
Novel Epidemic Metrics to Communicate Outbreak Risk at the Municipality Level: Dengue and Zika in the Dominican Republic



Abstract
:1. Introduction
2. Study Aims
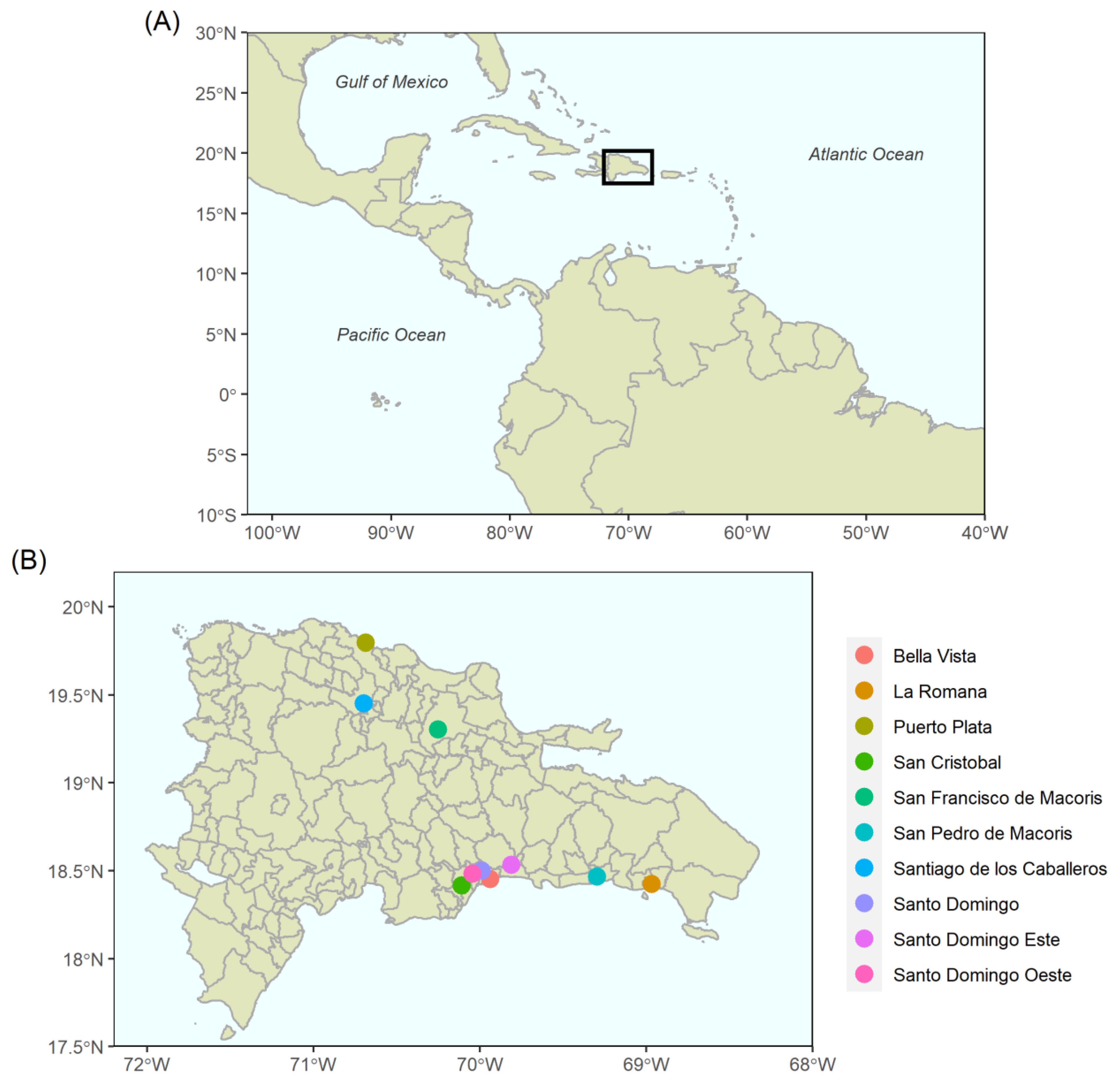
3. Country Context: Dominican Republic
4. Methods
4.1. Datasets
4.2. Mapping
4.3. Statistical Analyses
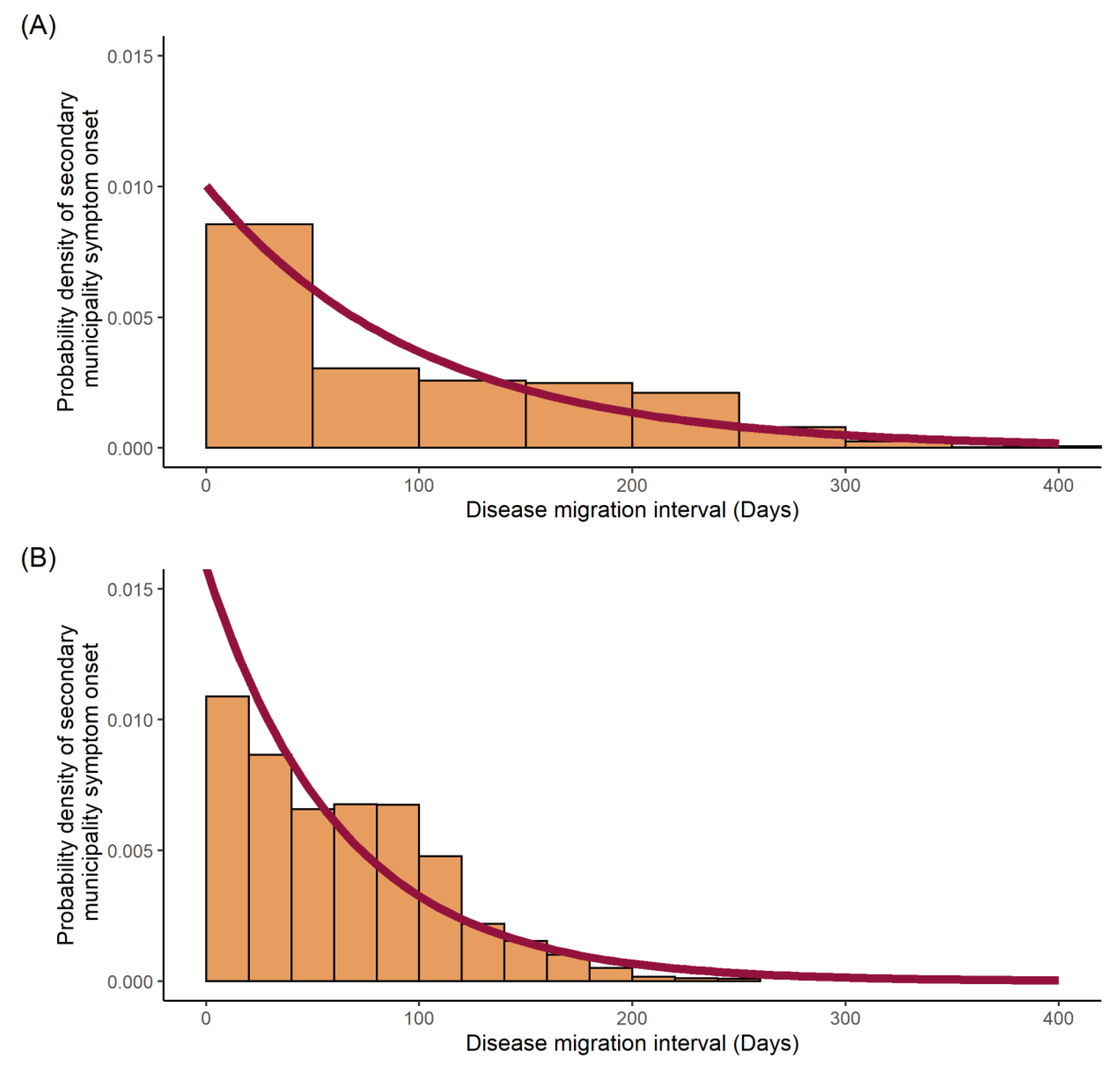
4.4. Disease Migration Interval Distribution
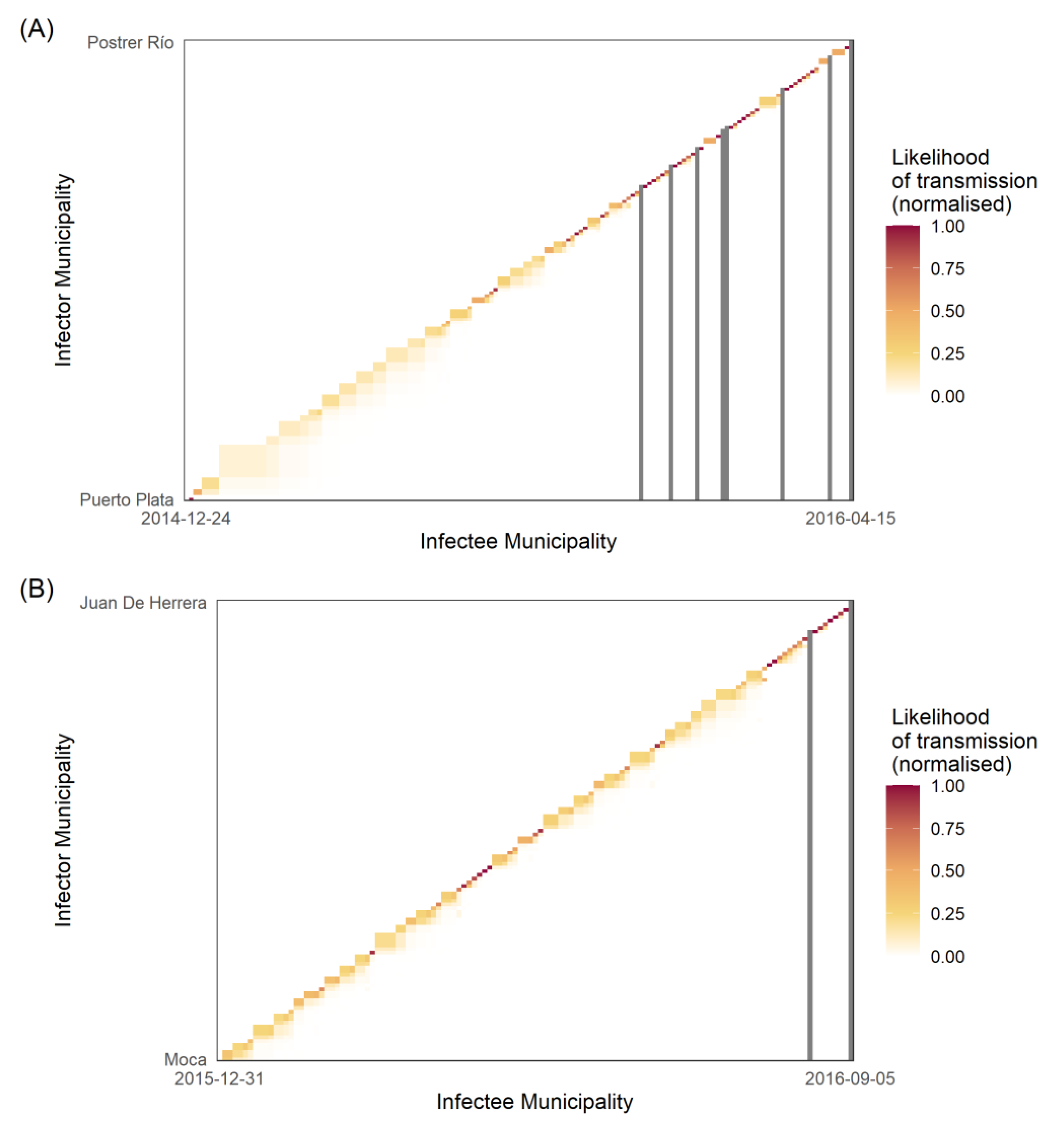
4.5. Determining the Transmission Likelihood and Network
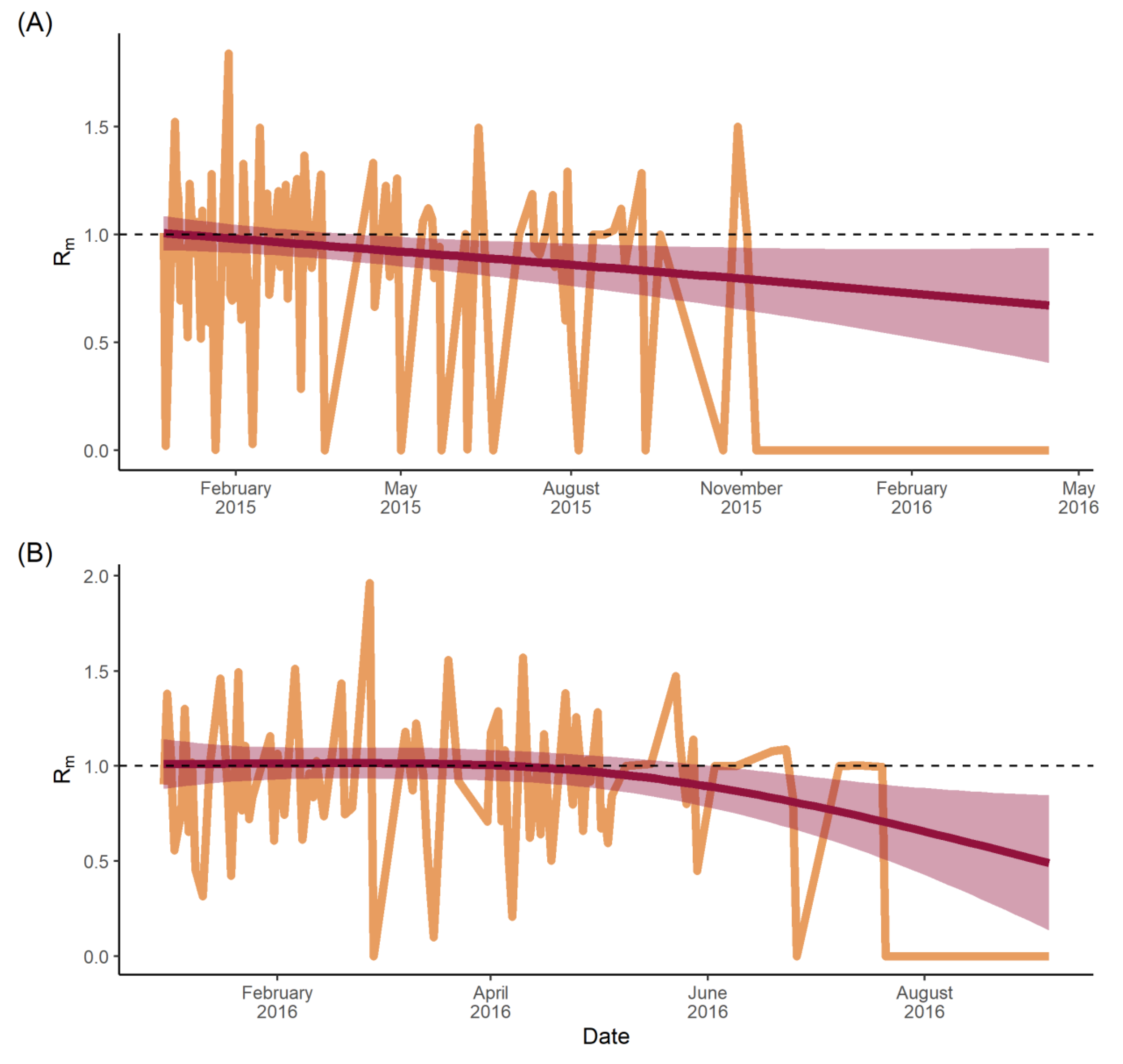
4.6. Estimating Time-Varying Municipal-Specific Reproduction Numbers
4.7. Ethics
5. Results
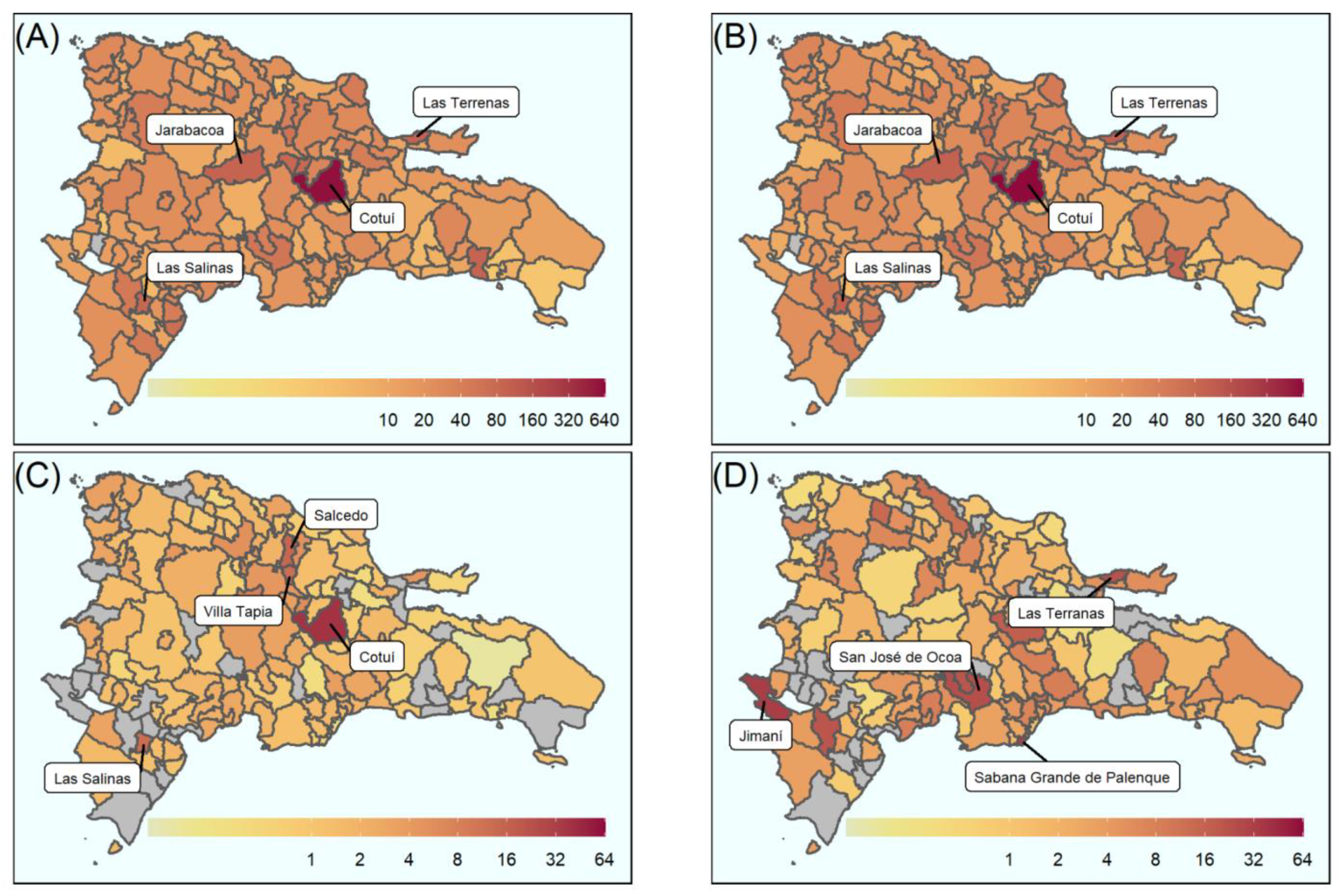
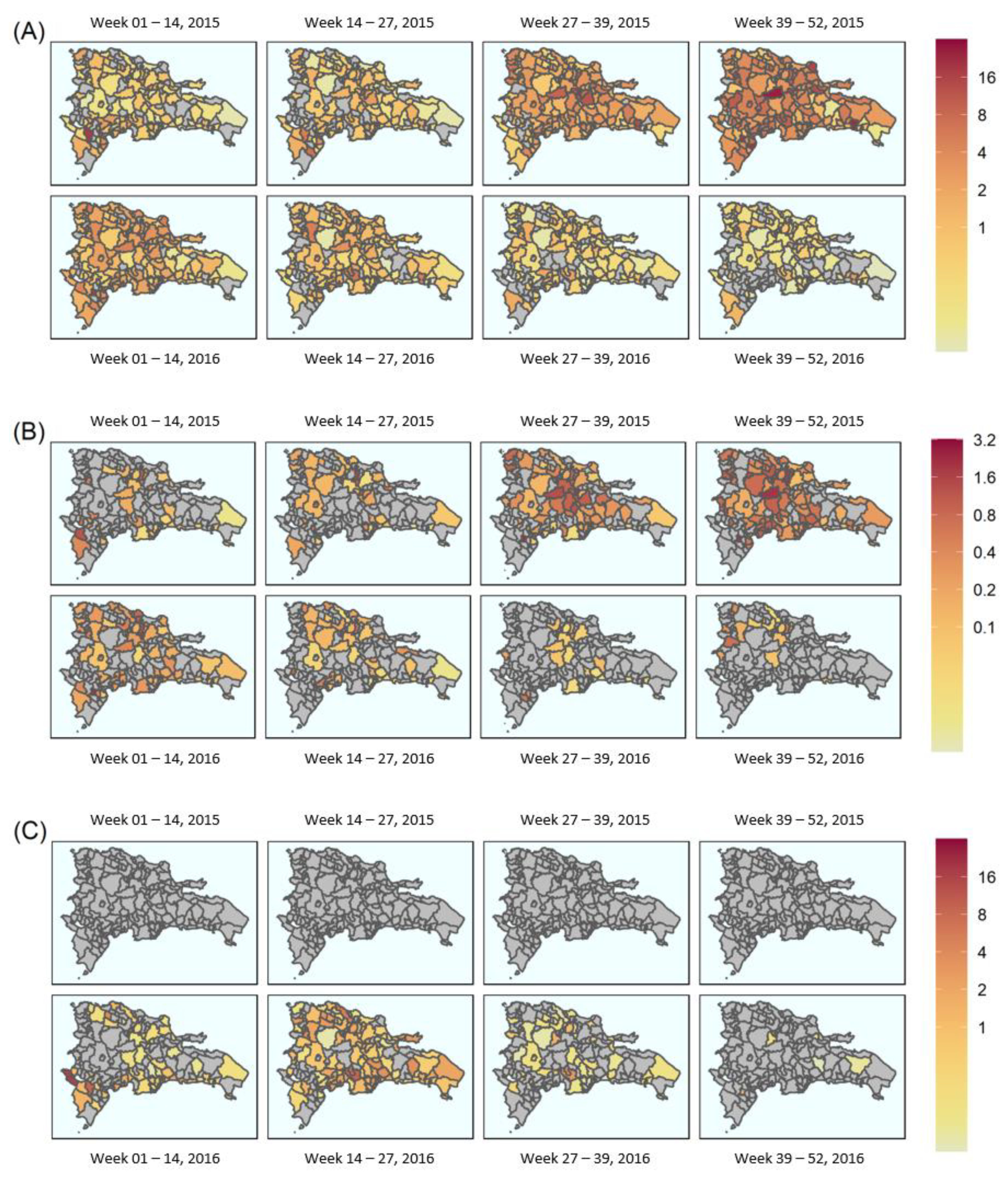
5.1. Geostatistical Mapping
5.2. Transmission Dynamics
6. Discussion
6.1. Human Mobility and Infrastructure
6.2. Transmission Chains
6.3. Municipal Reproduction Number (Rm)
6.4. Limitations
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, L.E.; Beckham, J.D.; Tyler, K.L. North American Encephalitic Arboviruses. Neurol. Clin. 2008, 26, 727–757. [Google Scholar] [CrossRef] [Green Version]
- Beckham, J.D.; Tyler, K.L. Arbovirus Infections. Contin. Lifelong Learn. Neurol. 2015, 21, 1599–1621.1. [Google Scholar] [CrossRef] [Green Version]
- Bowman, L.R.; Donegan, S.; McCall, P.J. Is Dengue Vector Control Deficient in Effectiveness or Evidence?: Systematic Review and Meta-analysis. PLoS Negl. Trop. Dis. 2016, 10, e0004551. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2010, 496, 504–507. [Google Scholar] [CrossRef]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the Global Spatial Limits of Dengue Virus Transmission by Evidence-Based Consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef] [PubMed]
- Pan American Health Organization; World Health Organization. PLISA Health Information Platform for the Americas: Cases of Zika Virus Disease by Country or Territory; WHO: Washington, DC, USA, 2019. [Google Scholar]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue Barriers to Arbovirus Infection in Mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [PubMed]
- Chye, J.K.; Lim, C.T.; Ng, K.B.; Lim, J.M.H.; George, R.; Lam, S.K. Vertical Transmission of Dengue. Clin. Infect. Dis. 1997, 25, 1374–1377. [Google Scholar] [CrossRef] [Green Version]
- Pomar, L.; Vouga, M.; Lambert, V.; Pomar, C.; Hcini, N.; Jolivet, A.; Benoist, G.; Rousset, D.; Matheus, S. Maternal-fetal transmission and adverse perinatal outcomes in pregnant women infected with Zika virus: Prospective cohort study in French Guiana. Obstet. Anesth. Dig. 2019, 39, 89–90. [Google Scholar] [CrossRef]
- Counotte, M.J.; Kim, C.R.; Wang, J.; Bernstein, K.; Deal, C.D.; Broutet, N.J.N.; Low, N. Sexual transmission of Zika virus and other flaviviruses: A living systematic review. PLoS Med. 2018, 15, e1002611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothman, K.; Greenland, S. Modern Epidemiology, 2nd ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1998. [Google Scholar]
- Barratt, H. Epidemic theory (effective & basic reproduction numbers, epidemic thresholds) & techniques for analysis of infectious disease data (construction & use of epidemic curves, generation numbers, exceptional reporting & identification of significant clusters). Health Knowl. 2019. Available online: https://www.healthknowledge.org.uk/public-health-textbook/research-methods/1a-epidemiology/epidemic-theory (accessed on 2 April 2020).
- Diekmann, O.; Heesterbeek, J.A.P.; Metz, J.A.J. On the definition and the computation of the basic reproduction ratio R 0 in models for infectious diseases in heterogeneous populations. J. Math. Biol. 1990, 28, 365–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kermack, W.; McKendrick, A. A contribution to the mathematical theory of epidemics. Proc. R. Soc. Lond. Ser. A Contain. Pap. A Math. Phys. Character 1927, 115, 700–721. [Google Scholar]
- Wallinga, J. Different Epidemic Curves for Severe Acute Respiratory Syndrome Reveal Similar Impacts of Control Measures. Am. J. Epidemiol. 2004, 160, 509–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Last, J.M. (Ed.) A Dictionary of Epidemiology, 4th ed.; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Routledge, I.; Chevéz, J.E.R.; Cucunubá, Z.M.; Rodriguez, M.G.; Guinovart, C.; Gustafson, K.B.; Schneider, K.; Walker, P.G.T.; Ghani, A.C.; Bhatt, S. Estimating spatiotemporally varying malaria reproduction numbers in a near elimination setting. Nat. Commun. 2018, 9, 2476. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C. Estimating Individual and Household Reproduction Numbers in an Emerging Epidemic. PLoS ONE 2007, 2, e758. [Google Scholar] [CrossRef] [PubMed]
- Frost, W.H. Some conceptions of epidemics in general by Wade Hampton Frost. Am. J. Epidemiol. 1976, 103, 141–151. [Google Scholar] [CrossRef]
- Abbey, H. An examination of the Reed-Frost theory of epidemics. Hum. Biol. 1952, 24, 201–233. [Google Scholar]
- Bowman, L.R.; Rocklöv, J.; Kroeger, A.; Olliaro, P.; Skewes, R. A comparison of Zika and dengue outbreaks using national surveillance data in the Dominican Republic. PLoS Negl. Trop. Dis. 2018, 12, e0006876. [Google Scholar] [CrossRef]
- Wikipedia Contributors. Dominican Republic; Wikipedia—Wikimedia Foundation: San Francisco, CA, USA, 2019. [Google Scholar]
- World Bank. Dominican Republic: Country Profile. Available online: https://databank.worldbank.org (accessed on 2 April 2020).
- Butler, R.A. Largest Cities in Dominican Republic: Mongabay. Mongabay, Dominican Republic. 2001. Available online: https://population.mongabay.com/population/dominican-republic/ (accessed on 2 April 2020).
- Dominican Republic Administrative Boundaries (Levels 0–5)—Humanitarian Data Exchange. 2014. Available online: https://data.humdata.org/dataset/dominican-republic-administrative-boundaries-levels-0-6 (accessed on 2 April 2020).
- Wickham. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016.
- Yamashiro, T.; Vardez, S.; Nishizono, A.; Petit, A.; Taveras, D.; Cesin, A.J.; Garcia, B.; Castro-Bello, M.; Lora-Orste, M.; Disla, M. seroprevalence of igg specific for dengue virus among adults and children in santo domingo, Dominican Republic. Am. J. Trop. Med. Hyg. 2004, 71, 138–143. [Google Scholar] [CrossRef] [Green Version]
- WHO. Dengue Guidelines for Diagnosis, Treatment, Prevention and Control; Ciceri, K., Tissot, P., Eds.; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- World Health Organization. Zika Virus Disease; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Bowman, L.R.; Tejeda, G.S.; Coelho, G.E.; Sulaiman, L.H.; Gill, B.S.; McCall, P.J.; Olliaro, P.L.; Ranzinger, S.R.; Quang, L.C.; Ramm, R.S.; et al. Alarm Variables for Dengue Outbreaks: A Multi-Centre Study in Asia and Latin America. PLoS ONE 2016, 11, e0157971. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2018; Available online: http://www.rstudio.com/ (accessed on 2 April 2020).
- RPubs—Age Adjusted Rates. Rpubs.com. 2017. Available online: https://rpubs.com/bpoulin-CUNY/321735 (accessed on 28 April 2020).
- Lessler, J.; Salje, H.; Giles, J. IDSpatialStats: Estimate Global Clustering in Infectious Disease. R package version 0.3.5.. 2018. Available online: https://CRAN.R-project.org/package=IDSpatialStats (accessed on 28 April 2020).
- Kempe, D.; Kleinberg, J.; Tardos, E. Maximizing the Spread of Influence through a Social Network. In Proceedings of the Ninth ACM SIGKDD International Conference on Knowledge Discovery and Data Mining—KDD ’03, Washington, DC, USA, 24–27 August 2003; pp. 137–146. [Google Scholar]
- Kilpatrick, A.M.; Randolph, S.E. Drivers, dynamics, and control of emerging vector-borne zoonotic diseases. Lancet 2012, 380, 1946–1955. [Google Scholar] [CrossRef] [Green Version]
- Energía y Minas en Jornada Contra Dengue Junto a Líderes Comunitarios Cotuí|Presidencia de la República Dominicana. 2016. Available online: https://gobiernodanilomedina.do/noticias/energia-y-minas-en-jornada-contra-dengue-junto-lideres-comunitarios-cotui (accessed on 25 April 2020).
- Kenneson, A.; Beltran-Ayala, E.; Borbor-Cordova, M.J.; Polhemus, M.E.; Ryan, S.J.; Endy, T.P.; Stewart-Ibarra, A.M. Social-ecological factors and preventive actions decrease the risk of dengue infection at the household-level: Results from a prospective dengue surveillance study in Machala, Ecuador. PLoS Negl. Trop. Dis. 2017, 11, e0006150. [Google Scholar] [CrossRef]
- The Relentless Rise of Two Caribbean Lakes Baffles Scientists. Available online: https://www.nationalgeographic.com/science/article/160303-haiti-dominican-republic-lakes.Nationalgeographic.com (accessed on 25 April 2020).
- Lednicky, J.; De Rochars, V.M.B.; El Badry, M.; Loeb, J.; Telisma, T.; Chavannes, S.; Anilis, G.; Cella, E.; Ciccozzi, M.; Rashid, M.; et al. Zika Virus Outbreak in Haiti in 2014: Molecular and Clinical Data. PLoS Negl. Trop. Dis. 2016, 10, e0004687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UNICEF. A Small Town in the Dominican Republic Becomes a Gateway for Relief Efforts in Haiti. UNICEF: 2010. Available online: https://reliefweb.int/report/haiti/small-town-dominican-republic-becomes-gateway-relief-efforts-haiti (accessed on 25 April 2020).
- Cosner, C.; Beier, J.; Cantrell, R.; Impoinvil, D.; Kapitanski, L.; Potts, M.; Troyo, A.; Ruan, S. The effects of human movement on the persistence of vector-borne diseases. J. Theor. Biol. 2009, 258, 550–560. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Castillo-Chavez, C. The role of residence times in two-patch dengue transmission dynamics and optimal strategies. J. Theor. Biol. 2015, 374, 152–164. [Google Scholar] [CrossRef]
- Dominican Republic Road Network—Logistics Capacity Assessment—Digital Logistics Capacity Assessments. Logcluster.org. Available online: https://dlca.logcluster.org/display/public/DLCA/2.3+Dominican+Republic+Road+Network (accessed on 23 April 2020).
- Sakkas, H.; Bozidis, P.; Giannakopoulos, X.; Sofikitis, N.; Papadopoulou, C. An Update on Sexual Transmission of Zika Virus. Pathogens 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanluca, C.; de Melo, V.C.A.; Mosimann, A.L.P.; dos Santos, G.I.V.; dos Santos, C.N.D.; Luz, K. First report of autochthonous transmission of Zika virus in Brazil. Memórias Do Inst. Oswaldo Cruz. 2015, 110, 569–572. [Google Scholar] [CrossRef]
- Campos, G.S.; Bandeira, A.C.; Sardi, S.I. Zika Virus Outbreak, Bahia, Brazil. Emerg. Infect. Dis. 2015, 21, 1885–1886. [Google Scholar] [CrossRef]
- Ferguson, N.M.; Donnelly, C.; Anderson, R. Transmission intensity and impact of control policies on the foot and mouth epidemic in Great Britain. Nature 2001, 413, 542–548. [Google Scholar] [CrossRef]
- Jombart, T.; Cori, A.; Didelot, X.; Cauchemez, S.; Fraser, C.; Ferguson, N. Bayesian Reconstruction of Disease Outbreaks by Combining Epidemiologic and Genomic Data. PLoS Comput. Biol. 2014, 10, e1003457. [Google Scholar] [CrossRef]
- Salje, H.; Lessler, J.; Paul, K.K.; Azman, A.; Rahman, M.W.; Cummings, D.; Gurley, E.S.; Cauchemez, S. How social structures, space, and behaviors shape the spread of infectious diseases using chikungunya as a case study. Proc. Natl. Acad. Sci. USA 2016, 113, 13420–13425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucharski, A.J.; Kwok, K.O.; Wei, V.W.I.; Cowling, B.; Read, J.; Lessler, J.; Cummings, D.A.; Riley, S. The Contribution of Social Behaviour to the Transmission of Influenza A in a Human Population. PLoS Pathog. 2014, 10, e1004206. [Google Scholar] [CrossRef]
- Zika-Epidemiological Report Dominican Republic. Pan American Health Organisation. 2017. Available online: https://www.paho.org/en/documents/zika-epidemiological-report-dominican-republic-0 (accessed on 25 April 2020).
- Duffy, R.D.; Chen, T.-H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef] [PubMed]
- Olliaro, P.; Fouque, F.; Kroeger, A.; Bowman, L.; Velayudhan, R.; Santelli, A.C.; Garcia, D.; Ramm, R.S.; Sulaiman, L.H.; Tejeda, G.S.; et al. Improved tools and strategies for the prevention and control of arboviral diseases: A research-to-policy forum. PLoS Negl. Trop. Dis. 2018, 12, e0005967. [Google Scholar] [CrossRef] [PubMed]
- Bowman, L.; Runge-Ranzinger, S.; McCall, P.J. Assessing the Relationship between Vector Indices and Dengue Transmission: A Systematic Review of the Evidence. PLoS Negl. Trop. Dis. 2014, 8, e2848. [Google Scholar] [CrossRef]
- Hussain-Alkhateeb, L.; Kroeger, A.; Olliaro, P.; Rocklöv, J.; Sewe, M.O.; Tejeda, G.; Benitez, D.; Gill, B.; Hakim, S.L.; Carvalho, R.G.; et al. Early warning and response system (EWARS) for dengue outbreaks: Recent advancements towards widespread applications in critical settings. PLoS ONE 2018, 13, e0196811. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Disease | |
|---|---|---|
| Mean disease migration interval (days) | Total dengue | Zika |
| 99.55 | 63.28 | |
| Rate (probability of transmission/day) | 0.01004 | 0.01580 |
| Standard deviation | 0.00009095 | 0.0001793 |
| Log likelihood | −66,950 | −39,670 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kingston, R.; Routledge, I.; Bhatt, S.; Bowman, L.R. Novel Epidemic Metrics to Communicate Outbreak Risk at the Municipality Level: Dengue and Zika in the Dominican Republic. Viruses 2022, 14, 162. https://doi.org/10.3390/v14010162
Kingston R, Routledge I, Bhatt S, Bowman LR. Novel Epidemic Metrics to Communicate Outbreak Risk at the Municipality Level: Dengue and Zika in the Dominican Republic. Viruses. 2022; 14(1):162. https://doi.org/10.3390/v14010162
Chicago/Turabian StyleKingston, Rhys, Isobel Routledge, Samir Bhatt, and Leigh R Bowman. 2022. "Novel Epidemic Metrics to Communicate Outbreak Risk at the Municipality Level: Dengue and Zika in the Dominican Republic" Viruses 14, no. 1: 162. https://doi.org/10.3390/v14010162
APA StyleKingston, R., Routledge, I., Bhatt, S., & Bowman, L. R. (2022). Novel Epidemic Metrics to Communicate Outbreak Risk at the Municipality Level: Dengue and Zika in the Dominican Republic. Viruses, 14(1), 162. https://doi.org/10.3390/v14010162