
Identification of Virulence Associated Region during Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus during Attenuation In Vitro: Complex Question with Different Strain Backgrounds



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Construction of Full-Length Chimeric cDNA Clones
2.3. In Vitro Transcription and Virus Recovery
2.4. Viral Growth Kinetics
2.5. Animal and Experimental Design
2.6. Viral Load Assessment
2.7. Cytokine Detection
2.8. Sequencing Analysis of Nine Series of PRRSV Passage Strains
2.9. Statistical Analysis
3. Results
3.1. Construction of Chimeric Clones and Recovery of Viable Chimeric Viruses
3.2. Viral Growth Kinetics on MARC-145 Cells
3.3. Rectal Temperature after Infection
3.4. Lung Macroscopic Scores after Infection
3.5. Virus Copies in Sera
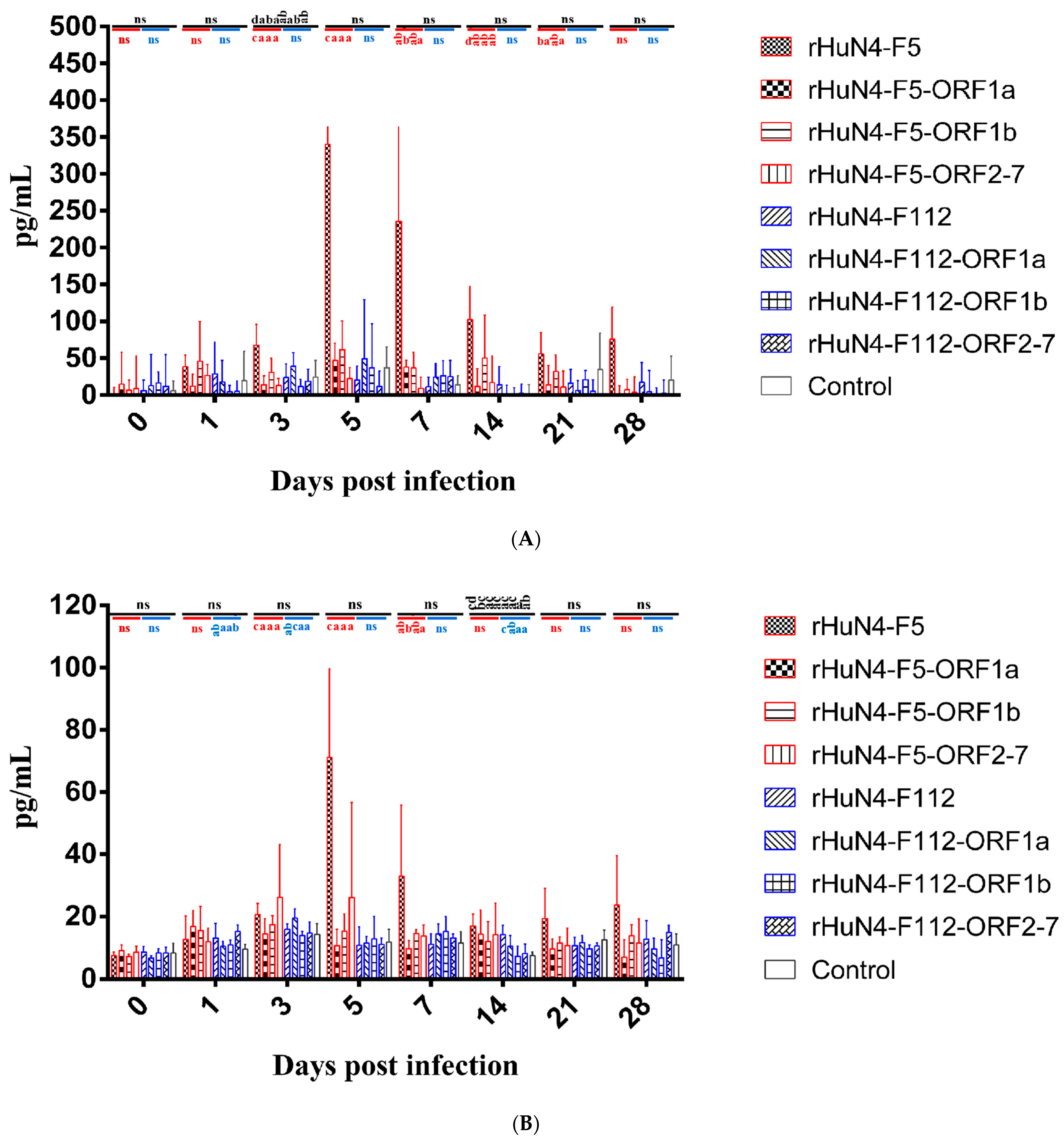
3.6. Results of Cytokine Detection
3.7. Analysis of Genome Change among Nine Series of PRRSV Strain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Wensvoort, G.; de Kluyver, E.P.; Pol, J.M.; Wagenaar, F.; Moormann, R.J.; Hulst, M.M.; Bloemraad, R.; den Besten, A.; Zetstra, T.; Terpstra, C. Lelystad virus, the cause of porcine epidemic abortion and respiratory syndrome: A review of mystery swine disease research at Lelystad. Vet. Microbiol. 1992, 33, 185–193. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, H. Porcine reproductive and respiratory syndrome in China. Virus Res. 2010, 154, 31–37. [Google Scholar] [CrossRef]
- Benfield, D.A.; Nelson, E.; Collins, J.E.; Harris, L.; Goyal, S.M.; Robison, D.; Christianson, W.T.; Morrison, R.B.; Gorcyca, D.; Chladek, D. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Investig. 1992, 4, 127–133. [Google Scholar] [CrossRef]
- Meulenberg, J.J.; Hulst, M.M.; de Meijer, E.J.; Moonen, P.L.; den Besten, A.; de Kluyver, E.P.; Wensvoort, G.; Moormann, R.J. Lelystad virus belongs to a new virus family, comprising lactate dehydrogenase-elevating virus, equine arteritis virus, and simian hemorrhagic fever virus. Arch. Virol. Suppl. 1994, 9, 441–448. [Google Scholar] [PubMed]
- Snijder, E.J.; Meulenberg, J.J. The molecular biology of arteriviruses. J. Gen. Virol. 1998, 79 Pt 5, 961–979. [Google Scholar] [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses. Arch. Virol. 2016, 161, 2921–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, J.H.; Lauck, M.; Bailey, A.L.; Shchetinin, A.M.; Vishnevskaya, T.V.; Bao, Y.; Ng, T.F.; LeBreton, M.; Schneider, B.S.; Gillis, A.; et al. Reorganization and expansion of the nidoviral family Arteriviridae. Arch. Virol. 2016, 161, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Bautista, E.M.; Meulenberg, J.J.; Choi, C.S.; Molitor, T.W. Structural polypeptides of the American (VR-2332) strain of porcine reproductive and respiratory syndrome virus. Arch. Virol. 1996, 141, 1357–1365. [Google Scholar] [CrossRef]
- Firth, A.E.; Zevenhoven-Dobbe, J.C.; Wills, N.M.; Go, Y.Y.; Balasuriya, U.B.; Atkins, J.F.; Snijder, E.J.; Posthuma, C.C. Discovery of a small arterivirus gene that overlaps the GP5 coding sequence and is important for virus production. J. Gen. Virol. 2011, 92 Pt 5, 1097–1106. [Google Scholar] [CrossRef]
- Fang, Y.; Snijder, E.J. The PRRSV replicase: Exploring the multifunctionality of an intriguing set of nonstructural proteins. Virus Res. 2010, 154, 61–76. [Google Scholar] [CrossRef] [PubMed]
- van Aken, D.; Zevenhoven-Dobbe, J.; Gorbalenya, A.E.; Snijder, E.J. Proteolytic maturation of replicase polyprotein pp1a by the nsp4 main proteinase is essential for equine arteritis virus replication and includes internal cleavage of nsp7. J. Gen. Virol. 2006, 87, 3473–3482. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.H.; Fang, Y.; Rowland, R.R.R.; Lawson, S.R.; Christopher-Hennings, J.; Yoon, K.J.; Nelson, E.A. The 2b protein as a minor structural component of PRRSV. Virus Res. 2005, 114, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Scortti, M.; Prieto, C.; Alvarez, E.; Simarro, I.; Castro, J.M. Failure of an inactivated vaccine against porcine reproductive and respiratory syndrome to protect gilts against a heterologous challenge with PRRSV. Vet. Rec. 2007, 161, 809–813. [Google Scholar] [PubMed]
- Prieto, C.; Martinez-Lobo, F.J.; Diez-Fuertes, F.; Aguilar-Calvo, P.; Simarro, I.; Castro, J.M. Immunisation of pigs with a major envelope protein sub-unit vaccine against porcine reproductive and respiratory syndrome virus (PRRSV) results in enhanced clinical disease following experimental challenge. Vet. J. 2011, 189, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, J.L.; Zeng, J.W.; Yin, S.Y.; Li, Y.H.; Zheng, L.Y.; Guo, X.; Ge, X.N.; Yang, H.C. The 30-Amino-Acid Deletion in the Nsp2 of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus Emerging in China Is Not Related to Its Virulence. J. Virol. 2009, 83, 5156–5167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Zhou, L.; Sun, W.; Zhang, P.; Ge, X.; Guo, X.; Han, J.; Yang, H. Nonstructural protein 9 residues 586 and 592 are critical sites in determining the replication efficiency and fatal virulence of the Chinese highly pathogenic porcine reproductive and respiratory syndrome virus. Virology 2018, 517, 135–147. [Google Scholar] [CrossRef]
- Zhao, K.; Gao, J.C.; Xiong, J.Y.; Guo, J.C.; Yang, Y.B.; Jiang, C.G.; Tang, Y.D.; Tian, Z.J.; Cai, X.H.; Tong, G.Z.; et al. Two Residues in NSP9 Contribute to the Enhanced Replication and Pathogenicity of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2018, 92, e02209-17. [Google Scholar] [CrossRef] [Green Version]
- Allende, R.; Kutish, G.F.; Laegreid, W.; Lu, Z.; Lewis, T.L.; Rock, D.L.; Friesen, J.; Galeota, J.A.; Doster, A.R.; Osorio, F.A. Mutations in the genome of porcine reproductive and respiratory syndrome virus responsible for the attenuation phenotype. Arch. Virol. 2000, 145, 1149–1161. [Google Scholar] [CrossRef]
- Kwon, B.; Ansari, I.H.; Pattnaik, A.K.; Osorio, F.A. Identification of virulence determinants of porcine reproductive and respiratory syndrome virus through construction of chimeric clones. Virology 2008, 380, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liang, Y.; Han, J.; Burkhart, K.M.; Vaughn, E.M.; Roof, M.B.; Faaberg, K.S. Attenuation of porcine reproductive and respiratory syndrome virus strain MN184 using chimeric construction with vaccine sequence. Virology 2008, 371, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef] [PubMed]
- Tong, G.Z.; Zhou, Y.J.; Hao, X.F.; Tian, Z.J.; An, T.Q.; Qiu, H.J. Highly pathogenic porcine reproductive and respiratory syndrome, China. Emerg. Infect. Dis. 2007, 13, 1434–1436. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Hao, X.F.; Tian, Z.J.; Tong, G.Z.; Yoo, D.; An, T.Q.; Zhou, T.; Li, G.X.; Qiu, H.J.; Wei, T.C.; et al. Highly virulent porcine reproductive and respiratory syndrome virus emerged in China. Transbound. Emerg. Dis. 2008, 55, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.J.; An, T.Q.; Zhou, Y.J.; Peng, J.M.; Hu, S.P.; Wei, T.C.; Jiang, Y.F.; Xiao, Y.; Tong, G.Z. An attenuated live vaccine based on highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) protects piglets against HP-PRRS. Vet. Microbiol. 2009, 138, 34–40. [Google Scholar] [CrossRef]
- Zhang, S.; Zhou, Y.; Zhu, J.; Tong, W.; Jiang, Y.; Tong, G. Generation of a infectious cDNA clone of highly pathogenic porcine reproductive and respiratory syndrome virus HuN4. Chin. J. Prev. Vet. Med. 2011, 33, 497–502. [Google Scholar]
- Zhang, S.; Zhou, Y.; Jiang, Y.; Li, G.; Yan, L.; Yu, H.; Tong, G. Generation of an infectious clone of HuN4-F112, an attenuated live vaccine strain of porcine reproductive and respiratory syndrome virus. Virol. J. 2011, 8, 410. [Google Scholar] [CrossRef] [Green Version]
- Thanawongnuwech, R.; Brown, G.B.; Halbur, P.G.; Roth, J.A.; Royer, R.L.; Thacker, B.J. Pathogenesis of porcine reproductive and respiratory syndrome virus-induced increase in susceptibility to Streptococcus suis infection. Vet. Pathol. 2000, 37, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, T.C.; Tian, Z.J.; An, T.Q.; Zhou, Y.J.; Xiao, Y.; Jiang, Y.F.; Hao, X.F.; Zhang, S.R.; Peng, J.M.; Qiu, H.J.; et al. Development and application of Taq Man-MGB fluorescence quantitative RT-PCR assay for detection of porcine reproductive and respiratory syndrome virus. Chin. J. Prev. Vet. Med. 2008, 30, 944–948. [Google Scholar]
- Jiang, Y.F.; Xia, T.Q.; Zhou, Y.J.; Yu, L.X.; Yang, S.; Huang, Q.F.; Li, L.W.; Gao, F.; Qu, Z.H.; Tong, W.; et al. Characterization of three porcine reproductive and respiratory syndrome virus isolates from a single swine farm bearing strong homology to a vaccine strain. Vet. Microbiol. 2015, 179, 242–249. [Google Scholar] [CrossRef]
- Song, S.; Xu, H.; Zhao, J.; Leng, C.; Xiang, L.; Li, C.; Fu, J.; Tang, Y.D.; Peng, J.; Wang, Q.; et al. Pathogenicity of NADC34-like PRRSV HLJDZD32-1901 isolated in China. Vet. Microbiol. 2020, 246, 108727. [Google Scholar] [CrossRef] [PubMed]
- Van Doorsselaere, J.; Brar, M.S.; Shi, M.; Karniychuk, U.; Leung, F.C.; Nauwynck, H.J. Complete genome characterization of a East European Type 1 subtype 3 porcine reproductive and respiratory syndrome virus. Virus Genes 2012, 44, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Faaberg, K.S.; Kehrli, M.E., Jr.; Lager, K.M.; Guo, B.; Han, J. In vivo growth of porcine reproductive and respiratory syndrome virus engineered nsp2 deletion mutants. Virus Res. 2010, 154, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Morgan, S.B.; Graham, S.P.; Salguero, F.J.; Sanchez Cordon, P.J.; Mokhtar, H.; Rebel, J.M.; Weesendorp, E.; Bodman-Smith, K.B.; Steinbach, F.; Frossard, J.P. Increased pathogenicity of European porcine reproductive and respiratory syndrome virus is associated with enhanced adaptive responses and viral clearance. Vet. Microbiol. 2013, 163, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, L.; Zhang, J.; Ge, X.; Zhou, R.; Zheng, H.; Geng, G.; Guo, X.; Yang, H. Nsp9 and Nsp10 contribute to the fatal virulence of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China. PLoS Pathog. 2014, 10, e1004216. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, X.; Lunney, J.K.; Lawson, S.; Sun, Z.; Brown, E.; Christopher-Hennings, J.; Knudsen, D.; Nelson, E.; Fang, Y. Immunodominant epitopes in nsp2 of porcine reproductive and respiratory syndrome virus are dispensable for replication, but play an important role in modulation of the host immune response. J. Gen. Virol. 2010, 91 Pt 4, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Duff, G.W.; Durum, S.K. The pyrogenic and mitogenic actions of interleukin-1 are related. Nature 1983, 304, 449–451. [Google Scholar] [CrossRef] [PubMed]
- McLaren, J.E.; Ramji, D.P. Interferon gamma: A master regulator of atherosclerosis. Cytokine Growth Factor Rev. 2009, 20, 125–135. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef]
- Saper, C.B.; Breder, C.D. The neurologic basis of fever. N. Engl. J. Med. 1994, 330, 1880–1886. [Google Scholar]
- Burgara-Estrella, A.; Diaz, I.; Rodriguez-Gomez, I.M.; Essler, S.E.; Hernandez, J.; Mateu, E. Predicted Peptides from Non-Structural Proteins of Porcine Reproductive and Respiratory Syndrome Virus Are Able to Induce IFN-gamma and IL-10. Viruses 2013, 5, 663–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudmundsdottir, I.; Risatti, G.R. Infection of porcine alveolar macrophages with recombinant chimeric porcine reproductive and respiratory syndrome virus: Effects on cellular gene transcription and virus growth. Virus Res. 2009, 145, 145–150. [Google Scholar] [CrossRef]
- Bautista, E.M.; Faaberg, K.S.; Mickelson, D.; McGruder, E.D. Functional properties of the predicted helicase of porcine reproductive and respiratory syndrome virus. Virology 2002, 298, 258–270. [Google Scholar] [CrossRef] [Green Version]
- den Boon, J.A.; Snijder, E.J.; Chirnside, E.D.; de Vries, A.A.; Horzinek, M.C.; Spaan, W.J. Equine arteritis virus is not a togavirus but belongs to the coronaviruslike superfamily. J. Virol. 1991, 65, 2910–2920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, J.; Wang, L.H.; Quan, R.; Fu, Y.; Zhang, H.X.; Feng, W.H. Induction of interleukin-10 is dependent on p38 mitogen-activated protein kinase pathway in macrophages infected with porcine reproductive and respiratory syndrome virus. Virol. J. 2012, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Wongyanin, P.; Buranapraditkul, S.; Yoo, D.W.; Thanawongnuwech, R.; Roth, J.A.; Suradhat, S. Role of porcine reproductive and respiratory syndrome virus nucleocapsid protein in induction of interleukin-10 and regulatory T-lymphocytes (T-reg). J. Gen. Virol. 2012, 93, 1236–1246. [Google Scholar] [CrossRef]
- Chung, H.K.; Chae, C. Expression of interleukin-10 and interleukin-12 in piglets experimentally infected with porcine reproductive and respiratory syndrome virus (PRRSV). J. Comp. Pathol. 2003, 129, 205–212. [Google Scholar] [CrossRef]
- Diaz, I.; Darwich, L.; Pappaterra, G.; Pujols, J.; Mateu, E. Immune responses of pigs after experimental infection with a European strain of Porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2005, 86, 1943–1951. [Google Scholar] [CrossRef]
- Meier, W.A.; Galeota, J.; Osorio, F.A.; Husmann, R.J.; Schnitzlein, W.M.; Zuckermann, F.A. Gradual development of the interferon-gamma response of swine to porcine reproductive and respiratory syndrome virus infection or vaccination. Virology 2003, 309, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Calvert, J.G.; Slade, D.E.; Shields, S.L.; Jolie, R.; Mannan, R.M.; Ankenbauer, R.G.; Welch, S.K. CD163 expression confers susceptibility to porcine reproductive and respiratory syndrome viruses. J. Virol. 2007, 81, 7371–7379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, P.B.; Dinh, P.X.; Ansari, I.H.; de Lima, M.; Osorio, F.A.; Pattnaik, A.K. The minor envelope glycoproteins GP2a and GP4 of porcine reproductive and respiratory syndrome virus interact with the receptor CD163. J. Virol. 2010, 84, 1731–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Number | Macroscopic (Lung) | |||
|---|---|---|---|---|---|
| Mean ± SD | Pathological Changes (Score) | ||||
| ≤30 | 30 to 50 | ≥50 | |||
| rHuN4-F5 | 5 | 65 ± 27.36 | 0 | 3 | 2 |
| rHuN4-F5-ORF1a | 5 | 30.8 ± 14.72 | 2 | 3 | 0 |
| rHuN4-F5-ORF1b | 5 | 33.4 ± 17.08 | 2 | 3 | 0 |
| rHuN4-F5-ORF2-7 | 5 | 20 ± 15.03 | 5 | 0 | 0 |
| rHuN4-F112 | 5 | 17.6 ± 11.15 | 5 | 0 | 0 |
| rHuN4-F112-ORF1a | 5 | 36.8 ± 21.11 | 3 | 1 | 1 |
| rHuN4-F112-ORF1b | 5 | 41.6 ± 19.76 | 3 | 1 | 1 |
| rHuN4-F112-ORF2-7 | 5 | 27.4 ± 14.19 | 4 | 0 | 1 |
| Control | 5 | 15.2 ± 13.92 | 5 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Tong, W.; Yu, L.; Li, L.; Gao, F.; Li, G.; Liu, C.; Chen, P.; Shen, Q.; Zhang, Y.; et al. Identification of Virulence Associated Region during Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus during Attenuation In Vitro: Complex Question with Different Strain Backgrounds. Viruses 2022, 14, 40. https://doi.org/10.3390/v14010040
Jiang Y, Tong W, Yu L, Li L, Gao F, Li G, Liu C, Chen P, Shen Q, Zhang Y, et al. Identification of Virulence Associated Region during Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus during Attenuation In Vitro: Complex Question with Different Strain Backgrounds. Viruses. 2022; 14(1):40. https://doi.org/10.3390/v14010040
Chicago/Turabian StyleJiang, Yifeng, Wu Tong, Lingxue Yu, Liwei Li, Fei Gao, Guoxin Li, Changlong Liu, Pengfei Chen, Qi Shen, Yujiao Zhang, and et al. 2022. "Identification of Virulence Associated Region during Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus during Attenuation In Vitro: Complex Question with Different Strain Backgrounds" Viruses 14, no. 1: 40. https://doi.org/10.3390/v14010040
APA StyleJiang, Y., Tong, W., Yu, L., Li, L., Gao, F., Li, G., Liu, C., Chen, P., Shen, Q., Zhang, Y., Zhou, Y., & Tong, G. (2022). Identification of Virulence Associated Region during Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus during Attenuation In Vitro: Complex Question with Different Strain Backgrounds. Viruses, 14(1), 40. https://doi.org/10.3390/v14010040