
Nectin-2 Acts as a Viral Entry Mediated Molecule That Binds to Human Herpesvirus 6B Glycoprotein B

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus Strains
2.2. Reverse Transcription-PCR (RT-PCR) and Quantitative RT-PCR
2.3. Antibodies
2.4. Flow Cytometric Analysis
2.5. Immunofluorescence Assay (IFA)
2.6. Establishment of a CD134-Membrane Unanchored HSY Cell Clone (Ex6 C7)
2.7. Establishment of Nectin-2 Knockout Ex6 C7 (Ex6 C7 Nec2-KO) Cell Clones
2.8. Expression Screening
2.9. Generation of Nectin-2- and CD134-Expressing Cell Lines
2.10. Generation of Nectin-2- and CD134-Expressing Cell Lines
2.11. Transfection, Immunoblotting, and Co-IP
2.12. Nectin-2 Binding Assay in HHV-6B-Infected Cells
2.13. Assay of Nectin-2 Binding to Purified HHV-6B gB
2.14. Statistical Analysis
3. Results
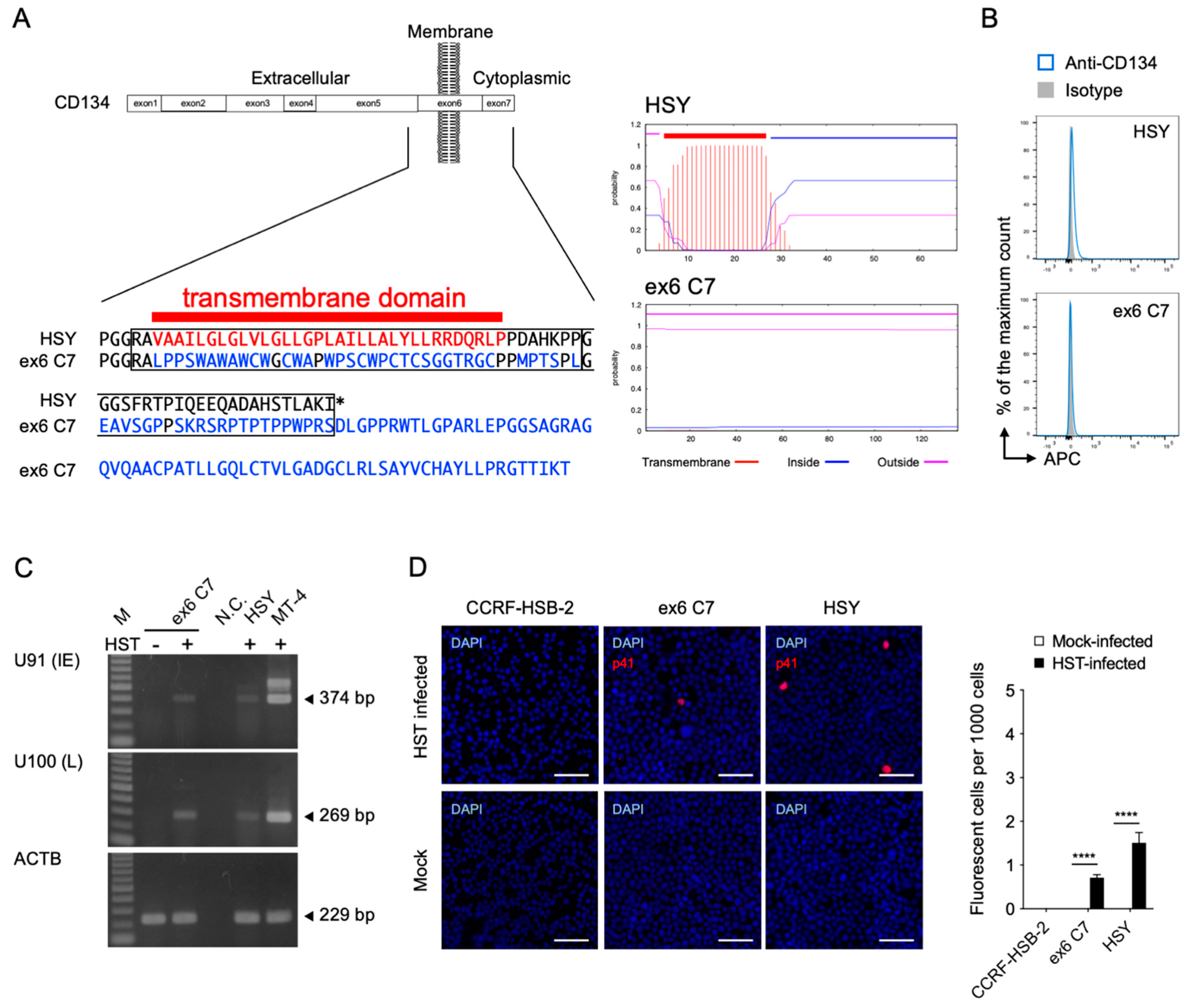
3.1. HHV-6B Infects the HSY Parotid Gland Cell Line
3.2. HHV-6B Infects Ex6 C7 Cells (CD134-Membrane Unanchored HSY Cells)
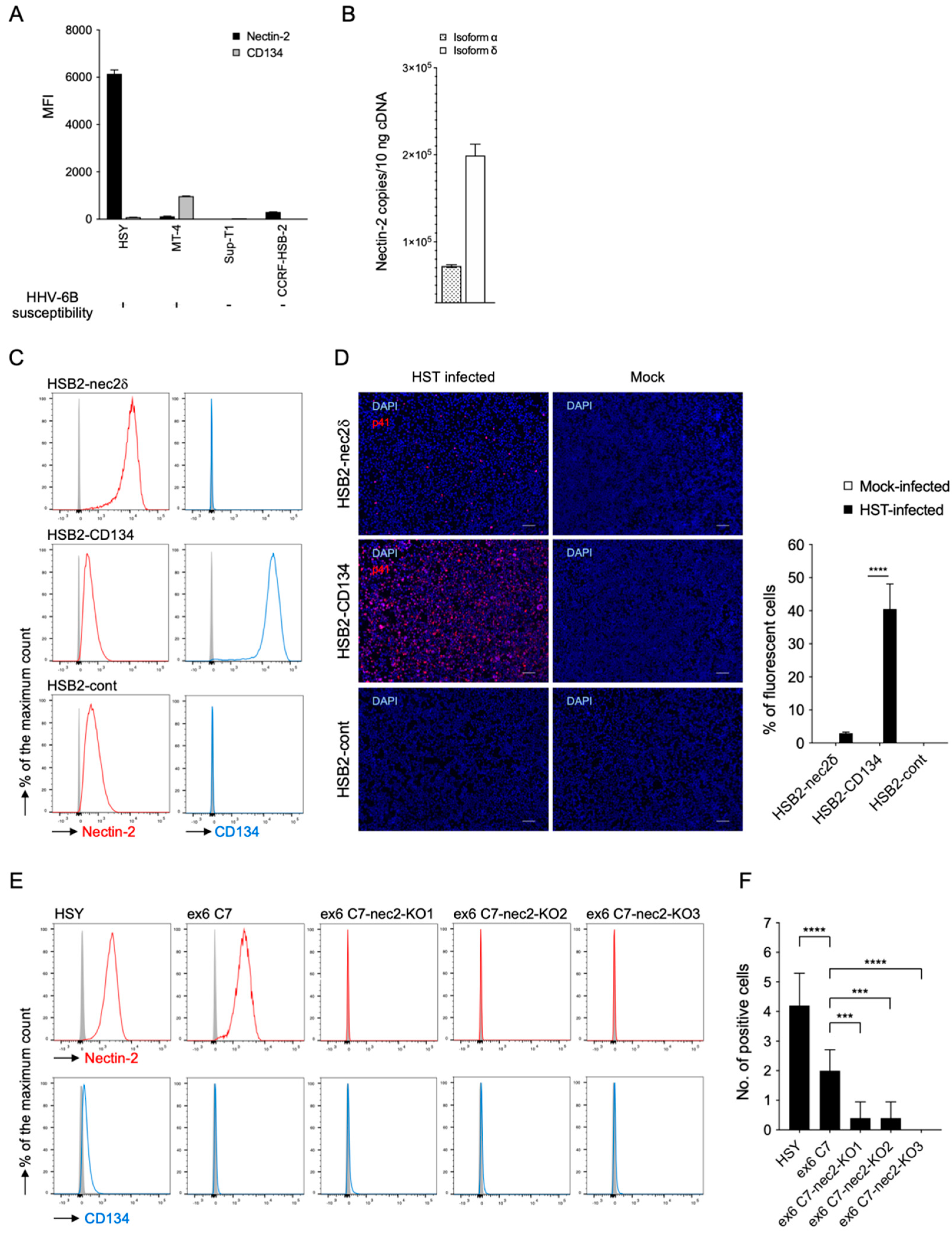
3.3. Nectin-2 Is Required for HHV-6B Infection
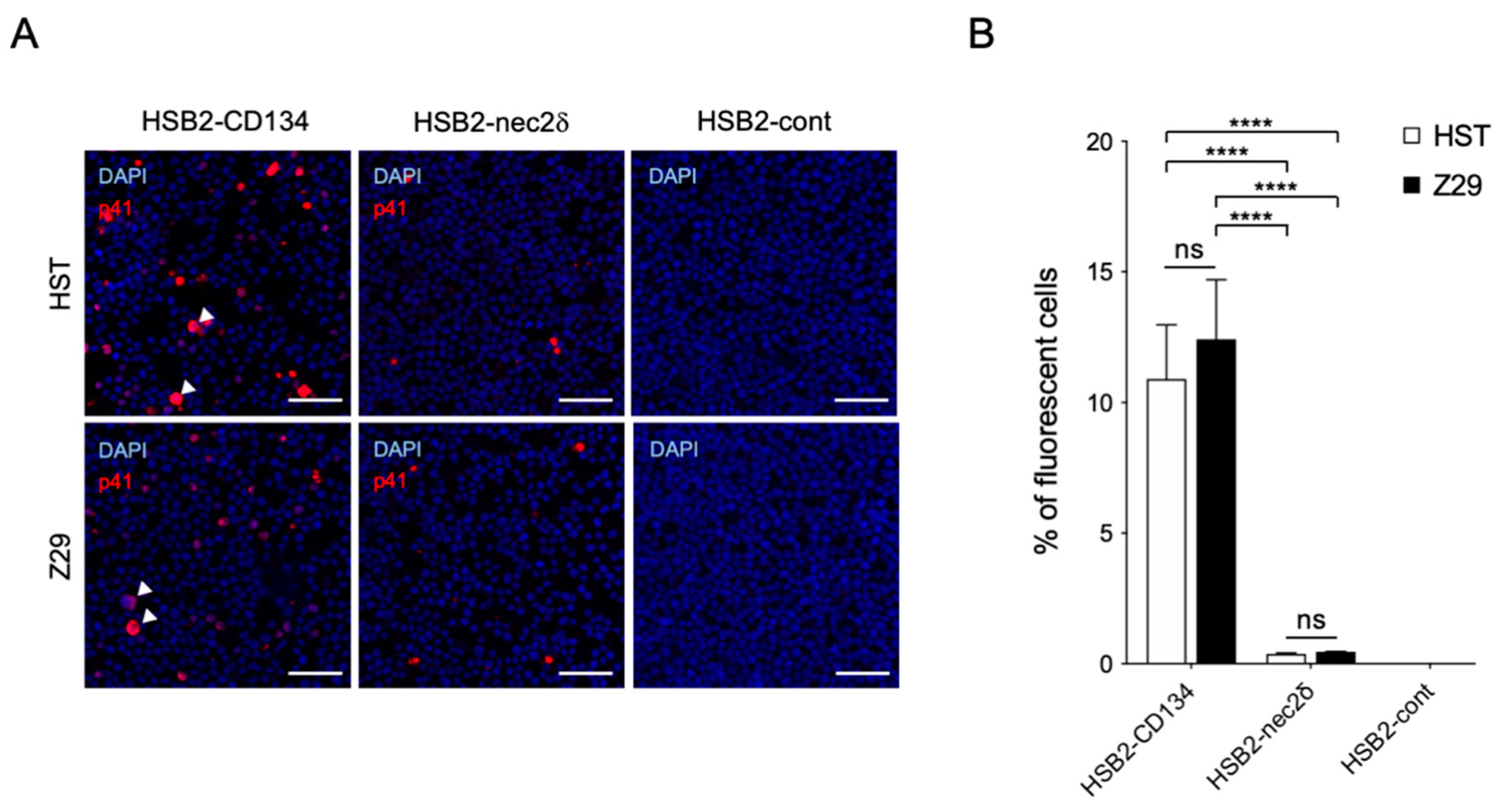
3.4. Nectin-2 Is a Functional Entry-Mediated Molecule for HHV-6B HST and Z29 Strains
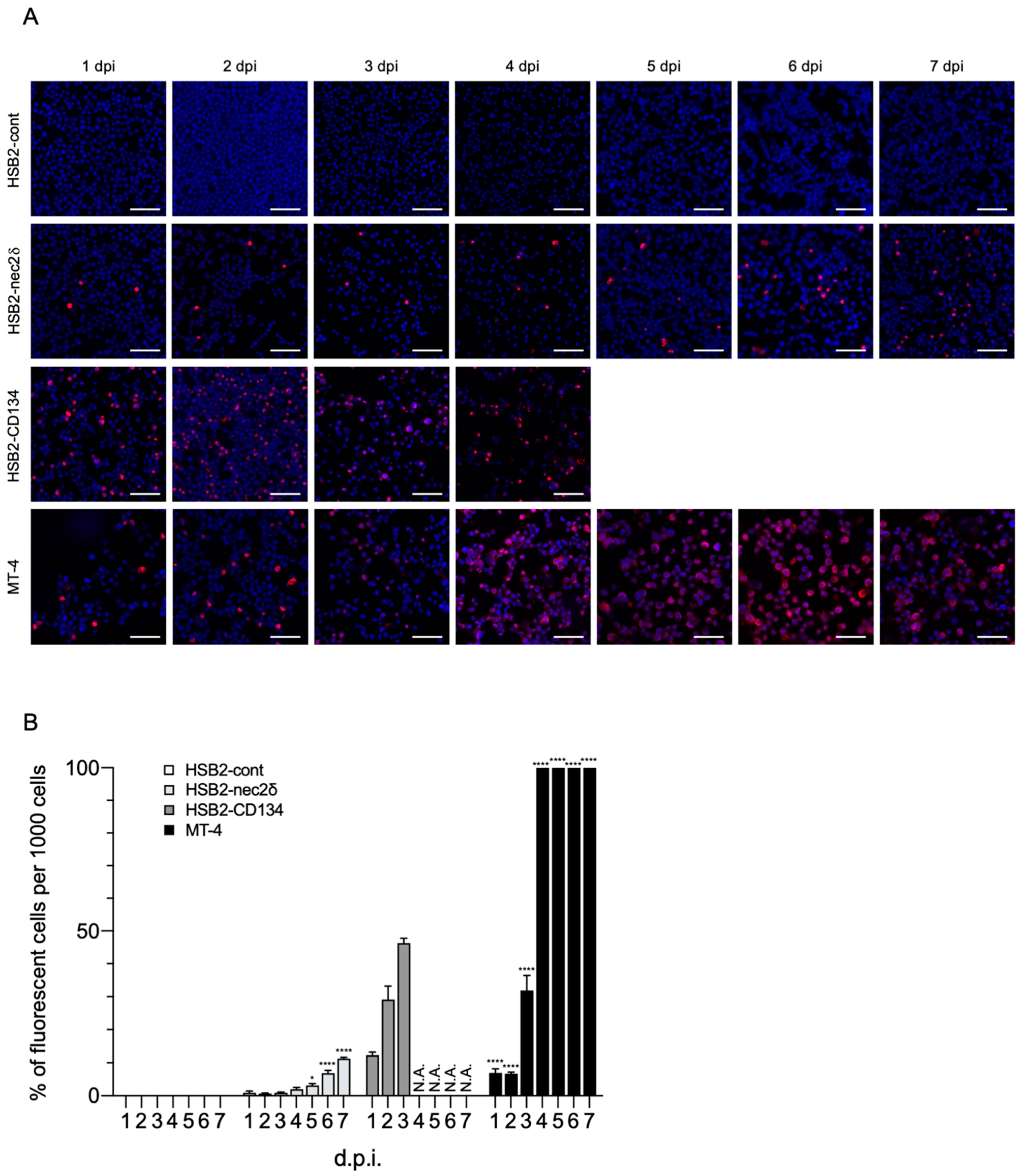
3.5. Nectin-2 Enables HHV-6B Replication in CCRF-HSB-2 Cells
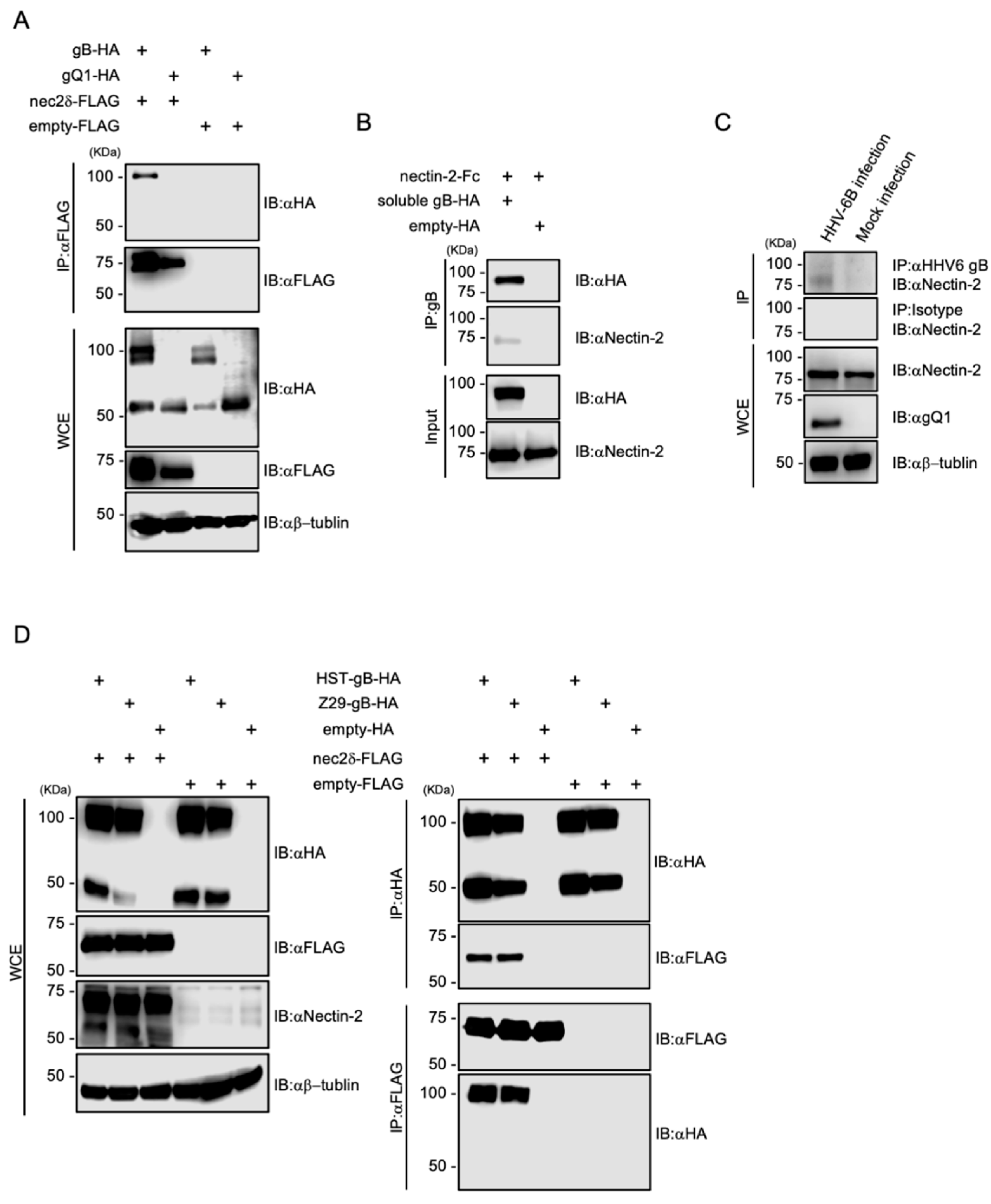
3.6. Nectin-2 Interacts with HHV-6B gB
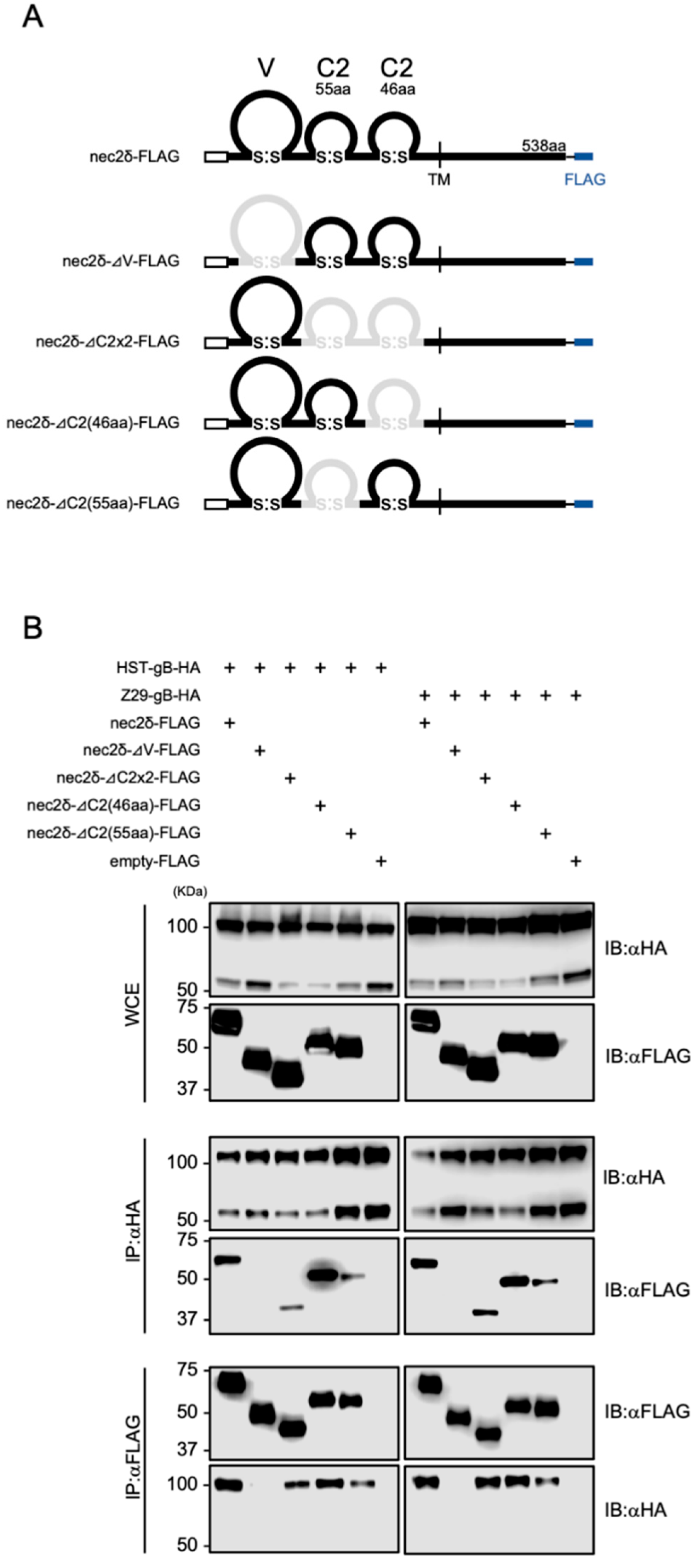
3.7. The V-set Domain of Nectin-2 Is Important for Interactions with HHV-6B gB
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ablashi, D.; Agut, H.; Alvarez-Lafuente, R.; Clark, D.A.; Dewhurst, S.; DiLuca, D.; Flamand, L.; Frenkel, N.; Gallo, R.; Gompels, U.A.; et al. Classification of HHV-6A and HHV-6B as distinct viruses. Arch. Virol. 2014, 159, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, Y.; Namba, H.; Torigoe, S.; Watanabe, M.; Yamashita, N.; Ogawa, H.; Morishima, T.; Yamada, M. Monitoring of human herpesviruses-6 and -7 DNA in saliva samples during the acute and convalescent phases of exanthem subitum. J. Med. Virol. 2017, 89, 696–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salahuddin, S.Z.; Ablashi, D.V.; Markham, P.D.; Josephs, S.F.; Sturzenegger, S.; Kaplan, M.; Halligan, G.; Biberfeld, P.; Wong-Staal, F.; Kramarsky, B.; et al. Isolation of a new virus, HBLV, in patients with lymphoproliferative disorders. Science 1986, 234, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Yamanishi, K.; Okuno, T.; Shiraki, K.; Takahashi, M.; Kondo, T.; Asano, Y.; Kurata, T. Identification of human herpesvirus-6 as a causal agent for exanthem subitum. Lancet 1988, 1, 1065–1067. [Google Scholar] [CrossRef]
- Frenkel, N.; Schirmer, E.C.; Wyatt, L.S.; Katsafanas, G.; Roffman, E.; Danovich, R.M.; June, C.H. Isolation of a new herpesvirus from human CD4+ T cells. Proc. Natl. Acad. Sci. USA 1990, 87, 748–752. [Google Scholar] [CrossRef] [Green Version]
- Caserta, M.T. Roseola (Human herpes viruses 6 and 7). In Nelson Textbook of Pediatrics, 19th ed.; Kliegman, R.M., Stanton, B.F., St. Geme III, J.W., Schor, N.F., Behrman, R.E., Eds.; Elsevier Saunders: Philadelphia, PA, USA, 2011; pp. 1117–1120. [Google Scholar]
- Tang, H.; Serada, S.; Kawabata, A.; Ota, M.; Hayashi, E.; Naka, T.; Yamanishi, K.; Mori, Y. CD134 is a cellular receptor specific for human herpesvirus-6B entry. Proc. Natl. Acad. Sci. USA 2013, 110, 9096–9099. [Google Scholar] [CrossRef] [Green Version]
- Croft, M.; So, T.; Duan, W.; Soroosh, P. The significance of OX40 and OX40L to T-cell biology and immune disease. Immunol. Rev. 2009, 229, 173–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lusso, P.; Markham, P.D.; Tschachler, E.; di Marzo Veronese, F.; Salahuddin, S.Z.; Ablashi, D.V.; Pahwa, S.; Krohn, K.; Gallo, R.C. In vitro cellular tropism of human B-lymphotropic virus (human herpesvirus-6). J. Exp. Med. 1988, 167, 1659–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Luca, D.; Mirandola, P.; Ravaioli, T.; Bigoni, B.; Cassai, E. Distribution of HHV-6 variants in human tissues. Infect. Agents Dis. 1996, 5, 203–214. [Google Scholar]
- Collot, S.; Petit, B.; Bordessoule, D.; Alain, S.; Touati, M.; Denis, F.; Ranger-Rogez, S. Real-time PCR for quantification of human herpesvirus 6 DNA from lymph nodes and saliva. J. Clin. Microbiol. 2002, 40, 2445–2451. [Google Scholar] [CrossRef] [Green Version]
- Asano, Y.; Yoshikawa, T.; Suga, S.; Yazaki, T.; Kondo, K.; Yamanishi, K. Fatal fulminant hepatitis in an infant with human herpesvirus-6 infection. Lancet 1990, 335, 862–863. [Google Scholar] [CrossRef]
- Ozaki, Y.; Tajiri, H.; Tanaka-Taya, K.; Mushiake, S.; Kimoto, A.; Yamanishi, K.; Okada, S. Frequent detection of the human herpesvirus 6-specific genomes in the livers of children with various liver diseases. J. Clin. Microbiol. 2001, 39, 2173–2177. [Google Scholar] [CrossRef] [Green Version]
- Chan, P.K.; Ng, H.K.; Hui, M.; Cheng, A.F. Prevalence and distribution of human herpesvirus 6 variants A and B in adult human brain. J. Med. Virol. 2001, 64, 42–46. [Google Scholar] [CrossRef]
- Ishiguro, N.; Yamada, S.; Takahashi, T.; Takahashi, Y.; Togashi, T.; Okuno, T.; Yamanishi, K. Meningo-encephalitis associated with HHV-6 related exanthem subitum. Acta. Paediatr. Scand. 1990, 79, 987–989. [Google Scholar] [CrossRef] [PubMed]
- Thul, P.J.; Akesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Bjork, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, 820. [Google Scholar] [CrossRef]
- Arii, J.; Hirohata, Y.; Kato, A.; Kawaguchi, Y. Nonmuscle myosin heavy chain IIb mediates herpes simplex virus 1 entry. J. Virol. 2015, 89, 1879–1888. [Google Scholar] [CrossRef] [Green Version]
- Spear, P.G.; Longnecker, R. Herpesvirus entry: An update. J. Virol. 2003, 77, 10179–10185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraghty, R.J.; Krummenacher, C.; Cohen, G.H.; Eisenberg, R.J.; Spear, P.G. Entry of alphaherpesviruses mediated by poliovirus receptor-related protein 1 and poliovirus receptor. Science 1998, 280, 1618–1620. [Google Scholar] [CrossRef]
- Warner, M.S.; Geraghty, R.J.; Martinez, W.M.; Montgomery, R.I.; Whitbeck, J.C.; Xu, R.; Eisenberg, R.J.; Cohen, G.H.; Spear, P.G. A cell surface protein with herpesvirus entry activity (HveB) confers susceptibility to infection by mutants of herpes simplex virus type 1, herpes simplex virus type 2, and pseudorabies virus. Virology 1998, 246, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, R.I.; Warner, M.S.; Lum, B.J.; Spear, P.G. Herpes simplex virus-1 entry into cells mediated by a novel member of the TNF/NGF receptor family. Cell 1996, 87, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Shukla, D.; Liu, J.; Blaiklock, P.; Shworak, N.W.; Bai, X.; Esko, J.D.; Cohen, G.H.; Eisenberg, R.J.; Rosenberg, R.D.; Spear, P.G. A novel role for 3-O-sulfated heparan sulfate in herpes simplex virus 1 entry. Cell 1999, 99, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Satoh, T.; Arii, J.; Suenaga, T.; Wang, J.; Kogure, A.; Uehori, J.; Arase, N.; Shiratori, I.; Tanaka, S.; Kawaguchi, Y.; et al. PILRalpha is a herpes simplex virus-1 entry coreceptor that associates with glycoprotein B. Cell 2008, 132, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Suenaga, T.; Satoh, T.; Somboonthum, P.; Kawaguchi, Y.; Mori, Y.; Arase, H. Myelin-associated glycoprotein mediates membrane fusion and entry of neurotropic herpesviruses. Proc. Natl. Acad. Sci. USA 2010, 107, 866–871. [Google Scholar] [CrossRef] [Green Version]
- Arii, J.; Goto, H.; Suenaga, T.; Oyama, M.; Kozuka-Hata, H.; Imai, T.; Minowa, A.; Akashi, H.; Arase, H.; Kawaoka, Y.; et al. Non-muscle myosin IIA is a functional entry receptor for herpes simplex virus-1. Nature 2010, 467, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Pellett, P.E.; Roizman, B. Herpesviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 2, pp. 1802–1822. [Google Scholar]
- Dominguez, G.; Dambaugh, T.R.; Stamey, F.R.; Dewhurst, S.; Inoue, N.; Pellett, P.E. Human herpesvirus 6B genome sequence: Coding content and comparison with human herpesvirus 6A. J. Virol. 1999, 73, 8040–8052. [Google Scholar] [CrossRef] [Green Version]
- Isegawa, Y.; Mukai, T.; Nakano, K.; Kagawa, M.; Chen, J.; Mori, Y.; Sunagawa, T.; Kawanishi, K.; Sashihara, J.; Hata, A.; et al. Comparison of the complete DNA sequences of human herpesvirus 6 variants A and B. J. Virol. 1999, 73, 8053–8063. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, A.; Jasirwan, C.; Yamanishi, K.; Mori, Y. Human herpesvirus 6 glycoprotein M is essential for virus growth and requires glycoprotein N for its maturation. Virology 2012, 429, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Gompels, U.A.; Nicholas, J.; Lawrence, G.; Jones, M.; Thomson, B.J.; Martin, M.E.; Efstathiou, S.; Craxton, M.; Macaulay, H.A. The DNA sequence of human herpesvirus-6: Structure, coding content, and genome evolution. Virology 1995, 209, 29–51. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Yang, X.; Akkapaiboon, P.; Okuno, T.; Yamanishi, K. Human herpesvirus 6 variant A glycoprotein H-glycoprotein L-glycoprotein Q complex associates with human CD46. J. Virol. 2003, 77, 4992–4999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, Y.; Akkapaiboon, P.; Yonemoto, S.; Koike, M.; Takemoto, M.; Sadaoka, T.; Sasamoto, Y.; Konishi, S.; Uchiyama, Y.; Yamanishi, K. Discovery of a second form of tripartite complex containing gH-gL of human herpesvirus 6 and observations on CD46. J. Virol. 2004, 78, 4609–4616. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.D.; Briggs, M.; Ward, P.A.; Tedder, R.S. Human herpesvirus 6 in salivary glands. Lancet 1990, 336, 590–593. [Google Scholar] [CrossRef]
- De Bolle, L.; Naesens, L.; De Clercq, E. Update on human herpesvirus 6 biology, clinical features, and therapy. Clin. Microbiol. Rev. 2005, 18, 217–245. [Google Scholar] [CrossRef] [Green Version]
- Yanagawa, T.; Hayashi, Y.; Nagamine, S.; Yoshida, H.; Yura, Y.; Sato, M. Generation of cells with phenotypes of both intercalated duct-type and myoepithelial cells in human parotid gland adenocarcinoma clonal cells grown in athymic nude mice. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1986, 51, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Eberlé, F.; Dubreuil, P.; Mattei, M.G.; Devilard, E.; Lopez, M. The human PRR2 gene, related to the human poliovirus receptor gene (PVR), is the true homolog of the murine MPH gene. Gene 1995, 159, 267–272. [Google Scholar] [CrossRef]
- Tsukazaki, T.; Yoshida, M.; Namba, H.; Yamada, M.; Shimizu, N.; Nii, S. Development of a dot blot neutralizing assay for HHV-6 and HHV-7 using specific monoclonal antibodies. J. Virol. Methods 1998, 73, 141–149. [Google Scholar] [CrossRef]
- Mirandola, P.; Menegazzi, P.; Merighi, S.; Ravaioli, T.; Cassai, E.; Di Luca, D. Temporal mapping of transcripts in herpesvirus 6 variants. J. Virol. 1998, 72, 3837–3844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujikura, D.; Ikesue, M.; Endo, T.; Chiba, S.; Higashi, H.; Uede, T. Death receptor 6 contributes to autoimmunity in lupus-prone mice. Nat. Commun. 2017, 8, 13957. [Google Scholar] [CrossRef] [Green Version]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [Green Version]
- Möller, S.; Croning, M.D.; Apweiler, R. Evaluation of methods for the prediction of membrane spanning regions. Bioinformatics 2001, 17, 646–653. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, M.; Fujikura, D.; Nanbo, A.; Marzi, A.; Noyori, O.; Kajihara, M.; Maruyama, J.; Matsuno, K.; Miyamoto, H.; Yoshida, R.; et al. Interaction between TIM-1 and NPC1 Is Important for Cellular Entry of Ebola Virus. J. Virol. 2015, 89, 6481–6493. [Google Scholar] [CrossRef] [Green Version]
- Niwa, H.; Yamamura, K.; Miyazaki, J. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 1991, 108, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, S.; Niki, H. Exonuclease III (XthA) Enforces In Vivo DNA Cloning of Escherichia coli To Create Cohesive Ends. J. Bacteriol. 2019, 201, e00660-18. [Google Scholar] [CrossRef] [Green Version]
- Takai, Y.; Ikeda, W.; Ogita, H.; Rikitake, Y. The immunoglobulin-like cell adhesion molecule nectin and its associated protein afadin. Annu. Rev. Cell Dev. Biol. 2008, 24, 309–342. [Google Scholar] [CrossRef]
- Lopez, M.; Cocchi, F.; Menotti, L.; Avitabile, E.; Dubreuil, P.; Campadelli-Fiume, G. Nectin2alpha (PRR2alpha or HveB) and nectin2delta are low-efficiency mediators for entry of herpes simplex virus mutants carrying the Leu25Pro substitution in glycoprotein D. J. Virol. 2000, 74, 1267–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, W.M.; Spear, P.G. Structural features of nectin-2 (HveB) required for herpes simplex virus entry. J. Virol. 2001, 75, 11185–11195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikyo, M.; Matozaki, T.; Kodama, A.; Kawabe, H.; Nakanishi, H.; Takai, Y. Cell-cell adhesion-mediated tyrosine phosphorylation of nectin-2delta, an immunoglobulin-like cell adhesion molecule at adherens junctions. Oncogene 2000, 19, 4022–4028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, I.; Taguchi, H.; Kubonishi, I.; Yoshimoto, S.; Ohtsuki, Y.; Shiraishi, Y.; Akagi, T. Type C virus-producing cell lines derived from adult T cell leukemia. Gann. Monogr. Cancer Res. 1982, 28, 219–228. [Google Scholar]
- Mühleisen, A.; Giaisi, M.; Köhler, R.; Krammer, P.H.; Li-Weber, M. Tax contributes apoptosis resistance to HTLV-1-infected T cells via suppression of Bid and Bim expression. Cell Death Dis. 2014, 5, e1575. [Google Scholar] [CrossRef] [Green Version]
- Heldwein, E.E.; Lou, H.; Bender, F.C.; Cohen, G.H.; Eisenberg, R.J.; Harrison, S.C. Crystal structure of glycoprotein B from herpes simplex virus 1. Science 2006, 313, 217–220. [Google Scholar] [CrossRef] [Green Version]
- Foà-Tomasi, L.; Guerrini, S.; Huang, T.; Campadelli-Fiume, G. Characterization of human herpesvirus-6(U1102) and (GS) gp112 and identification of the Z29-specified homolog. Virology 1992, 191, 511–516. [Google Scholar] [CrossRef]
- Santoro, F.; Kennedy, P.E.; Locatelli, G.; Malnati, M.S.; Berger, E.A.; Lusso, P. CD46 is a cellular receptor for human herpesvirus 6. Cell 1999, 99, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Lusso, P.; Secchiero, P.; Crowley, R.W.; Garzino-Demo, A.; Berneman, Z.N.; Gallo, R.C. CD4 is a critical component of the receptor for human herpesvirus 7: Interference with human immunodeficiency virus. Proc. Natl. Acad. Sci. USA 1994, 91, 3872–3876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mach, M.; Osinski, K.; Kropff, B.; Schloetzer-Schrehardt, U.; Krzyzaniak, M.; Britt, W. The carboxy-terminal domain of glycoprotein N of human cytomegalovirus is required for virion morphogenesis. J. Virol. 2007, 81, 5212–5224. [Google Scholar] [CrossRef] [Green Version]
- Oliver, S.L.; Sommer, M.; Zerboni, L.; Rajamani, J.; Grose, C.; Arvin, A.M. Mutagenesis of varicella-zoster virus glycoprotein B: Putative fusion loop residues are essential for viral replication, and the furin cleavage motif contributes to pathogenesis in skin tissue in vivo. J. Virol. 2009, 83, 7495–7506. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Suenaga, T.; Matsumoto, M.; Seya, T.; Arase, H. Herpesvirus 6 glycoproteins B (gB), gH, gL, and gQ are necessary and sufficient for cell-to-cell fusion. J. Virol. 2013, 87, 10900–10903. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Seya, T.; Huang, H.L.; Akkapaiboon, P.; Dhepakson, P.; Yamanishi, K. Human herpesvirus 6 variant A but not variant B induces fusion from without in a variety of human cells through a human herpesvirus 6 entry receptor, CD46. J. Virol. 2002, 76, 6750–6761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawabata, A.; Oyaizu, H.; Maeki, T.; Tang, H.; Yamanishi, K.; Mori, Y. Analysis of a neutralizing antibody for human herpesvirus 6B reveals a role for glycoprotein Q1 in viral entry. J. Virol. 2011, 85, 12962–12971. [Google Scholar] [CrossRef] [Green Version]
- Honda, T.; Shimizu, K.; Kawakatsu, T.; Yasumi, M.; Shingai, T.; Fukuhara, A.; Ozaki-Kuroda, K.; Irie, K.; Nakanishi, H.; Takai, Y. Antagonistic and agonistic effects of an extracellular fragment of nectin on formation of E-cadherin-based cell-cell adhesion. Genes Cells 2003, 8, 51–63. [Google Scholar] [CrossRef]
- Tatsuo, H.; Ono, N.; Tanaka, K.; Yanagi, Y. SLAM (CDw150) is a cellular receptor for measles virus. Nature 2000, 406, 893–897. [Google Scholar] [CrossRef]
- Mühlebach, M.D.; Mateo, M.; Sinn, P.L.; Prüfer, S.; Uhlig, K.M.; Leonard, V.H.; Navaratnarajah, C.K.; Frenzke, M.; Wong, X.X.; Sawatsky, B.; et al. Adherens junction protein nectin-4 is the epithelial receptor for measles virus. Nature 2011, 480, 530–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalgleish, A.G.; Beverley, P.C.; Clapham, P.R.; Crawford, D.H.; Greaves, M.F.; Weiss, R.A. The CD4 (T4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature 1984, 312, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, C.L.; Wimmer, E.; Racaniello, V.R. Cellular receptor for poliovirus: Molecular cloning, nucleotide sequence, and expression of a new member of the immunoglobulin superfamily. Cell 1989, 56, 855–865. [Google Scholar] [CrossRef]
- Cocchi, F.; Lopez, M.; Menotti, L.; Aoubala, M.; Dubreuil, P.; Campadelli-Fiume, G. The V domain of herpesvirus Ig-like receptor (HIgR) contains a major functional region in herpes simplex virus-1 entry into cells and interacts physically with the viral glycoprotein D. Proc. Natl. Acad. Sci. USA 1998, 95, 15700–15705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, N.; Tatsuo, H.; Tanaka, K.; Minagawa, H.; Yanagi, Y. V domain of human SLAM (CDw150) is essential for its function as a measles virus receptor. J. Virol. 2001, 75, 1594–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, S.; Ise, I.; Nomoto, A. Functional domains of the poliovirus receptor. Proc. Natl. Acad. Sci. USA 1991, 88, 4104–4108. [Google Scholar] [CrossRef] [Green Version]
- Arthos, J.; Deen, K.C.; Chaikin, M.A.; Fornwald, J.A.; Sathe, G.; Sattentau, Q.J.; Clapham, P.R.; Weiss, R.A.; McDougal, J.S.; Pietropaolo, C.; et al. Identification of the residues in human CD4 critical for the binding of HIV. Cell 1989, 57, 469–481. [Google Scholar] [CrossRef]
- Morrison, M.E.; He, Y.J.; Wien, M.W.; Hogle, J.M.; Racaniello, V.R. Homolog-scanning mutagenesis reveals poliovirus receptor residues important for virus binding and replication. J. Virol. 1994, 68, 2578–2588. [Google Scholar] [CrossRef] [Green Version]
- Di Giovine, P.; Settembre, E.C.; Bhargava, A.K.; Luftig, M.A.; Lou, H.; Cohen, G.H.; Eisenberg, R.J.; Krummenacher, C.; Carfi, A. Structure of herpes simplex virus glycoprotein D bound to the human receptor nectin-1. PLoS Pathog. 2011, 7, e1002277. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Yan, J.; Lu, G.; Guo, Z.; Fan, Z.; Wang, J.; Shi, Y.; Qi, J.; Gao, G.F. Binding of herpes simplex virus glycoprotein D to nectin-1 exploits host cell adhesion. Nat. Commun. 2011, 2, 577. [Google Scholar] [CrossRef]
- Liu, J.; Qian, X.; Chen, Z.; Xu, X.; Gao, F.; Zhang, S.; Zhang, R.; Qi, J.; Gao, G.F.; Yan, J. Crystal structure of cell adhesion molecule nectin-2/CD112 and its binding to immune receptor DNAM-1/CD226. J. Immunol. 2012, 188, 5511–5520. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, G.; Harber, J.; Zibert, A.; deCrombrugghe, M.; Wimmer, E. The poliovirus receptor: Identification of domains and amino acid residues critical for virus binding. Virology 1994, 203, 344–356. [Google Scholar] [CrossRef]
- De Bolle, L.; Van Loon, J.; De Clercq, E.; Naesens, L. Quantitative analysis of human herpesvirus 6 cell tropism. J. Med. Virol. 2005, 75, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Takai, Y.; Nakanishi, H. Nectin and afadin: Novel organizers of intercellular junctions. J. Cell Sci. 2003, 116 Pt 1, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Baker, O.J. Current trends in salivary gland tight junctions. Tissue Barriers 2016, 4, e1162348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, M.; Tahara, M.; Nagata, N.; Seki, F. Wild-Type Measles Virus is Intrinsically Dual-Tropic. Front. Microbiol. 2011, 2, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, M.; Mandai, K.; Maruo, T.; Sato, J.; Shiotani, H.; Kaito, A.; Itoh, Y.; Wang, S.; Fujiwara, T.; Mizoguchi, A.; et al. Localization of nectin-2delta at perivascular astrocytic endfoot processes and degeneration of astrocytes and neurons in nectin-2 knockout mouse brain. Brain Res. 2016, 1649 Pt A, 90–101. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogawa, H.; Fujikura, D.; Namba, H.; Yamashita, N.; Honda, T.; Yamada, M. Nectin-2 Acts as a Viral Entry Mediated Molecule That Binds to Human Herpesvirus 6B Glycoprotein B. Viruses 2022, 14, 160. https://doi.org/10.3390/v14010160
Ogawa H, Fujikura D, Namba H, Yamashita N, Honda T, Yamada M. Nectin-2 Acts as a Viral Entry Mediated Molecule That Binds to Human Herpesvirus 6B Glycoprotein B. Viruses. 2022; 14(1):160. https://doi.org/10.3390/v14010160
Chicago/Turabian StyleOgawa, Hirohito, Daisuke Fujikura, Hikaru Namba, Nobuko Yamashita, Tomoyuki Honda, and Masao Yamada. 2022. "Nectin-2 Acts as a Viral Entry Mediated Molecule That Binds to Human Herpesvirus 6B Glycoprotein B" Viruses 14, no. 1: 160. https://doi.org/10.3390/v14010160
APA StyleOgawa, H., Fujikura, D., Namba, H., Yamashita, N., Honda, T., & Yamada, M. (2022). Nectin-2 Acts as a Viral Entry Mediated Molecule That Binds to Human Herpesvirus 6B Glycoprotein B. Viruses, 14(1), 160. https://doi.org/10.3390/v14010160