunogenicity in Mice. Viruses 2022, 14, 140. https://doi.org/10.3390/v14010140
Zheng H, Pan Y, Wang X, Tian W, Yao L, Sun J. Impact of Molecular Modification on the Efficiency of Recombinant Baculovirus Vector Invasion to Mammalian Cells and Its Immunogenicity in Mice. Viruses. 2022; 14(1):140. https://doi.org/10.3390/v14010140
Chicago/Turabian StyleZheng, Hao, Yong Pan, Xiong Wang, Weibin Tian, Lunguang Yao, and Jingchen Sun. 2022. "Impact of Molecular Modification on the Efficiency of Recombinant Baculovirus Vector Invasion to Mammalian Cells and Its Immunogenicity in Mice" Viruses 14, no. 1: 140. https://doi.org/10.3390/v14010140
APA StyleZheng, H., Pan, Y., Wang, X., Tian, W., Yao, L., & Sun, J. (2022). Impact of Molecular Modification on the Efficiency of Recombinant Baculovirus Vector Invasion to Mammalian Cells and Its Immunogenicity in Mice. Viruses, 14(1), 140. https://doi.org/10.3390/v14010140